HSPC238相互作用蛋白RPL5的初步筛选①
2016-01-08陈敬林谭家余钟裕恒万志丹
陈敬林 黄 湘 谭家余 钟裕恒 万志丹
(南方医科大学第三临床医学院 南方医科大学附属中山博爱医院,中山528403)
①本文受国家自然科学基金(81101534)、广东省自然科学基金(S2012010010824)和广东省医学科研基金(A2013875)项目资助。
②南方医科大学附属中山市博爱医院产前诊断中心,中山528403。
③南方医科大学附属中山市博爱医院中心ICU,中山528403。
[摘 要] 目的:构建HSPC238诱饵载体,筛选与HSPC238相互作用的目标蛋白。方法:基因合成法合成HSPC238基因,sfiIA和sfiIB双酶切后与pGBKT7诱饵载体连接,获得诱饵质粒pGBKT7-HSPC238,经测序鉴定后与酵母双杂交空质粒pGBKT7共同转化到酵母菌株AH109,在营养缺陷培养基中观察pGBKT7-HSPC238的自激活作用,进一步从人胎肝cDNA文库中筛选与HSPC238相互作用的目标蛋白。结果:诱饵载体pGBKT7-HSPC238构建成功,经表型筛选检测无自激活作用,经酵母双杂交技术结合文献分析,从人胎肝cDNA文库中初步筛选发现核糖体蛋白L5(Ribosomal protein L5,RPL5)可能是HSPC238相互作用的目标蛋白之一。 结论:成功构建pGBKT7-HSPC238诱饵质粒载体,且经酵母双杂交技术结合文献分析发现RPL5可能是与HSPC238相互作用的目标蛋白之一。
[关键词] HSPC238;酵母双杂交;核糖体蛋白L5;相互作用蛋白
doi:10.3969/j.issn.1000-484X.2015.11.011
·免疫学技术与方法·
HSPC238相互作用蛋白RPL5的初步筛选①
陈敬林黄湘②谭家余③钟裕恒②万志丹②
(南方医科大学第三临床医学院南方医科大学附属中山博爱医院,中山528403)
①本文受国家自然科学基金(81101534)、广东省自然科学基金(S2012010010824)和广东省医学科研基金(A2013875)项目资助。
②南方医科大学附属中山市博爱医院产前诊断中心,中山528403。
③南方医科大学附属中山市博爱医院中心ICU,中山528403。
[摘要]目的:构建HSPC238诱饵载体,筛选与HSPC238相互作用的目标蛋白。方法:基因合成法合成HSPC238基因,sfiIA和sfiIB双酶切后与pGBKT7诱饵载体连接,获得诱饵质粒pGBKT7-HSPC238,经测序鉴定后与酵母双杂交空质粒pGBKT7共同转化到酵母菌株AH109,在营养缺陷培养基中观察pGBKT7-HSPC238的自激活作用,进一步从人胎肝cDNA文库中筛选与HSPC238相互作用的目标蛋白。结果:诱饵载体pGBKT7-HSPC238构建成功,经表型筛选检测无自激活作用,经酵母双杂交技术结合文献分析,从人胎肝cDNA文库中初步筛选发现核糖体蛋白L5(Ribosomal protein L5,RPL5)可能是HSPC238相互作用的目标蛋白之一。 结论:成功构建pGBKT7-HSPC238诱饵质粒载体,且经酵母双杂交技术结合文献分析发现RPL5可能是与HSPC238相互作用的目标蛋白之一。
[关键词]HSPC238;酵母双杂交;核糖体蛋白L5;相互作用蛋白
doi:10.3969/j.issn.1000-484X.2015.11.011
中图分类号R392
文献标志码码A
文章编号号1000-484X(2015)11-1494-04
作者简介:陈敬林(1988年-),男,主要从事肿瘤免疫学相关研究,E-mail:842630809@qq.com。
通讯作者及指导教师:黄湘(1971年-),女,主任技师,硕士生导师,主要从事肿瘤免疫学相关研究,E-mail:340382761@ qq.com。
[Abstract]Objective:To construct a bait vector for HSPC238,and to screen the target proteins which interact with the HSPC238.Methods: Gene synthesis method was used to synthetic gene HSPC238,then connected with the pGBKT7 vector after digesting by the sfiIA and sfiIB,to obtain the bait plasmid pGBKT7-HSPC238,then transferred into the yeast strains AH109 with the empty plasmid pGBKT7after sequencing,to observe its self-activating effect in the nutrient deficiencies medium,and further to screen the target proteins which interact with HSPC238 from the human fetal liver cDNA library.Results: The bait vector pGBKT7-HSPC238 was successfully constructed,and it had no self-activating effect through the phenotypic screening,after the yeast two-hybrid technology with literature analysis,we preliminary screened and found that the ribosomal protein L5(RPL5) may be the one of the target proteins which interacted with HSPC238 from the human fetal liver cDNA library.Conclusion: We successfully constructed the bait plasmid vector PGBKT7-HSPC238,and after the yeast two-hybrid technology with literature analysis,we preliminary screened and found that the ribosomal protein L5(RPL5) may be the one of the target proteins which interacted with HSPC238.
Preliminary screening of RPL5 interacting with HSPC238
CHENJing-Lin,HUANGXiang,TANJia-Yu,ZHONGYu-Heng,WANZhi-Dan.TheThirdClinicalCollegeofSouthernMedicalUniversity,BoaiHospitalAffiliatedtoSouthernMedicalUniversity,Zhongshan528403,China
[Key words]HSPC238;Yeast two-hybridsy;RPL5;Interaction proteins
HSPC238 蛋白是由LOC51255 基因( NM_016494)编码的含153个氨基酸的,包含RING 指(RING-type zinc finger)结构域的蛋白质,属一种C3HC4 型锌指蛋白[1]。其特殊的结构提示该蛋白可能参与泛素蛋白酶体途径,并与细胞的生长有关,但目前国内外尚未见有关其功能的任何报道。运用UniGene数据库进行电子表达谱分析提示该基因广泛表达于人体各类组织中,在多种肿瘤组织中也均有表达。本研究旨在构建HSPC238酵母双杂交诱饵载体,并初步筛选与HSPC238相互作用的目标蛋白,为进一步系统研究HSPC238蛋白在肝癌发生发展中的作用机制奠定基础。
1材料与方法
1.1材料酵母双杂交系统的诱饵表达载体质粒pGBKT7、pGADT7酵母工程菌株AH109、人胎肝 cDNA文库均购自美国Clontech公司;宫颈癌Hela细胞株为本实验室保存;T4DNA连接酶购自美国Promega公司;限制性内切酶、MulV Reverse Transcriptase均购自加拿大Ferments公司;Trizol试剂购自美国Invitrogen公司;各种营养缺陷培养基及基础耗材购自南京钟鼎生物公司;DNA胶纯化试剂盒、质粒小提取试剂盒购自AXYGEN公司,引物合成、DNA测序由上海生工公司完成。
1.2方法
1.2.1载体构建前期用基因合成法合成HSPC238,基因合成时在其首末端同时合成sfiIA和sfiIB酶切位点的序列,构建PcDNA3.1-HSPC238质粒,转化大肠杆菌Top10感受态扩增目标基因并抽提质粒。同时对PcDNA3.1-HSPC238及诱饵载体pGBKT7质粒进行SfiI酶切,酶切产物琼脂糖电泳-割胶回收(Axygen胶回收试剂盒)对目的基因进行测序验证,以确保获取足量正确的目的基因片段。将回收的目的基因及诱饵载体进行连接(即构建pGBKT7-HSPC238载体),连接体系转化大肠杆菌感受态,随机挑取3个转化子接种于带卡那抗性的液体LB培养基,于37℃,250 r/min振荡培养16 h(过夜)后,行质粒抽提(Axygen质粒小量抽提试剂盒)并电泳,初步鉴定质粒pGBKT7和插入的目标片段。
1.2.2酵母转化取鉴定正确的pGBKT7-HSPC238载体和pGADT7共转化感受态酵母AH109菌株,涂布选择性平板(SD-Trp-Leu)培养3 d,同时单转pGBKT7-HSPC238载体涂布SD-Trp平板做对照。
1.2.3自激活检测 随机挑取5个共转化(pGBKT7-HSPC238+pGADT7)酵母转化子进行自激活检测,同时检测3个报告基因(LacZ,HIS和ADE2),三个报告基因均未出现自激活现象的转化子用于后续文库筛选。
1.2.4胎肝cDNA文库转化及阳性克隆筛选用含有正确pGBKT7-HSPC238载体且未出现自激活作用的AH109酵母菌作为受体菌制备感受态,将文库质粒(Matchmaker Human Fetal Liver Library;人胎儿肝脏cDNA酵母双杂交文库 Cat.No:638805)转入其中,涂SD-3+3AT平板。用绒布对筛库平板进行影印清除,以消除背景生长菌落的影响,继续培养1~2周。挑取再次长出的转化子进一步检测LacZ,HIS和ADE2三个报告基因,初步判断在这些菌落内的相互作用。
1.2.5阳性克隆质粒的抽提及BLAST比对将步骤1.2.4中可能存在相互作用的阳性菌落转接种于SD-Leu液体培养基,振荡培养过夜后抽提酵母质粒,转化大肠杆菌Top10新鲜感受态进行扩增培养。将含有阳性克隆的Top10转化子转接于含有Amp的LB液体培养基,扩增后抽提质粒,对质粒进行DNA测序,根据测序结果在www.ncbi.nim.nih.gov网站进行BLAST比对分析。
2结果
2.1pGBKT7-HSPC238诱饵载体的构建及鉴定将基因合成的含sfiIA和sfiIB酶切位点的HSPC238 基因与诱饵质粒载体pGBKT7 连接,获得重组诱饵质粒pGBKT7-HSPC238。转化大肠杆菌感受态后,随机挑取3个转化子接种于带卡那抗性的液体LB培养基,抽提质粒进行电泳检测。结果显示,pGBKT7-HSPC238的3个随机克隆经sfiI酶切都能得到目的条带,鉴定结果见图1。选取其中一个用T7引物进行测序进一步验证,测序及结果见图2。由此比对结果可以看出:包括sfiI酶切位点在内的序列都正常,未发生突变,用作后续文库筛选的实验。
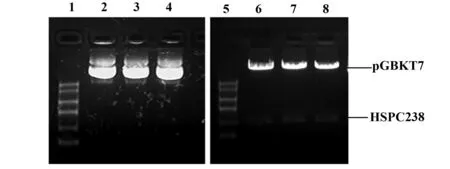
图1 pGBKT7-HSPC238诱饵载体的电泳鉴定图Fig.1 Electrophoresis for identification of pGBKT7-HSPC238 bait vectorNote: 1,5.Marker;2,3,4.pGBKT7-HSPC238 bait vector before enzyme digestion by sfiI;6,7,8.pGBKT7-HSPC238 bait vector after which enzyme digested with SfiI.

图2 插入片段的测序结果比对Fig.2 Gene sequencing and gene alignment for insertedfragment

表1 BLAST比对分析部分结果列表Tab.1 Partial results list of BLAST alignment analysis
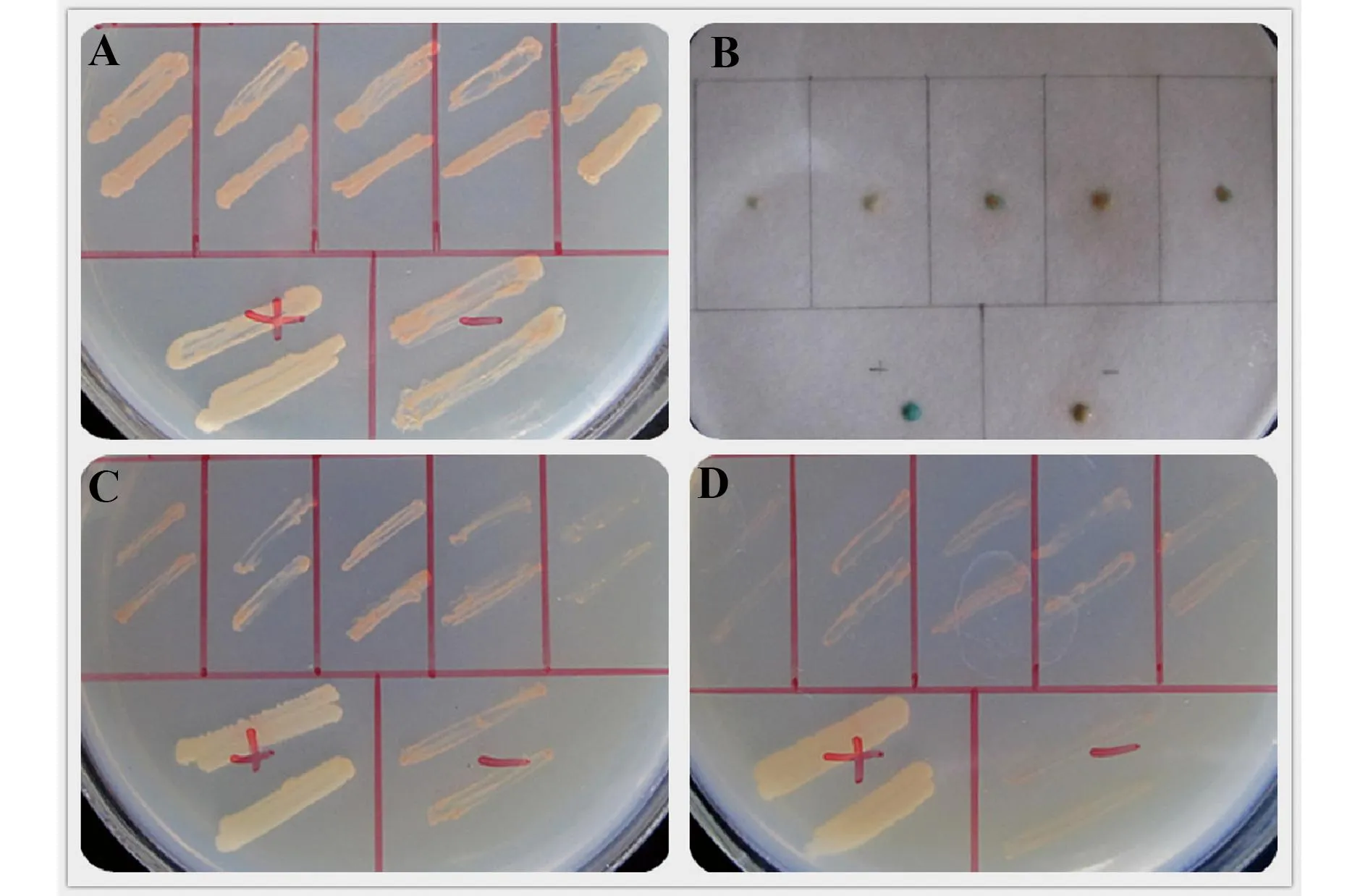
图3 诱饵载体的自激活及毒性检测图Fig.3 Self-activity and toxicity detection for bait vectorNote: The first line of each picture were the five randomly picked yeast transformants which containing pGBKT7-HSPC238 and pGADT7,the left in the second row of each picture were positive control(AD-T + BD-p53),the right were negative control(AD-T + BD-Lam).Fig A was the bacteria growth in the SD-TL plates,as the positive control.Fig B was the result of LacZ detection.Fig C and D were bacteria in SD-3 + 5 mmol/L 3AT and SD-4 plates.
2.2诱饵载体自激活检测鉴定正确的pGBKT7-HSPC238诱饵载体转化到酵母AH109菌株,随机挑取5个转化子进行自激活检测,分别同时检测3个报告基因(LacZ,HIS3和ADE2),如图3。检测结果表明,LacZ、HIS3和ADE2这三个报告基因均未出现自激活现象,因此该诱饵载体pGBKT7-HSPC238可用于后续文库筛选。
2.3cDNA文库转化及阳性克隆筛选文库质粒转化AH109感受态酵母菌后,在SD-3+3AT平板上继续培养2周,共长出124个转化菌落,挑选该124个阳性菌落检测LacZ,HIS和ADE2三个报告基因。结果表明:60个酵母菌落内无相互作用,45个酵母菌落内有较强相互作用,19个酵母菌落内有较弱的相互作用。
2.4阳性克隆质粒的抽提及BLAST比对对64个有较强和较弱相互作用的阳性菌落进行质粒抽提做进一步DNA测序和BLAST比对分析,结果列表如下(见表1)。经查阅文献,初步发现RPL5可能是与HSPC238相互作用的目标蛋白之一。
3讨论
Entrez PubMed 数据库提示HSPC238 基因编码的蛋白质含有一个锌指结构域,锌指结构是许多真核生物或病毒蛋白质所含有的保守的、富含半胱氨酸的、由40到60个氨基酸残基组成的区域[2]。既往研究提示,锌指结构域是泛素蛋白酶体途径中E3连接酶的重要活性区域,而许多含锌指结构的蛋白都可以在泛素蛋白酶体通路中扮演E3酶的角色[3,4]。另有研究表明含锌指结构的蛋白可参与转录调控,在肿瘤的发生、发展及侵袭中发挥作用[5-8]。由此可见,锌指结构域可能通过介导蛋白质之间的相互作用,参与信号转导、病毒复制等多种生物学途径,因而该结构域成为各种蛋白质发挥其重要功能的关键部位所在[9]。已有文献报道HSPC238蛋白具有E3酶活性[10],本课题组前期研究还表明,该蛋白质在肝癌中具有明显的抑癌作用[11],此外,再未检索到该蛋白的相关研究报道。然而对于该基因如何控制肝癌细胞的凋亡及其通过何种下游靶蛋白发生作用,其作用的靶蛋白有哪些?这些问题仍有待解决。酵母双杂交(yeast two-hybrid system)技术恰好为回答这些问题提供了切实可行的实验途径。
为此,本研究拟构建pGBKT7-HSPC238诱饵载体,利用酵母双杂交技术初步筛选与HSPC238相互作用的蛋白质分子。本研究中,pGBKT7-HSPC238的3个随机克隆经sfiI酶切都能得到目的条带(见图1)。同时对回收的插入片段进行测序比对,可以看到包括sfiI酶切位点在内的序列都正常(见图2),未发生突变,可用作后续实验。此外,由于某些蛋白质可直接激活酵母双杂交系统中的报告基因,从而出现假阳性结果,甚至某些蛋白质在酵母菌株内表达时对酵母菌可以产生一定的毒性作用,所以对诱饵蛋白自激活作用及毒力的检测是非常必要的。本研究在诱饵重组质粒pGBKT7-HSPC238转化感受态酵母菌株AH109的同时转化pGBKT7空载体作为对照来检测诱饵表达载体是否有自激活作用及毒性作用。结果表明,分别转化重组诱饵载体pGBKT7-HSPC238和空载体pGBKT7后的酵母菌AH109在SD/-Trp平板中生长出的单克隆数量大致相近,表明pGBKT7-HSPC238对AH109无毒性作用;同时含pGBKT7-HSPC238的AH109在SD/-Trp、SD/-His、SD/-Leu、SD/-Ura平板上的不同生长状况排除了自激活报告基因的作用,符合筛选cDNA文库的需要。
本研究进一步将胎肝cDNA文库转入上述未出现自激活作用的AH109酵母菌,涂SD-3+3AT平板,继续培养1~2周,获得了124个阳性转化菌落。挑取该阳性转化菌落进一步检测LacZ、HIS和ADE2三个报告基因,初步判断在这些菌落内的相互作用。挑取可能存在相互作用的64个菌落增菌培养后抽提质粒,做DNA测序和BLAST比对分析。查阅文献,初步发现核糖体蛋白L5(Ribosomal protein L5,RPL5)可能是与HSPC238相互作用目标蛋白之一,这为下一步的研究指明了方向。但由于通过酵母双杂交技术筛选获得的相互作用蛋白可能存在一定的假阳性结果,下一步本研究团队还需通过激光共聚焦、Pull-Down、免疫共沉淀等技术对HSPC238与RPL5的相互作用进行系统验证。
核糖体蛋白L5在正常细胞增殖中具有重要的作用,有研究表明,RPL5的遗失并不会诱导细胞周期的停滞,但由于核糖体的减少及蛋白翻译能力下降会抑制细胞分化周期的进程[11],近年来有学者发现RPL5的变异与先天性纯红细胞再生障碍性贫血相关[12,13],但RPL5与肝癌的关系目前尚未见相关文献报道,这也是本课题组后续研究的一个方向。
总之,本研究成功构建了HSPC238诱饵融合蛋白的表达载体pGBKT7-HSPC238,通过酵母双杂交技术及文献分析初步发现RPL5可能是与HSPC238相互作用的目标蛋白之一,为后续进一步深入研究HSPC238在肝癌中的作用机制奠定了的基础。
参考文献:
[1]黄湘,谭家余,邱玉林,等.pDsRed1-C3/LOC51255真核表达质粒的构建及在HePG_2细胞的表达定位[J].细胞与分子免疫学杂志,2009,25(5):402-405.
[2]Besold AN,Michel SL.Neural Zinc finger factor/myelin transcription factor proteins: metal binding,fold,and function[J].Biochemistry,2015,54(29):4443-4452.
[3]Hassink G,Kikkert M,Van Voorden S,etal.TEB4 is a C4HC3 RING finger-containing ubiquitin ligase of the endoplasmic reticulum[J].Biochem J,2005,388(Pt 2):647-655.
[4]Nakashima H,Nguyen T,Goins WF,etal.Interferon-stimulated gene 15 (ISG15) and ISG15-linked proteins can associate with members of the selective autophagic process,histone deacetylase 6 (HDAC6) and SQSTM1/p62[J].J Biol Chem,2015,290(3):1485-1495.
[5]Xie XM,Deng JY,Hou YC,etal.Evaluating the clinical feasibility: The direct bisulfite genomic sequencing for examination of methylated status of E3 ubiquitin ligase RNF180 DNA promoter to predict the survival of gastric cancer[J].Cancer Biomark,2015,15(3):259-265.
[6]Li Y,Yan X,Yan L,etal.High expression of Zinc-finger protein X-linked is associated with reduced E-cadherin expression and unfavorable prognosis in nasopharyngeal carcinoma[J].Int J Clin Exp Pathol,2015,8(4):3919-3927.
[7]Yuan L,Han J,Meng Q,etal.Muscle-specific E3 ubiquitin ligases are involved in muscle atrophy of cancer cachexia: an in vitro and in vivo study[J].Oncol Rep,2015,33(5):2261-2268.
[8]赵志虎,马清钧.锌指结构:最普遍的核酸识别元件[J].生物技术通讯,2001,12(1):36-41.
[9]Brophy TM,Raab M,Daxecker H,etal.RN181,a novel ubiquitin E3 ligase that interacts with the KVGFFKR motif of platelet integrin alpha(IIb)beta3[J].Biochem Biophys Res Commun,2008,369(4):1088-1093.
[10]Wang S,Huang X,Li Y,etal.RN181 suppresses hepatocellular carcinoma growth by inhibition of the ERK/MAPK pathway[J].Hepatology,2011,53(6):1932-1942.
[11]Teng T,Mercer CA,Hexley P,etal.Loss of tumor suppressor RPL5/RPL11 does not induce cell cycle arrest but impedes proliferation due to reduced ribosome content and translation capacity[J].Mol Cell Biol,2013,33(23):4660-4671.
[12]Quarello P,Garelli E,Carando A,etal.Diamond-Blackfan anemia: genotype-phenotype correlations in Italian patients with RPL5 and RPL11 mutations[J].Haematologica,2010,95(2):206-213.
[13]Delaporta P,Sofocleous C,Stiakaki E,etal.Clinical phenotype and genetic analysis of RPS19,RPL5,and RPL11 genes in Greek patients with Diamond Blackfan Anemia[J].Pediatr Blood Cancer,2014,61(12):2249-2255.
[收稿2015-05-20修回2015-07-20]
(编辑张晓舟)
