神经细胞黏附分子衍生肽FGL药理作用研究进展
2016-02-14王苏平储成艳
王苏平,高 月,储成艳,李 深
(大连市中心医院神经内一科,辽宁大连 116033)
神经细胞黏附分子衍生肽FGL药理作用研究进展
王苏平,高 月,储成艳,李 深
(大连市中心医院神经内一科,辽宁大连 116033)
神经细胞黏附分子(NCAM)具有诱导神经新生、促进神经分化、保护神经细胞等生物学功能,对于神经系统的发育和功能起着关键性调节作用。FGL是根据NCAM的空间结构和其与成纤维细胞生长因子受体的结合位点设计的含15个氨基酸的生物活性多肽。作为NCAM的功能核心片段,FGL除了可保护神经、诱导神经分化外,还具备抗炎和调节神经可塑性等多种药理作用,在缺血性脑损伤、老年认知障碍、阿尔茨海默病和抑郁症等神经精神疾病的治疗应用上显现出巨大的价值。不仅如此,临床试验结果显示,FGL对人体是安全的,阿尔茨海默病的临床Ⅰ期试验正在进行中。FGL极有可能成为用于治疗神经精神疾病的有效药物,具有广阔的临床应用前景。
FGL;神经细胞黏附分子;神经精神疾病;多肽
神经细胞黏附分子(neural cell adhesion mol⁃ecule,NCAM)属于免疫球蛋白超家族细胞黏附分子,主要表达于神经元和胶质细胞表面,是中枢神经系统内表达量最大的细胞黏附分子。按相对分子质量的大小,主要分为NCAM120、NCAM140和NCAM180三种亚型,它们的胞外段均包含5个免疫球蛋白(Ig)样结构域和2个纤连蛋白Ⅲ(fibro⁃nectinⅢ,FNⅢ)结构域[1-2]。NCAM表达异常会导致新生神经元的突起生长受阻,阻断神经细胞之间的联系,不仅影响神经系统的发育,且与精神分裂症和抑郁症等精神疾病的发生密切相关[3-4]。此外,NCAM还能够促进脊髓损伤和脑外伤等神经损伤后的再生与修复,NCAM缺乏则会促进损伤部位神经细胞的凋亡[3,5]。NCAM功能的发挥主要依赖于胞外部分通过与嗜性结合(NCAM-NCAM)和异嗜性结合激活细胞内信号传导通路。因此,学者们试图通过研究NCAM下游分子与其结合的部位合成能模拟NCAM功能的多肽片段。目前已合成C3,P2和plannexin等多种NCAM衍生肽[6-7]。FGL(FG loop)是根据NCAM的空间结构和成纤维细胞生长因子受体(fibroblast growth factor receptors,FGFR)的结合位点设计的一段活性多肽片段,相当于NCAM中第2个FNⅢ中的F和Gβ折叠与它们之间的连接环。NCAM很多重要功能都是通过与FG⁃FR胞外IgⅡ-Ⅲ片段结合实现,FGL可直接激活FGFR介导的信号传导通路,诱导神经细胞分化、保护神经细胞和抗炎等,并能增强大鼠的学习能力和空间、社交记忆,促进新生大鼠的运动协调[8-13]。FGL因其功能多样性及独特的药理学特性和潜在的临床应用前景在众多NCAM衍生肽中脱颖而出,成为众多NCAM衍生肽中的研究焦点。本文就FGL的结构、合成方法、药理学作用及其治疗神经精神疾病的进展进行综述。
1 FGL结构、种类及其合成方法
FGL是一种氨基酸序列为EVYVVAENQQGK⁃SKA的十五肽,相当于人类、大鼠和小鼠NCAM的Glu681-Ala695。目前体内外的研究对象为FGL的二聚体和四聚体,FGL单体虽然也能激活FGFR,但其效能远低于二聚体和四聚体。FGL二聚体主要有2种合成方法(图1),一种是通过Fmoc固相多肽合成法,在多肽的N端利用亚氨基二乙酸将2个FGL单体连接,C端乙酰化处理,这种设计能同时结合2个FGFR,对受体的磷酸化和下游信号的传导起着关键作用[14]。另一种方法同样进行Fmoc固相多肽合成,但是利用赖氨酸残基在多肽C端连接2个FGL单体,上述2种方法合成的FGL均有活性。第一种是目前通用的合成方法,这种通过N端连接的FGL二聚体又称为FGLL。FGL四聚体(FGLd)是4个FGL单体以赖氨酸为骨架合成的树枝状分子。二聚体与四聚体FGL具有相似的生物学功能,但FGLL拥有分子质量小、可经皮下注射等特征,且已被用于临床试验而研究更为广泛。
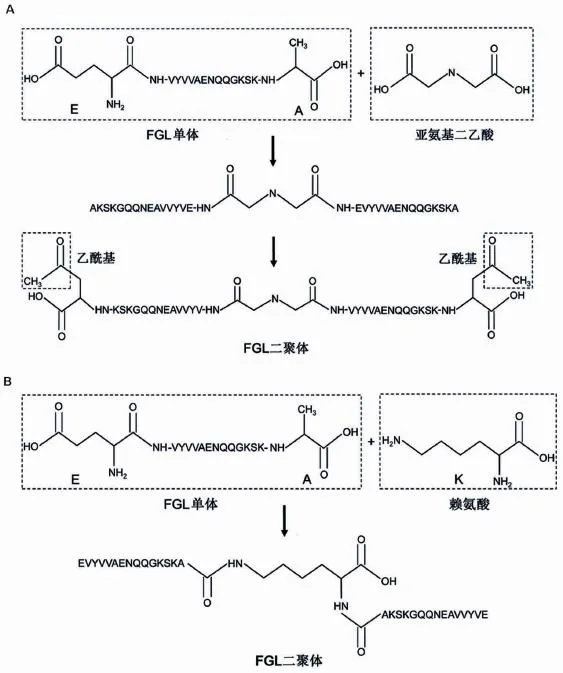
图1 FGL二聚体的结构及合成路线.A:亚氨基二乙酸的2个羧基通过与FGL N端氨基的结合连接2个单体,C端乙酰化处理;B:赖氨酸的2个氨基通过与FGL C端羧基的结合连接2个单体.
2 FGL多肽在结构和功能上的优点
完整蛋白质的合成费用昂贵,且相对不稳定,特别是在体内实验中不易通过血脑屏障,并具有免疫原性。合成的活性多肽可通过生理性受体途径激发快速活化或抑制性反应,且多肽分子小,相对稳定,无免疫原性,人工化学合成的难度和费用均远低于完整蛋白质。FGL作为NCAM衍生肽拥有上述活性肽的所有特性,在功能上被认为是FGFR的激动剂,并且可模拟NCAM与FGFR结合从而直接激活相应的信号通路[15]。大鼠、犬和猴的临床前研究表明FGL无全身毒性。同时,临床试验表明,健康人对FGL具有良好的耐受性,经鼻吸入FGL后身体各项生理指标均未出现明显异常。另外,FGL在体内的药代动力学与给药剂量相关,人体吸入FGL 25 mg后,血液中未能检测到FGL,而吸入FGL 100和200 mg后,其在血液中存在的时间分别为1和4 h,药峰浓度分别为0.52和1.38 μg·L-1[11]。
3 FGL体内给药途径及其分布
FGL因结构差异有不同的体内给药途径,FGLd主要通过枕骨下、脑室和鼻腔途径给药,其中,经立体定位仪行脑室注射比较常用。FGLd经脑室注射后10 min便可出现在海马齿状回中,还能扩散至隔核、皮质、纹状体、海马CA1和CA3区,应用免疫组织化学染色可清晰地观察到FGL分布在细胞膜表面和胞内侧[16]。除适用所有四聚体结构的给药途径外,FGLL还可经皮下注射,利用酶联免疫吸附实验检测FGLL经皮下注射后在成年大鼠血液和脑脊液中的浓度,发现注射10 min后即可在血液和脑脊液中监检到FGLL,并持续5 h,表明FGLL能通过血脑屏障。实验还发现,注射后第一个2 h血液中的FGLL平均浓度是脑脊液中的10~17倍。FGLL经皮下注射1 h后就能在海马中检测出FGFR磷酸化,提示海马可能是FGL的主要作用脑区[17]。
4 FGL对缺血性脑损伤的保护作用
FGL可明显促进新生大鼠的海马神经元、7日龄大鼠的小脑颗粒神经元(cerebellar granule neu⁃rons,CGN)和15日龄大鼠的多巴胺神经元神经突起的生长,提高神经毒物6-羟基多巴胺暴露下的多巴胺神经元存活率、抑制氯化钾浓度改变导致的CGN凋亡[13]。进一步的研究发现,FGL诱导神经分化及神经保护作用依赖于FGFR及其下游丝裂原活化蛋白激酶(mitogen-activated protein kinas⁃es,MAPK)和磷脂酰肌醇3-激酶(phosphati⁃dylinositol 3-kinase,PI3K)信号通路。将编码成纤维细胞生长因子受体1显性负性分子(dominant negative FGFR1,dnFGFR1)通过载体转染至CGN。结果显示,dnFGFR1可完全阻断FGL对CGN神经突起生长的促进作用。在CGN培养基中分别加入FGFR1抑制剂、MEK抑制剂或PI3K抑制剂,不仅可明显抑制FGL诱导的神经突起生长的作用,还能降低FGL促神经元存活的效应[13]。最近的研究还发现,接头蛋白Scr同源和胶原蛋白A(src homologous and collagen A)、底物2α和磷脂酶Cγ也参与FGL诱导神经生长的作用[18]。
Skibo等[20]的研究显示,FGL还能维持缺糖低氧环境下海马神经元的代谢能力和突触前功能,这对于损伤后脑功能的维持与恢复有重要意义。在沙鼠短暂全脑缺血模型中,一次性皮下注射FGL可显著抑制因缺血导致的海马神经元死亡,进而有效促进沙鼠脑损伤后神经功能的恢复。Pedersen等[21]研究显示,FGL能显著恢复外伤性脑损伤后的神经功能,并应用基因组学研究方法探讨其产生这种效应的途径。结果发现,FGL通过上调损伤大鼠体内的Fas凋亡抑制性分子、G蛋白信号转导调节蛋白14等基因表达促进神经新生、抑制神经元的退行性病变和死亡,FGL还可通过调控基因的表达参与损伤动物脑组织的神经分化、突触传递和细胞循环等生理过程。
5 FGL抗老年认知障碍作用
年龄相关的认知障碍与脑组织中促炎因子水平的上升和胶质细胞活性的增强共同导致的神经炎症反应密切相关[22-23]。Ojo等[24]研究发现,FGL可降低老年大鼠小胶质细胞活性标志物(CD86)和促炎因子白细胞介素1β(interleukin-1β,IL-1β)的表达,同时抑制老年大鼠海马中神经胶质细胞的活性、降低小胶质细胞密度进而促进老年大鼠认知能力的恢复[11]。体外实验表明,FGL能促进胶质细胞释放抗炎因子IL-4,IL-4有助于神经元CD200的表达,而CD200则可通过维持小胶质细胞处于静息状态保护突触功能[25-26]。后续的体内研究结果同样表明,FGL对小胶质细胞的活性抑制作用就是依赖于CD200表达的增加[11]。另外,FGL还能有效抑制海马中具有促进CD200表达作用的胰岛素样生长因子1水平的下降,同时阻止小胶质细胞激活物干扰素γ(interferon-γ,IFN-γ)的增加[27-28]。
突触可塑性的调节被认为是学习与记忆的神经生物学基础[29-30]。FGL可通过加快海马神经元突触小泡的释放和回收来增强突触前功能,同时促进突触的形成及长时程增强的持续时间,以延缓或阻止老年认知障碍的进程[31]。除引起突触功能的变化外,FGL还能通过改变突触的形态结构调节突触可塑性。Popov等[32]利用三维重建技术观察到FGL对老年大鼠海马的体积、突触数目和棘突密度均无影响,但不同类型棘突上的突触分布却发生了变化,刷状棘突突触数显著增加,细长棘突突触数则明显减少,而短粗棘突突触和树突干突触数几乎没有变化。大部分刷状棘突突触都是穿孔型的,这种突触比非穿孔型含有更多的α-氨基-3-羟基-5-甲基-4-异唑丙酸(α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid,AMPA)和N-甲基-D-天冬氨 酸(N-methyl-D-aspartafe,NMDA)受体[33-34]。Ojo等[34]研究发现,FGL可有效降低大鼠因老年化所导致的海马CA3区和门区突触泡蛋白的减少。Knafo等[14]应用生化学、形态学、电生理学和行为学方法探究FGL调节突触可塑性的作用机制,发现FGL通过易化突触AMPA引起海马CA1区神经元突触传递的持续增强,同时强化NMDA介导的长时程增强,而这些效应都依赖于FGFR、蛋白激酶C和钙调蛋白依赖性蛋白激酶Ⅱ的活化。
6 FGL抗阿尔茨海默病(Alzheimer dis⁃ease,AD)作用
FGL在体外可降低β淀粉样蛋白肽片段25-35(Aβ25-35)导致的海马神经元的死亡[13]。体内研究则证实,FGL能阻止年轻大鼠脑室内注射Aβ25-35后导致的AD样短时程记忆减退和认知障碍。Klemen⁃tiev等[35]依据Aβ25-35产生的神经病理特征建立早期AD模型,同时探讨FGL对AD的治疗是否有积极意义。结果发现,FGL主要通过抑制糖原合酶激酶3β(glycogen synthase kinase-3 beta,GSK-3β)活性阻止Aβ25-35引起的内源性Aβ沉积、Tau蛋白磷酸化、神经元退化等神经病理变化和认知障碍。GSK-3β的主要功能是调节微管稳定性、细胞凋亡、神经突起回缩和认知功能[36-38]。Corbett等[32]最近研究显示,FGL明显减少了Aβ25-35导致的海马背侧CA1区锥体神经细胞的丢失,这种作用同样与GSK-3β的失活有关。目前,FGL正用于AD患者的临床治疗试验[39]。
7 FGL抗癫痫作用
基于FGL高效的神经保护、神经营养和抗炎等药理作用,目前在神经退行性疾病和脑损伤的动物实验研究中均已取得了一定进展。最近,Zel⁃linger等[40]从行为学和细胞学层面上分析了FGL经皮下注射后对点燃小鼠模型癫痫发作的影响。FGL治疗明显降低了癫痫发作的阈值,但对点燃小鼠的行为学无影响。另外,点燃小鼠癫痫模型海马细胞的增殖率显著提高,高剂量的FGL(10 mg·kg-1)则可有效降低海马中神经前体细胞数目,而低剂量(2 mg·kg-1)对相关的改变无影响。总之,FGL对点燃模型癫痫的改善没有确凿证据,未来的研究需要继续评估FGL在自发性癫痫模型对癫痫发生、发展的影响。
8 FGL抗抑郁和抗精神分裂症作用
据文献报道,FGL可改善大鼠因慢性应激导致的海马新生细胞数目减少,继而阻止慢性应激对大鼠空间记忆能力的损害[19];FGF2-FGFR信号通路具有显著的抗抑郁作用[41]。Turner等[42]将FGL和FGF2分别注射到大鼠侧脑室,发现均可改善大鼠在强迫性游泳实验中的行为表现。与此同时,Aon⁃urm-Helm等[43]对野生型和NCAM-/-突变型小鼠进行悬尾实验和糖水偏好实验。结果表明,NCAM-/-小鼠表现出抑郁情绪,经过FGL一次性注射或重复注射后,NCAM-/-小鼠的抑郁样行为明显减轻。这种效应可能与FGL对新生神经元的保护作用以及对脑组织中环磷腺苷效应元件结合蛋白的活化密切相关。认知缺陷被认为是精神分裂症最典型的特征,其中包括工作记忆缺陷[44]。Secher等[45]通过利用苯环利定处理新生大鼠建立表现出与精神分裂症相关的长时程学习和记忆障碍的动物模型,这种障碍体现在工作记忆和反转学习上,而在参考记忆任务中的长时程记忆并未受到影响。经过FGL长期治疗后,反转学习障碍虽未得到改善,但工作记忆能力有了明显恢复,几乎达到对照组的水平。
单独将FGL注射到健康的年轻大鼠体内,虽然齿状回未成熟的神经元数目有所增加,但背侧海马体积明显减小,CA1和CA3区锥体神经元数目下降明显,提示FGL的体内作用可能与年龄和生理状况密切相关[17,46]。尽管如此,FGL在神经精神疾病治疗中的积极作用已得到广泛的证实。近年来,FGL在再生医学及组织工程学领域中的应用逐渐受到关注。研究发现,FGL可促进神经干细胞的增殖与分化,甚至能诱导人类胚胎干细胞向神经元方向分化[6,47]。将FGL与自聚合肽纳米纤维材料结合建立的三维培养体系更有利于神经细胞的分化。最新的研究表明,FGL还能促进该三维体系中脊髓来源神经干细胞的增殖、迁移、黏附及分化[48]。因此,随着干细胞疗法与组织工程技术的快速发展,将FGL与这些领先的治疗方法结合来治疗至今难以攻克的疾病会有广阔前景。
[1]Weledji EP,Assob JC.The ubiquitous neural cell adhesion molecule(N-CAM)[J].Ann Med Surg(Lond),2014,3(3):77-81.
[2]Irintchev A,Schachner M.The injured and regener⁃ating nervous system:immunoglobulin superfamily members as key players[J].Neuroscientist,2012,18(5):452-466.
[3]Li S,Leshchyns'ka I,Chernyshova Y,Schachner M,Sytnyk V.The neural cell adhesion molecule(NCAM)associates with and signals through p21-activated kinase 1(Pak1)[J].J Neurosci,2013,33(2):790-803.
[4]Vawter MP.Dysregulation of the neural cell adhe⁃sion molecule and neuropsychiatric disorders[J].Eur J Pharmacol,2000,405(1-3):385-395.
[5]Skaper SD.Neuronal growth-promoting and inhibi⁃tory cues in neuroprotection and neuroregenera⁃tion[J].Methods Mol Biol,2012,846:13-22.
[6]Xu R,Feyeux M,Julien S,Nemes C,Albrecht⁃sen M,Dinnyés A,et al.Screening of bioactive peptides using an embryonic stem cell-based neu⁃ron differentiation assay[J].AAPS J,2014,16(3):400-412.
[7]Mie M,Sasaki S,Kobatake E.Construction of a bFGF-tethered multi-functional extracellular matrix protein through coiled-coil structures for neurite outgrowth induction[J].Biomed Mater,2014,9(1):015004.
[8]Irintchev A,Schachner M.The injured and regener⁃ating nervous system:immunoglobulin superfamily members as keyplayers[J].Neuroscientist,2012,18(5):452-466.
[9]Ono S,Hane M,Kitajima K,Sato C.Novel regula⁃tion of fibroblast growth factor 2(FGF2)-mediated cell growth by polysialic acid[J].J Biol Chem,2012,287(6):3710-3722.
[10]Norden DM,Godbout JP.Review:microglia of the aged brain:primed to be activated and resistant to regulation[J].Neuropathol Appl Neurobiol,2013,39(1):19-34.
[11]Woodbury ME,Ikezu T.Fibroblast growth factor-2 signaling in neurogenesis and neurodegeneration[J].J Neuroimmune Pharmacol,2014,9(2):92-101.
[12]Tam RY,Fuehrmann T,Mitrousis N,Shoichet MS. Regenerative therapies for central nervous system diseases:a biomaterials approach[J].Neuropsy⁃chopharmacology,2014,39(1):169-188.
[13]Neiiendam JL,Køhler LB,Christensen C,Li S,Pedersen MV,Ditlevsen DK,et al.An NCAM-derived FGF-receptor agonist,the FGL-peptide, induces neurite outgrowth and neuronal survival in primary rat neurons[J].J Neurochem,2004,91(4):920-935.
[14]Knafo S,Venero C,Sánchez-Puelles C,Pereda-Peréz I,Franco A,Sandi C,et al.Facilitation of AMPA receptor synaptic delivery as a molecular mechanism for cognitive enhancement[J].PLoS Biol,2012,10(2):e1001262.
[15]Bojesen KB,Clausen O,Rohde K,Christensen C,Zhang L,Li S,et al.Nectin-1 binds and signals through the fibroblast growth factor receptor[J].J Biol Chem,2012,287(44):37420-37433.
[16]Dallérac G,Zerwas M,Novikova T,Callu D,Leb⁃lanc-Veyrac P,Bock E,et al.The neural cell adhe⁃sion molecule-derived peptide FGL facilitates long-term plasticity in the dentate gyrusin vivo[J].Learn Mem,2011,18(5):306-313.
[17]Corbett NJ,Gabbott PL,Klementiev B,Davies HA,Colyer FM,Novikova T,et al.Amyloid-beta in⁃duced CA1 pyramidal cell loss in young adult rats is alleviated by systemic treatment with FGL,a neuralcelladhesion molecule-derived mimetic peptide[J].PLoS One,2013,8(8):e71479.
[18]Chen Y,Li S,Berezin V,Bock E.The fibroblast growth factor receptor(FGFR)agonist FGF1 and the neural cell adhesion molecule-derived peptide FGL activate FGFR substrate 2alpha differently[J].J Neurosci Res,2010,88(9):1882-1889.
[19]Borcel E,Pérez-Alvarez L,Herrero AI,Brionne T,Varea E,Berezin V,et al.Chronic stress in adult⁃hood followed by intermittent stress impairs spatial memory and the survival of newborn hippocampal cells in aging animals:prevention by FGL,a pep⁃tide mimetic of neural cell adhesion molecule[J].Behav Pharmacol,2008,19(1):41-49.
[20]Skibo GG,Lushnikova IV,Voronin KY,Dmitrieva O,Novikova T,Klementiev B,et al.A synthetic NCAM-derived peptide,FGL,protects hippocam⁃pal neurons from ischemic insult bothin vitroandin vivo[J].Eur J Neurosci,2005,22(7):1589-1596.
[21]Pedersen MV,Helweg-Larsen RB,Nielsen FC,Berezin V,Bock E,Penkowa M.The synthetic NCAM-derived peptide,FGL,modulates the tran⁃scriptional response to traumatic brain injury[J].Neurosci Lett,2008,437(2):148-153.
[22] Hohsfield LA,Humpel C.Intravenous infusion of monocytes isolated from 2-week-old mice enhanc⁃es clearance of beta-amyloid plaques in an Al⁃zheimer mouse model[J].PLoS One,2015,10(4):e0121930.
[23]Kodali M,Parihar VK,Hattiangady B,Mishra V,Shuai B,Shetty AK.Resveratrol prevents age-re⁃lated memory and mood dysfunction with in⁃creased hippocampal neurogenesis and microvas⁃culature,and reduced glial activation[J].Sci Rep,2015,5:8075.
[24]Ojo B,Rezaie P,Gabbott PL,Cowely TR,Med⁃vedev NI,Lynch MA,et al.A neural cell adhesion molecule-derived peptide,FGL,attenuates glial cell activation in the aged hippocampus[J].Exp Neurol,2011,232(2):318-328.
[25] Hernangómez M,Carrillo-Salinas FJ,Mecha M, Correa F,Mestre L,Loría F,et al.Brain innate im⁃munity in the regulation of neuroinflammation:ther⁃apeutic strategies by modulating CD200-CD200R interaction involve the cannabinoid system[J].Curr Pharm Des,2014,20(29):4707-4722.
[26]Cox FF,Berezin V,Bock E,Lynch MA.The neural cell adhesion molecule-derived peptide,FGL,atten⁃uates lipopolysaccharide-induced changes in glia in a CD200-dependent manner[J].Neurosci⁃ence,2013,235(4):141-148.
[27] Wong WT.Microglial aging in the healthy CNS:phenotypes,drivers,and rejuvenation[J].Front Cell Neurosci,2013,7:22.
[28]Zhou X,Zöller T,Krieglstein K,Spittau B.TGFβ1inhibits IFNγ-mediated microglia activation and protects mDA neurons from IFNγ-driven neurotox⁃icity[J].J Neurochem,2015,134(1):125-134.
[29]Massobrio P,TessadoriJ, Chiappalone M,Ghirardi M.In vitrostudies of neuronal networks and synaptic plasticity in invertebrates and in mam⁃mals using multielectrode arrays[J].Neural Plast,2015:196195.
[30]Beesley PW,Herrera-Molina R,Smalla KH,Seidenbecher C.The neuroplastin adhesion mole⁃cules:key regulators of neuronal plasticity and syn⁃aptic function[J].J Neurochem,2014,131(3):268-283.
[31]Cambon K,Hansen SM,Venero C,Herrero AI,Skibo G,Berezin V,et al.A synthetic neural cell adhesion molecule mimetic peptide promotes syn⁃aptogenesis,enhances presynaptic function,and facilitates memory consolidation[J].J Neurosci,2004,24(17):4197-4204.
[32]Popov VI,Medvedev NI,Kraev IV,Gabbott PL,Davies HA,Lynch M,et al.A cell adhesion mole⁃cule mimetic,FGL peptide,induces alterations in synapse and dendritic spine structure in the den⁃tate gyrus of aged rats:a three-dimensional ultra⁃ structural study[J].Eur J Neurosci,2008,27(2):301-314.
[33]Hodges JL,Vilchez SM,Asmussen H,Whitmore LA,Horwitz AR.α-Actinin-2 mediates spine mor⁃phology and assembly of the post-synaptic density in hippocampal neurons[J].PLoS One,2014,9(7):e101770.
[34]Ojo B,Rezaie P,Gabbott PL,Davies H,Colyer F,Cowley TR,et al.Age-related changes in the hip⁃pocampus(loss of synaptophysin and glial-synap⁃tic interaction)are modified by systemic treatment with an NCAM-derived peptide,FGL[J].Brain Behav Immun,2012,26(5):778-788.
[35]KlementievB,NovikovaT,NovitskayaV,Walmod PS,Dmytriyeva O,Pakkenberg B,et al. A neural cell adhesion molecule-derived peptide re⁃duces neuropathological signs and cognitive im⁃pairment induced by Abeta25-35[J].Neurosci⁃ence,2007,145(1):209-224.
[36]Golpich M,Amini E,Hemmati F,Ibrahim NM,Rahmani B,Mohamed Z,et al.Glycogen syn⁃thase kinase-3 beta(GSK-3β)signaling:implica⁃tions for Parkinson′s disease[J].Pharmacol Res,2015,97:16-26.
[37]Park SJ,Jin ML,An HK,Kim KS,Ko MJ,Kim CM,et al.Emodin induces neurite outgrowth through PI3K/Akt/GSK-3β-mediated signaling pathways in Neuro2acells[J].NeurosciLett,2015,588:101-107.
[38]Huang P,Li C,Fu T,Zhao D,Yi Z,Lu Q,et al. Flupirtine attenuates chronic restraint stressinduced cognitive deficits and hippocampal apopto⁃sis in male mice[J].Behav Brain Res,2015,288:1-10.
[39]Development of a novel FGL therapy and transla⁃tional tests for regenerative treatment of neurologi⁃cal disorders.European Commission[EB/OL];(2012-01-01)http://cordis.europa.eu/search/index. cfm?fuseaction=proj.docum ent&PJ_RCN=12673448.
[40]Zellinger C,Salvamoser JD,Seeger N,Russmann V,Potschka H.Impact of the neural cell adhe⁃sion molecule-derived peptide FGL on seizure pro⁃gression and cellular alterations in the mouse kin⁃dling model[J].ACS Chem Neurosci,2014,5(3):185-193.
[41]Elsayed M,Banasr M,Duric V,Fournier NM,Licznerski P,Duman RS.Antidepressant effects of fibroblast growth factor-2 in behavioral and cellu⁃lar models of depression[J].Biol Psychiatry,2012,72(4):258-265.
[42]Turner CA,Gula EL,Taylor LP,Watson SJ,AkilH.Antidepressant-like effects of intracerebroven⁃tricular FGF2 in rats[J].Brain Res,2008,1224:63-68.
[43] Aonurm-Helm A,Jurgenson M,Zharkovsky T,Sonn K,Berezin V,Bock E,et al.Depressionlike behaviour in neural cell adhesion molecule(NCAM)-deficient mice and its reversal by an NCAM-derived peptide,FGL[J].Eur J Neurosci,2008,28(8):1618-1628.
[44]Erickson M,Hahn B,Leonard C,Robinson B,Luck S,Gold J.Enhanced vulnerability to distrac⁃tion does not account for working memory capaci⁃ty reduction in people with schizophrenia[J].Schizophr Res Cogn,2014,1(3):149-154.
[45]Secher T,Berezin V,Bock E,Glenthøj B.Effect of an NCAM mimetic peptide FGL on impairment in spatial learning and memory after neonatal phencyclidine treatment in rats[J].Behav Brain Res,2009,199(2):288-297.
[46]Ojo B,GabbottPL,Rezaie P,Corbett N,Medvedev NI,Cowley TR,et al.An NCAM mimet⁃ic,FGL,alters hippocampal cellular morphometry in young adult(4 month-old)rats[J].Neurochem Res,2013,38(6):1208-1218.
[47]Klein R,Blaschke S,Neumaier B,Endepols H,Graf R,Keuters M,et al.The synthetic NCAM mimetic peptide FGL mobilizes neural stem cellsin vitroandin vivo[J].Stem Cell Rev,2014,10(4):539-547.
[48]Wang J,Zheng J,Zheng Q,Wu Y,Wu B,Huang S,etal.FGL-functionalized self-assembling nanofiber hydrogel as a scaffold for spinal cord-de⁃rived neuralstem cells[J].Mater Sci Eng C Mater Biol Appl,2015,46:140-147.
Research progress in function and pharmacology of neural cell adhesion molecule-derived peptide FG loop
WANG Su-ping,GAO Yue,CHU Cheng-yan,LI Shen
(NO.1 Department of Neurology,Dalian Municipal Central Hospital,Dalian 116033,China)
Neural cell adhesion molecules(NCAMs)are able to promote neurogenesis,neuronal differentiation,and neuronal protection,playing a pivotal role in the regulation of the development and functions of the nervous system.FG loop(FGL),a bioactive polypeptide composed of 15 amino acids,is developed according to the spatial structure of NCAMs and binding site with fibroblast growth factor receptors.As the functional core segment of NCAMs,FGL has been proved to be neuroprotective,promote neuronal differentiation,have anti-inflammatory effects and regulate neural plasticity,which has a great potential in treatment of various neuropsychiatric diseases,such as ischemic brain injury,cognitive disorder,Alzheimer disease(AD)and depression.Furthermore,clinical researches have demonstrated that FGL is well tolerated in healthy male volunteers with no safety concerns.PhaseⅠclinical trial of FGL for AD is underway.FGL may be a potential therapeutic agent for treatment of human neuropsychiatric diseases in the future,which shows a promising application.
FGL;neural cell adhesion molecule;neuropsychiatric diseases;polypeptides
LI Shen,Tel:(0411)84418617,E-mail:listenlishen@hotmail.com
R971
A
1000-3002-(2016)08-0867-07
10.3867/j.issn.1000-3002.2016.08.011
Foundation item:The project supported by National Natural Science Foundation of China(81300985)
2015-04-15 接受日期:2015-08-27)
(本文编辑:乔 虹)
国家自然科学基金项目(81300985)
王苏平,男,硕士,主任医师,主要从事神经系统疾病的临床治疗及干细胞应用研究;李 深,女,博士,副主任医师,主要从事神经系统发育及再生研究。
李 深,E-mail:listenlishen@hotmail.com,Tel:(0411)84412001-8617
