畜禽肌内脂肪相关基因的研究进展
2015-12-26郭月英王乐程海星张静任霆靳烨
郭月英,王乐,程海星,张静,任霆,靳烨
(内蒙古农业大学食品科学与工程学院,内蒙古呼和浩特010018)
畜禽肌内脂肪相关基因的研究进展
郭月英,王乐,程海星,张静,任霆,靳烨*
(内蒙古农业大学食品科学与工程学院,内蒙古呼和浩特010018)
肉质是肉用畜禽的重要经济指标,肌内脂肪含量是影响肉质指标如嫩度、多汁性和风味的重要因素。肌内脂肪的沉积是一个复杂的生理生化过程,这一过程受到新城代谢酶和肌肉生长及脂肪沉积的功能基因的调控。一些基因被证实是影响肌内脂肪的候选基因。心脏脂肪酸结合蛋白基因(H-FABP)、脂肪细胞脂肪酸结合蛋白(A-FABP)、脂蛋白脂酶(LPL)、生肌决定基因5(MFY5)和瘦素受体(LEPR)等基因是影响肌内脂肪的候选基因。
肉质;肌内脂肪;基因
近年来,我国逐渐成为肉类生产和消费的大国。肉类是人类膳食蛋白质的主要来源之一,随着经济的发展,消费者对肉质的关注程度日益提高,人们对肉的要求逐渐由量向质转变。肉质是一个综合性状,受多种因素的影响。肉质性状的形成从本质上讲是基因和环境相互作用的产物。研究表明,遗传因素即基因对肉质起着关键作用。肉质由多个基因控制,有研究显示一些基因对肉质起关键作用。伴随着生物技术的飞速发展,人类对基因结构和功能的认识和研究不断深入,肉质性状的遗传机制不断被发现,如何改善肉质成为世界各国研究的热点。
1 肉质
肉质是肉用家畜的重要经济指标。品种、饲养方式、营养水、宰前情况、宰后处理和遗传因素对肉质均会产生影响[1-3]。目前,评定肉质的主要指标一般包括嫩度、多汁性、肉色、风味、系水力、肌内脂肪含量(大理石纹)pH等[4]。
2 肌内脂肪与肉质
肌肉中的肌内脂肪(Intramuscular fat,IMF)主要存在于肌外膜和肌束膜,是具有中等偏高遗传力的重要经济性状。肌内脂肪是肉质特性的重要决定因素,已经有研究表明,肌内脂肪的含量及组成对肌肉的肉质性状具有重要的影响,肌内脂肪含量适中且分布均匀能使肌肉剪切力降低,使肌肉味美多汁、口感好且烹饪损失较低,即肌内脂肪可影响肉质的嫩度、多汁性和风味以及消费者的可接受性[5-9]。肌内脂肪对嫩度的改善主要体现于两个方面,一方面肌内脂肪氧化时可以溶解肌纤维束,进而提高肌肉嫩度和多汁性,另一方面是肌内脂肪一般沉积于结缔组织,肌内脂肪含量的增加,降低了结缔组织的含量,有利于肉质嫩度的提高。
影响IMF含量的因素很多,大量研究结果表明,遗传因素等是影响肌内脂肪沉积的一个重要因素。目前,已经证实一些基因与脂肪沉积相关。
3 影响肌内脂肪的基因
3.1 脂肪酸结合蛋白(FABP)基因
脂肪酸结合蛋白(FABPs)是脂蛋白(LBP)超家族中的成员,分子量为14 kDa~16 kDa,这些脂肪酸结合蛋白分别由脂肪酸结合蛋白基因编码[10-11]。在哺乳动物中,根据组织分布,分为9种不同的脂肪酸结合蛋白[12],基因名称和表达组织如表1所示。
在哺乳动物中有两种脂肪酸结合蛋白基因被作为IMF的候选基因,一种是心脏脂肪酸结合蛋白(heart fatty acid binding protein,H-FABP)基因,另一种是脂肪细胞型脂肪酸结合蛋白(adipocyte fatty acid binding protein,A-FABP)基因。
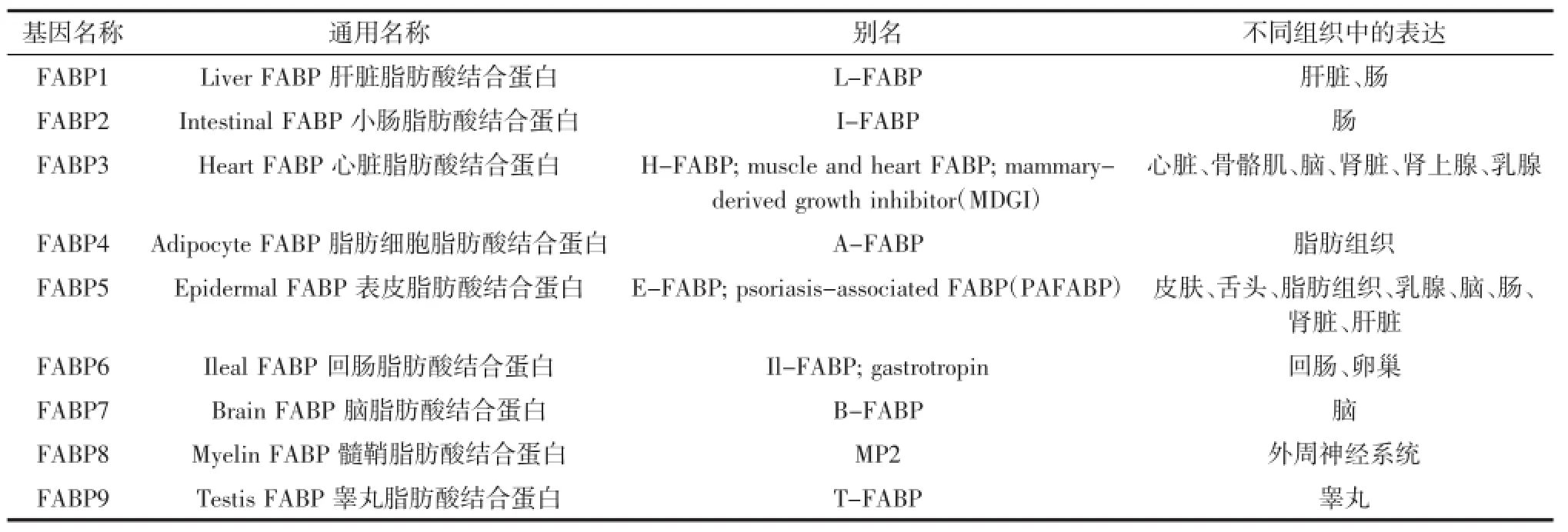
表1 各种脂肪酸结合蛋白基因Table1 Types of FABP genes
3.1.1 心脏脂肪酸结合蛋白基因(H-FABP)
心脏脂肪酸结合蛋白(H-FABP,又称FABP3),是FABP家族中分布最广的一个类型,是一个15kDa的蛋白质,在细胞内对长链脂肪酸的运输和代谢起关键作用,在肉质性状的形成中具有重要的生物学功能[11-12]。心脏脂肪酸结合蛋白由H-FABP基因编码,H-FABP基因在多种组织中均有表达,在心肌、骨骼肌和乳腺等中表达较多。是影响肌内脂肪的主要基因之一[13]。
心脏脂肪酸结合蛋白(H-FABP)与肌内脂肪的相关性在不同物种被研究和发现。国内外学者对猪做了大量的研究。Lee等2010年发现,H-FABP基因的HinfI酶切的HH基因型的肌内脂肪含量高于其它类型[13]。Li等2010年的研究证实,H-FABP基因的多态性影响猪的脂肪沉积并与背膘厚和IMF含量具有相关性[14]。Tyra等2011年发现了猪的H-FABP基因的多个多态位点,其中c.103CNT和c.1811GNC与腿肉的IMF含量存在相关性,多态位点c.103CNT与腰肉的IMF含量存在相关性[15]。杨文平2011年在对猪的HFABP基因的第1内含子的研究中发现,H-FABP基因的BB基因型个体与AA基因型个体间IMF含量差异显著[16]。Han等2012年在大白猪的研究发现中,HFABP基因的多态性与IMF显著相关,推断H-FABP基因的多态可作为基因标记对大白猪进行IMF的选育[17]。2012年王静在猪的研究中发现IMF含量与背最长肌H-FABP基因表达呈正相关[18]2013年Tyra等发现,H-FABP基因的表达量影响猪IMF含量但对腰肉和腿肉的皮下脂肪未产生影响[19]。江佳伟等2013年对滇南小耳猪的研究中发现,H-FABP的不同基因型影响猪肌内脂肪细胞中脂肪分解代谢相关基因的表达,其中HH基因型脂肪分解代谢相关基因的表达量较高,推断脂类代谢活动更强,从而相对增加了肌内脂肪沉积[20]。
在牛羊的研究中,也有类似的报道。李志才等2010年对湘西黄牛进行了研究,结果表明,H-FABP基因的多态性与IMF含量和大理石花纹间存在显著的相关性[21]。王兰萍等2012年在黄淮山羊的H-FABP基因的研究中发现了两种基因型,且证实在背最长肌和腿肌中,H-FABP基因的不同基因型对IMF含量有显著影响[22]。2012年郑程莉在对天府肉羊的研究中发现,H-FABP基因的表达量与IMF含量呈极显著正相关[23]。邓龙华等在2010年对鸡的H-FABP基因的3个多态位点的检测中发现,G+926A位点与胸肌粗脂肪含量呈极显著相关(P<0.01),与腿肌粗脂肪含量也有一定的相关趋势(0.05<P=0.063 2<0.1)[24]。
3.1.2 脂肪细胞脂肪酸结合蛋白(A-FABP)基因
A-FABP(又称FABP4或aP2)基因主要在脂肪细胞中表达。该基因在脂肪细胞内的脂类代谢和平衡起重要作用。研究表明,哺乳动物A-FABP是甘油三酯的贮存库。在甘油三酯形成及脂解过程中,A-FABP储存或释放大量脂肪酸,参与调控甘油三酷生成及溶解的生化循环[25]。A-FABP被认为是IMF侯选基因之一。
有一些研究表明,A-FABP基因与畜禽的肌内脂肪存在相关性。邝良德等在2010年对九龙牦牛进行了研究,发现A-FABP基因在背最长肌中的表达表达量与背最长肌的IMF含量存在显著相关性[26]。朱晓岩2012年在对肉牛的研究中发现,A-FABPmRNA表达水平和背最长肌IMF含量上存在互作效应;夏洛莱、利木赞和西门塔尔杂交肉牛的A-FABP的基因型影响活重和胴体重,、蒸煮损失、IMF含量、背膘厚和嫩度等;A-FABP基因的CC基因型与DD基因型肉牛的IMF含量存在显著差异[27]。
徐秋良2011年在绵羊的研究中发现小尾寒羊背最长肌中A-FABP基因表达量与背最长肌中IMF显著相关[28]。李祥辉2011年在4个不同品种的猪的研究中发现,A-FABP基因第一内含子的不同基因型个体间的IMF存在显著差异[29]。朱淑斌2013年研究发现,姜曲海猪和苏姜猪的A-FABP基因第1内含子的多态性与猪的大理石纹和肌内脂肪存在显著相关性[30]。Chen等2013年在对猪的研究中发现,A-FABP基因与猪的IMF的生长显著相关[31]。何俊2013年在对鸭的研究中发现,鸭A-FABP基因多态性与胸肌脂肪含量具有显著相关性[32]。
3.2 脂蛋白脂酶(Lipoprotein lipase,LPL)基因
脂蛋白脂酶(LPL)1943年被发现,是水解甘油三酯成不饱和脂肪酸和乳糜的限速酶,在脂肪的新城代谢中起着关键作用[33]。目前,已经有一些研究证实LPL基因与IMF的含量存在相关性。
姚飞2008年在对苏尼特肥羔的背最长肌、股二头肌中LPL活性发育规律的研究中发现,苏尼特肥羔羊肌肉组织中LPL是IMF沉积的重要参与者,对IMF的沉积有显著的影响[34]。曲桂娟2011年在牛的研究中发现,利杂牛和利草杂牛LPL表达量和IMF含量存在显著的正相关[35]。Ding等2012年在牦牛LPL基因的第七外显子发现一个新的单核苷酸多态位点(nt19913 C→T),该突变使编码的氨基酸由苯丙氨酸转变为丝氨酸,且该多态位点与屠宰性状和脂肪沉积相关[36]。2013年祝仁铸在野莱F1猪的研究中发现,猪LPL基因表达量与IMF呈显著正相关[37]。Wang等2013年在猪的研究中发现,LPL基因的表达量与单不饱和脂肪酸呈正相关而与饱和脂肪酸呈负相关[38]。Wu等2013年发现中国的雉鸡的LPL基因的第二外显子的碱基突变(c.6731 T→C)与鸡胸和鸡腿中的IMF含量呈现相关性,推断LPL基因可作为中国雉鸡IMF含量的候选基因[39]。2013王晶在对广西黄鸡的研究中发现,LPL基因的相对表达量、LPL酶活性与IMF含量的发育性变化之间存在一致的正相关[40]。
3.3 生肌决定基因5(MYF5)基因
生肌决定基因5(MYF5)属于生肌决定基因(MYOD)家族。MYF5基因时肌内脂肪含量及产肉性能的候选基因。
2012年,贾伟德在对中国的秦川牛、夏南牛、南阳牛、延边牛、郏县红牛和鲁西牛群体进行了研究,结果发现,Myf5(1142AG)突变类型与肌内脂肪含量,大理石花纹,系水力,眼肌面积和嫩度有显著相关[41]。Verner等2007年研究了254头,MyoD基因多态性与大白猪和长白猪的产肉性能的相关性,Myf5与肌内脂肪含量(IMF)和瘦肉率显著相关[42]。
2007年梁艳在对多个猪种的研究中发现MYF5的多个多态位点,其研究结果表明,MYF5基因的不同基因型与大理石评分、瘦肉率、眼肌面积等存在相关性。BB基因型有助于改善大理石纹评分,增加IMF含量;GG基因型也有助于增加IMF含量[43]。2008年Liu等对大白猪×梅山猪F2的研究表明,MYF5基因的第一外显子Hsp92II多态使编码的氨基酸发生了转变,导致蛋氨酸替代亮氨酸,并且该多态与肌内脂肪含量显著相关[44]。
3.4 瘦素受体(leptin receptor,LEPR)基因
瘦蛋白受体基因(Leptin recaptor,LEPR)是1995年发现并克隆的[45]。瘦素受体(LEPR)属I类细胞因子受体和信号传导家族,由瘦素受体基因编码,广泛分布于机体各组织和器官中。瘦素受体与瘦素(leptin)相互结合,与哺乳动物的食物摄入和能量代谢相关[46]。LEPR影响脂肪代谢和脂肪沉积,是影响肌内脂肪的候选基因。
有研究表明,LEPR基因影响家畜的肌内脂肪。2010年Li等的研究发现,LEPR AvaII的酶切多态性与猪的肌内脂肪存在显著相关性[14]。Tyra等2011年的研究发现,在猪的胴体及主要分割肉中,LEPR基因的表达量与脂肪含量显著正相关[15]。2012年付言峰等的研究表明,LEPR基因在苏钟猪的多个组织中均有表达,且背膘中的表达量最高,LEPR是苏钟猪肉质改良育种中潜在的重要候选基因[47]。2013年,Tyra等对不同猪种的不同月龄的LEPR基因的转录本丰度与皮下脂肪和肌内脂肪的相关性进行了研究,结果显示,LEPR基因的表达量随着月龄的增长显著升高,并且LEPR基因的转录本的水平与个体的脂肪含量存在显著相关性[19]。
2011年,黄爱霞在对鸡的研究中发现,胸肌中LEPR基因表达量与肌内脂肪的含量呈中等程度的正相关[48]。
4 展望
肉质是一个重要的经济性状,肉类生产和销售企业以及消费者对肉质关注使得科研工作者对肉质的改善成为一项迫切的任务。脂肪的合成与分解代谢过程直接由脂肪合成酶和分解酶的含量与活性共同调控,此外,激素和脂肪细胞因子间接地调控脂肪代谢。现代生物技术的发展及其应用使一些与肌内脂肪(IMF)相关的基因被发现,影响IMF沉积基因及其影响机制的进一步研究,从而建立一个多基因协调调控网,为肉质的改善提供新的途径。从遗传基因的角度提高肉的品质,将在生产实践中产生巨大的经济效益。
[1]赵珺,李金泉,张文广,等.影响山羊肉质差异候选基因的研究进展[J].安徽农业科学,2013,41(16):7180-7181
[2]孟丽云.反刍家畜骨骼肌肉质特性及其关联基因的初步研究[D].呼和浩特市:内蒙古农业大学,2013
[3] Grunert K G,Bredahl L,Brunsø K.Consumer perception of meat quality and implications for product development in the meat sector—a review[J].Meat Science,2004,66(2):259-272
[4]闫秋良,金海国,赵玉民,等.不同杂交组合育肥羔羊生长,屠宰性能和肉品质的研究[J].云南农业大学学报:自然科学版,2013,28 (1):69-72
[5] Wood JD,Enser M,Fisher A V,et al.Fat deposition,fatty acid composition and meat quality:A review[J].Meat Science,2008,78 (4):343-358
[6]陈香凝.黑毛和牛肌内前体脂肪细胞分化的基因表达与肌内脂肪性能的关联性研究[D].扬州:扬州大学,2012
[7] 张宏宇,单安山,徐林,等.猪肌内脂肪调控研究进展[J].东北农业大学学报,2010(2):156-160
[8]Purchas RW,Burnham D L,Morris ST.Effects of growth potential and growth path on tenderness of beef longissimus muscle from bulls and steers[J].J Anim Sci,2002,80(12):3211-3221
[9]OkeudoN J,Moss BW.Interrelationships amongst carcass and meat quality characteristics of sheep[J].Meat Science,2005,69(1):1-8
[10]Chen QM,Wang H,Zeng Y Q,et al.Developmental changes and effect on intramuscular fat content of H-FABP and A-FABP mRNA expression in pigs[J].Journal of applied genetics,2013,54(1):119-123
[11]Yin H D,Wang Y,Zhao X L,et al.Effect Analysis of Heart Fatty Acid-Binding Protein Gene Assisted Selection on Intramuscular Fat Content in Chicken[J].J Anim.Vet.Adv,2012,11(10):1595-1600
[12]Chmurzyńska A.The multigene family of fatty acid-binding proteins (FABPs):function,structure and polymorphism[J].Journal of applied genetics,2006,47(1):39-48
[13]Lee SH,Choi YM,Choe JH,et al.Association between polymorphisms of the heart fatty acid binding protein gene and intramuscular fat content,fatty acid composition,and meat quality in Berkshire breed[J].Meat science,2010,86(3):794-800
[14]LiX,Kim SW,Choi JS,et al.Investigation of porcine FABP3 and LEPR gene polymorphisms and mRNA expression for variation in intramuscular fat content[J].Molecular biology reports,2010,37(8): 3931-3939
[15]Tyra M,Ropka-Molik K.Effect of the FABP3 and LEPR gene polymorphisms and expression levels on intramuscular fat(IMF) content and fat cover degree in pigs[J].Livestock Science,2011,142 (1):114-120
[16]杨文平,李彩桃,高爽,等.猪H-FABP基因内含子1的遗传多态性及其遗传效应分析[J].农业科学与技术:英文版,2011,12(6): 887-889
[17]Han X,Jiang T,Yang H,et al.Investigation of four porcine candidate genes(H-FABP,MYOD1,UCP3 and MASTR)for meat quality traits in Large White pigs[J].Molecular biology reports,2012,39(6): 6599-6605
[18]王静.猪肉品质的品种差异与营养调控及其机制研究[D].四川农业大学,2012
[19]Tyra M,Ropka-Molik K,Terman A,et al.Association between subcutaneous and intramuscular fat content in porcine ham and loin depending on age,breed and FABP3 and LEPR genes transcript abundance[J].Molecular biology reports,2013,40(3):2301-2308
[20]江佳伟,黄英,杨明华,等.H-FABP不同基因型对滇南小耳猪脂肪分解代谢相关基因表达的影响[J].畜牧兽医学报,2013,44(6): 837-842
[21]李志才,易康乐.湘西黄牛的H-FABP基因对大理石花纹和肌内脂肪含量相关性分析[J].中国牛业科学,2010,36(1):1-4
[22]王兰萍,耿荣庆,冀德君,等.黄淮山羊H-FABP基因多态性及其与肌内脂肪含量的关联分析[J].云南农业大学学报:自然科学版, 2012,27(4):522-525
[23]郑程莉.天府肉羊H-FABP,L-FABP和LPL基因生物信息学与组织表达分析[D].四川:四川农业大学,2012
[24]邓龙华,谢亮,罗成龙,等.鸡心脏型脂肪酸结合蛋白(H-FABP)基因多态性对肉质性状和组织表达的影响[J].农业生物技术学报, 2010,18(3):545-555
[25]Hertzel A V,Bennaars-Eiden A,Bernlohr D A.Increased lipolysis in transgenic animals overexpressing the epithelial fatty acid binding protein in adipose cells[J].Journal of lipid research,2002,43(12): 2105-2111
[26]邝良德,林亚秋,徐亚欧,等.九龙牦牛脂肪细胞型脂肪酸结合蛋白基因(A-FABP)的克隆及其表达谱[J].农业生物技术学报, 2010,18(6):1098-1102
[27]朱晓岩.营养和遗传因素对杂交肉牛IMF含量及其主效基因表达的影响[D].长春:吉林大学,2012
[28]徐秋良,张庆莉,陈玉林.绵羊脂肪细胞型脂肪酸结合蛋白基因(FABP4)cDNA的克隆,表达及其结构模拟分析[J].农业生物技术学报,2011,19(3):483-489
[29]李祥辉,黄大鹏,黄玉兰.4个不同品种猪A-FABP基因多态性及其与IMF相关性研究[J].中国畜牧杂志,2011,47(7):11-14
[30]朱淑斌,赵旭庭,周春宝,等.姜曲海猪A-FABP基因内含子1多态性与肉质性状的相关分析[J].中国畜牧杂志,2013,49(13): 12-15
[31]Chen QM,Wang H,Zeng YQ,et al.Developmental changes and effect on intramuscular fat content of H-FABP and A-FABPmRNA expression in pigs[J].Journal of applied genetics,2013,54(1):119-123
[32]何俊.鸭L-FABP,A-FABP基因多态性与肌内脂肪含量的相关性及其对脂肪代谢相关基因表达的影响[D].杭州:浙江大学, 2013
[33]Sagoo G S,Tatt I,Salanti G,et al.Seven lipoprotein lipase gene polymorphisms,lipid fractions,and coronary disease:a HuGE association review and meta-analysis[J].American journal of epidemiology,2008,168(11):1233-1246
[34]姚飞,卢德勋,姚焰础,等.苏尼特肥羔肌肉组织LPL和HSL的发育性变化及其对肌内脂肪沉积的影响[J].饲料工业,2008,29(5): 37-40
[35]曲桂娟,杨连玉,秦贵信,等.不同杂交品种肉牛背最长肌LPL基因表达的发育性变化及其对IMF的影响[J].中国兽医学报, 2011,31(2):272-274
[36]Ding X Z,Liang CN,Guo X,et al.A novel single nucleotide polymorphism in exon 7 of LPL gene and its association with carcass traits and visceral fat deposition in yak (Bos grunniens)steers[J]. Molecular biology reports,2012,39(1):669-673
[37]祝仁铸,尹逊河,王元虎,等.猪肌肉组织MDH和LPL基因表达与肌内脂肪含量和脂肪酸组成关系的研究[J].畜牧兽医学报, 2013,44(8):1182-1188
[38]Wang W,Xue W,Jin B,et al.Candidate gene expression affects intramuscular fat content and fatty acid composition in pigs[J].Journal ofapp lied genetics,2013,54(1):113-118
[39]Wu Q,Xing X M,Rong M,et al.Discovery of a SNP in exon 2 the lipoprotein lipase gene and its association with intramuscular fat content in Chinese ring-necked pheasant[J].Journal of Animal &Plant Sciences,2013,20(2):3122-3125
[40]王晶.广西三黄鸡肉质性状分析及LPL、H-FABP基因表达与肌内脂肪含量的相关研究[D].南宁:广西大学,2013
[41]贾伟德.牛MyoD基因家族多态性及其与肉质性状的关联性分析[D].杨凌:西北农林科技大学,2012
[42]Verner J,Humpolíček P,Knoll A.Impact of MYOD family genes on pork traits in Large White and Landrace pigs[J].Journal of Animal Breeding and Genetics,2007,124(2):81-85
[43]梁艳.猪Myf-5基因和L-FABP基因的遗传多态性及其与胴体和肉质性状间关联性研究[D].雅安市:四川农业大学,2007
[44]Liu M,Peng J,Xu D Q,et al.Association of MYF5 and MYOD1 gene polymorphisms and meat quality traits in Large White×Meishan F2 pig populations[J].Biochemical genetics,2008,46(11/12): 720-732
[45]Tartaglia L A,Dembski M,Weng X,et al.Identification and expression cloning of a leptin receptor,OB-R[J].Cell,1995,83(7):1263-1271
[46]Ernst CW,Kapke P A,Yerle M,et al.The leptin receptor gene (LEPR)maps to porcine chromosome6[J].Mammalian genome,1997, 8(3):226-226
[47]付言峰,方晓敏,李碧侠,等.瘦素受体表达量在苏钟猪脂肪沉积调控中的作用[J].中国兽医学报,2012,32(9):1266-1271
[48]黄爱霞.浙江省主要鸡种肉品质分析及Leptin receptor基因对鸡脂肪代谢影响分子机制的研究[D].杭州:浙江大学,2011
Development of IMF-related Genes in Livestock and Poultry
GUO Yue-ying,WANG Le,CHENG Hai-xing,ZHANG Jing,REN Ting,JIN Ye*
(College of Food Science and Engineering,Inner Mongolia Agricultural University,Huhhot,010018,Inner Mongolia,China)
Meat quality is an important economic trait in livestock and poultry.IMF content is an important determinant of meat quality characteristics such as tenderness,juiciness and flavor.IMF deposition was a complicated physiological and biochemical process which is regulated by metabolic enzymes and controlled by functional genes in the growth and development process of muscle and adipose tissue.Some genes influencing IMF had been studied,including H-FABP gene,A-FABP gene,LPL gene,MFY5 and LEPR gene.
:meat quality;intramuscular fat;gene
10.3969/j.issn.1005-6521.2015.01.033
2014-09-10
国家自然基金项目(31360393)
郭月英(1977—),女(汉),讲师,博士研究生,研究方向:畜产品加工与贮藏。
*通信作者
