乳酸菌胞外多糖生理功能及合成途径的研究进展
2015-11-07秦晓萌张远森柳陈坚张昆林
秦晓萌,张远森,柳陈坚,张昆林,杨 恩
(昆明理工大学,生命科学与技术学院,云南昆明650500)
乳酸菌胞外多糖生理功能及合成途径的研究进展
秦晓萌,张远森,柳陈坚,张昆林,杨恩*
(昆明理工大学,生命科学与技术学院,云南昆明650500)
乳酸菌(Lactic acid bacteria,LAB)是一类重要的益生微生物,同时也是一类重要的工业发酵菌,在食品发酵生产上应用广泛,它的代谢产物也受到极大关注。本文总结了乳酸菌代谢合成的胞外多糖(Exopolysaccharide,EPS)的主要生理生化功能以及胞外多糖合成的四个主要过程,以期为乳酸菌EPS的应用、产量提高及结构改造提供指导。
乳酸菌,胞外多糖,生理功能,代谢途径
乳酸菌(Lactic acid bacteria,LAB)是一类能发酵糖类产生乳酸的细菌的总称,目前发现的这类细菌在分类学上至少有23个属,其中在工业生产上应用较多的有7个属[1],包括链球菌属(Streptococcus),乳杆菌属(Lactobacillus),明串珠菌属(Leuconostoc),双歧杆菌属(Bifidobacterium),片球菌属(Pediococcus)等。乳酸菌在食品行业中应用广泛,与人体健康关系密切,具有调节肠道菌群平衡、免疫调节、抗肿瘤等多种生理活性[1],有学者认为上述生理活性与细胞表面多糖分子相关[2]。乳酸菌胞外多糖(Exopolysaccharide,EPS)是一种次生代谢产物,与细胞连接松散或直接分泌到环境中[3]。胞外多糖可分为粘液多糖和荚膜多糖,荚膜多糖和粘液多糖之间没有明确的界限,细菌荚膜多糖经常会失去与细胞膜的结合而溶解到溶液中[4]。乳酸菌EPS的分子量通常在4.0×104~6.0×106u之间[3],可分为同多糖和杂多糖。同多糖仅由一种单糖所构成,一般为葡萄糖或果糖。而杂多糖则是由多种单糖构成,单糖先聚合为3~8个单体构成的寡糖重复单元,再聚合成为多糖[5]。乳酸菌代谢产生的EPS是一种安全的天然产物,通常是作为食品添加剂和增稠剂使用[6-7]。
目前具有商业价值的乳酸菌多糖是由一种肠膜明串珠菌合成的同多糖——右旋糖苷,可以用于凝胶过滤的组分和血浆的替代品[8]。但是由于乳酸菌EPS的产量过低,提取成本高,无法满足工业规模的需求[9]。如果将EPS用作食品添加剂,那么EPS发酵生产的浓度需要达到10~15g/L,而目前已报道的最高生产浓度是2.767g/L[5]。然而,现代分子生物学和基因工程技术的不断发展以及比较基因组学等方法应用于乳酸菌的相关研究,为提高乳酸菌EPS产量提供了有效的理论基础和技术手段。
1 乳酸菌胞外多糖的生理功能
1.1乳酸菌EPS的粘附作用。
EPS可以作为一种粘附因子,帮助乳酸菌在肠道黏膜上定植[10],具有改善人体肠道环境的功能[4]。EPS还能选择性促进有益菌的增殖,进而抑制有害菌的增殖和有害物质的产生[11]。当前,比较基因组学和功能基因组研究发现人乳中的特定的糖对婴幼儿肠道微生物的发展起到了重要的作用[12]。然而,细菌胞外多糖的粘附作用也会产生一定的负面影响,例如变异链球菌(S.mutans)能够合成一种葡聚糖(Glucan),使菌体黏附在牙齿的表面腐蚀牙釉质而导致龃齿[13]。
1.2乳酸菌EPS的抗氧化作用
乳酸菌EPS具有抗氧化活性并且具有低细胞毒性已得到广泛的报道[14],有研究认为多糖的抗氧化性质与分子量相关,分子量越小抗氧化活性越强[15]。Lai等[16]对分离自高粱酒糟泡菜的产EPS乳酸菌的抗氧化性质进行了研究,发现短乳杆菌A3(L.brevis A3)、植物乳杆菌B2(L.plantarum B2)、短乳杆菌C9(L.brevis C9)和短乳杆菌D7(L.brevis D7)四种乳酸菌的完整细胞和无细胞提取物均具有抗氧化活性。在对羟自由基、DPPH和脂质过氧化的分析中,L.brevis D7的完整细胞和无细胞提取物均具有最好的氧化清除活性,而同时L.brevis D7的胞外多糖产量也最高,说明EPS对乳酸菌的抗氧化性质具有重要作用。将动物双歧杆菌RH(B.animalis RH)所产EPS分离纯化,得到一种主要的胞外多糖EPSa,并发现浓度为0.2mg/mL时,EPSa片段对DPPH自由基的清除活性与0.25mg/mL的抗坏血酸相当;而EPSa对羟自由基和超氧阴离子自由基的清除活力、脂质超氧化的抑制能力都要明显优于抗坏血酸[17]。
1.3乳酸菌EPS的免疫调节作用
乳酸菌EPS具有免疫调节作用,但并不是所有的多糖都具有免疫活化功能,这表明多糖的理化特征与其生物功能特性密切相关。EPS的结构特性与免疫调节性质的关系包括以下两个方面[18]:含有负电荷侧链结构的酸性杂多糖具有良好的诱导免疫反应的作用;杂多糖的体积或分子量大就会抑制免疫反应的发生。
乳酸乳球菌乳脂亚种KSV20(Lactococcus lactis subsp.cremoris KSV20)是一种广泛用于斯堪的纳维亚发酵酸奶viili生产的菌种,该菌合成一种杂多糖能够诱导体外培养的鼠脾巨噬细胞产生IFNγ和IL-1α。这种多糖结构中的吡喃型半乳糖侧链被磷酸基团所取代[19],多糖中的酸性片段能够有效地诱导巨噬细胞的增殖和活化。将杂多糖中的磷酸基团脱去后发现多糖的免疫刺激减弱,从而证明了磷酸基是一种免疫反应的分子激活物[20]。除了磷酸基团外,多糖中含有硫酸基团也可以刺激免疫细胞分泌出细胞素类物质。嗜盐四联球菌KK221(Tetragenococcus halophilus KK221)合成的岩藻聚糖(fucoidan)[21](有26.7%硫酸基团),通过比较完整多糖和脱硫酸化多糖的免疫刺激效应发现,在含有硫酸基团的状态下具有良好的免疫激活作用,而脱硫酸化多糖的免疫激活作用下降。多糖的免疫激活作用与硫酸基团有关,而与多糖链的长度无关,这说明硫酸基团对上调细胞素类物质是必需的。
乳酸菌多糖分子量与免疫调节作用的关系也有一系列的报道。鼠李糖乳杆菌ATCC9595(L.rhamnosus ATCC9595)及其变异菌株鼠李糖乳杆菌RW-9595M(L.rhamnosus RW-9595M)产生的低分子量EPS和高分子量EPS分别刺激老鼠腹腔粘膜的巨噬细胞,L.rhamnosus ATCC9595合成的低分子量的EPS可以诱导巨噬细胞产生高水平的TNF、IL-6和IL-12,而高分子量的EPS诱导产生的TNF和IL-6水平较低[22]。
此外,乳酸菌的免疫调节作用还具有菌株差异性,即不同的乳酸菌及计量可以引起不同的免疫反应。一些乳酸菌主要是诱导发生先天性免疫反应,包括抑制炎症应答、调节Toll样受体(Toll-like receptors,TLRs)的表达、活化自然杀伤细胞(Nature killer cells,NK cells)、活化树状细胞(Dendritic cells,DCs)和其他抗原呈递细胞(Antigen presenting cells,APC cells);另一些乳酸菌则诱导后天免疫反应,包括促进淋巴细胞增殖、胃肠道特异性的IgA的产生以及Th1/Th2应答的平衡[23]。
目前乳酸菌细胞表面糖类物质的免疫调节作用已经得到普遍认同,未来研究的重点是了解乳酸菌与宿主在体内的互作以及这些表面免疫物质的分子基础[24]。
1.4乳酸菌的抗肿瘤作用
乳酸菌具有抗肿瘤活性,特别是对结直肠癌具有很好的预防作用。一些研究对来自于乳酸菌的组分的抗肿瘤活性进行分析,发现来自于乳酸菌的肽聚糖、多糖片段和糖蛋白都具有抗肿瘤作用[25]。Chong[11]认为乳酸菌的抗癌效应机制包括以下几个方面:乳酸菌能够利用胞外多糖粘附在肠道黏膜表面,竞争性抑制有害菌的附着,改善肠道的菌落结构;改变肠道菌落的酶活性,抑制与致癌物产生相关的酶活性,包括β-葡糖苷酸酶(β-glucuronidase)、β-葡糖苷酶(β-glucosidase)、硝基还原酶(Nitroreductase)和偶氮还原酶(Azoreductase)。这些酶类催化产生致癌物,可以导致结直肠癌的发生;乳酸菌缺少7α-脱羟化酶(7α-dehydroxylase)活性,可以减少具有致癌作用的次级胆酸(脱氧胆酸Deoxycholic acid,DCA;石胆酸Lithocholic acid,LCA)的产生,EPS有利于机体中胆酸的排除。再者乳酸菌EPS及肽聚糖还可以结合体内的N-亚硝基组分和黄曲霉毒素等致癌物、致突变剂,预防由DNA损伤诱导的癌变发生。
乳酸菌作为一类广泛利用的微生物,不仅具有抗氧化、抗肿瘤、抗炎和免疫调节[26]等重要的生理活性,并具有分泌产糖的特性。这使得乳酸菌一直是工业生产、医药保健等领域研究开发的热点之一。而乳酸菌EPS由于其多方面的作用,且不同于其他的食品添加剂,是一种原位发酵的天然产物,因而被认为是一种安全的食品物质(General Recognised As Safe status,GRAS)[3],受到越来越多的关注。但乳酸菌合成EPS的产量普遍较低,不能满足工业生产的需求,还不能成为动物性多糖和植物性多糖的替代品[27]。因此,需要对乳酸菌胞外多糖合成相关的基因和合成途径进行全面研究,使基因工程操作成为可能。
2 乳酸菌EPS的合成
2.1乳酸菌杂多糖(Heteropolysaccharides,HePS)的合成
EPS的合成过程是复杂的,有大量的基因参与这个过程。目前了解还不全面,但可以确定EPS的合成途径分为四个过程[6],包括:将参与反应的单糖转运进入细胞质中;葡糖-1-磷酸的合成;糖的活化和链接聚合;EPS的输出过程。
EPS合成中编码相关的酶和调节蛋白基因的座位是不同的,有的乳酸菌的基因位于质粒上,而另一些乳酸菌的基因则位于染色体上[6]。乳酸菌产生胞外多糖的能力也是不稳定的,这与细胞中遗传物质的不稳定性有关。
2.1.1参与反应的单糖转运进入细胞质单糖的转运系统分为三类:初级转运系统:通过转运特异性ATP酶将单糖的转运与ATP的水解偶联起来,利用ATP水解获得的能量实现单糖的运输;二级转运系统:这一系统中单糖的转运不与ATP的水解偶联,而是与离子或其他溶质的同向或反向跨膜运输相偶联;烯醇式丙酮酸-糖特异性磷酸转移酶系统(PEPPTS)。在这一系统中,磷酸烯醇式丙酮酸(PEP)提供能量,糖特异性的磷酸转移酶系统(PTS)负责单糖的转运[7]。
将单糖转运进入细胞质的最常见机制是烯醇式丙酮酸-糖特异性磷酸转移酶系统(PEP-PTS)[28]。PEP-PTS系统包括一系列蛋白,分别负责底物连接、跨膜转运和底物的磷酸化。PEP-PTS系统中第一类蛋白包括酶I(即包含组氨酸残基的磷酸载体蛋白HPr)和酶II(即糖特异性的透过酶复合物)[29]。如图1所示[6],首先磷酸烯醇式丙酮酸(PEP)将磷酸基团传递给酶I,酶I的组氨酸残基接受磷酸基团,HPr转变为磷酸化的HPr(His-P)[28]。同时酶II复合物与糖单体结合,负责糖的跨膜运输并将HPr(His-P)上的磷酸基团转移到糖单体上[30]。在单糖的运输及磷酸化的过程中,PEP是磷酸基团的供体,单糖是磷酸基的最终受体。微生物不同,PEP-PTS系统的组成也可能不同,L.lactis的乳糖转运PEP-PTS系统的酶II由三种蛋白构成,分别是:酶IIA、酶IIB、酶IIC[31]。
第二类蛋白具有调节营养物质吸收的功能。目前最清楚的调节蛋白是由一种代谢调控蛋白CcpA和PTS载体蛋白HPr形成的一种二聚体蛋白[32]。HPr有His和Ser两个磷酸化位点。CcpA蛋白与Ser磷酸化的HPr(Ser-P)结合,抑制活性单糖的转运[33]。
2.1.2葡糖-1-磷酸的合成细胞质中磷酸化单糖的代谢取决于其磷酸化的状态,葡糖-6-磷酸进入分解途径,葡糖-1-磷酸参与多糖的合成。通过PEPPTS系统进入细胞的一般是葡糖-6-磷酸。而在葡糖-6-磷酸转变为葡糖-1-磷酸的过程中,磷酸葡糖变位酶(PGMs)发挥了重要作用。PGMs对糖的代谢具有调控作用[34-35]。细胞中存在两种磷酸葡糖变位酶(PGMs):α-PGM和β-PGM。如图1所示,β-PGM的作用是将β-葡糖-1-磷酸转变为葡糖-6-磷酸;而α-PGM则是将葡糖-6-磷酸转变为α-葡糖-1-磷酸并用于合成EPS[34-35]。但是用葡萄糖培养L.lactis时,葡萄糖必须先转换为葡糖-6-磷酸再用于EPS的合成[36]。除了上述产生α-葡糖-1-磷酸的途径外,还有一些替代途径,这些途径的不同之处在于糖原和转运系统的差异[37-38]。
2.1.3糖核苷酸的合成与EPS重复单元的聚合此过程所需的基因分为两组:核苷糖生物合成基因和EPS合成特异性基因。
第一组基因与核苷糖的合成有关,核苷糖用于重复单元的合成,主要有UDP-glucose、UDP-galactose和dTDP-rhamnose[6]。核苷糖合成完成,进入EPS合成阶段。这一阶段的酶是EPS特异性酶,属于第二组基因的编码产物。这些基因的组成在不同细胞内相对保守,由四个互不重叠的结构域组成,如图2所示[5]。在革兰氏阳性细菌中,EPS特异性基因簇依次为调节序列、链长决定序列、重复单元合成序列、聚合及输出序列[5]。
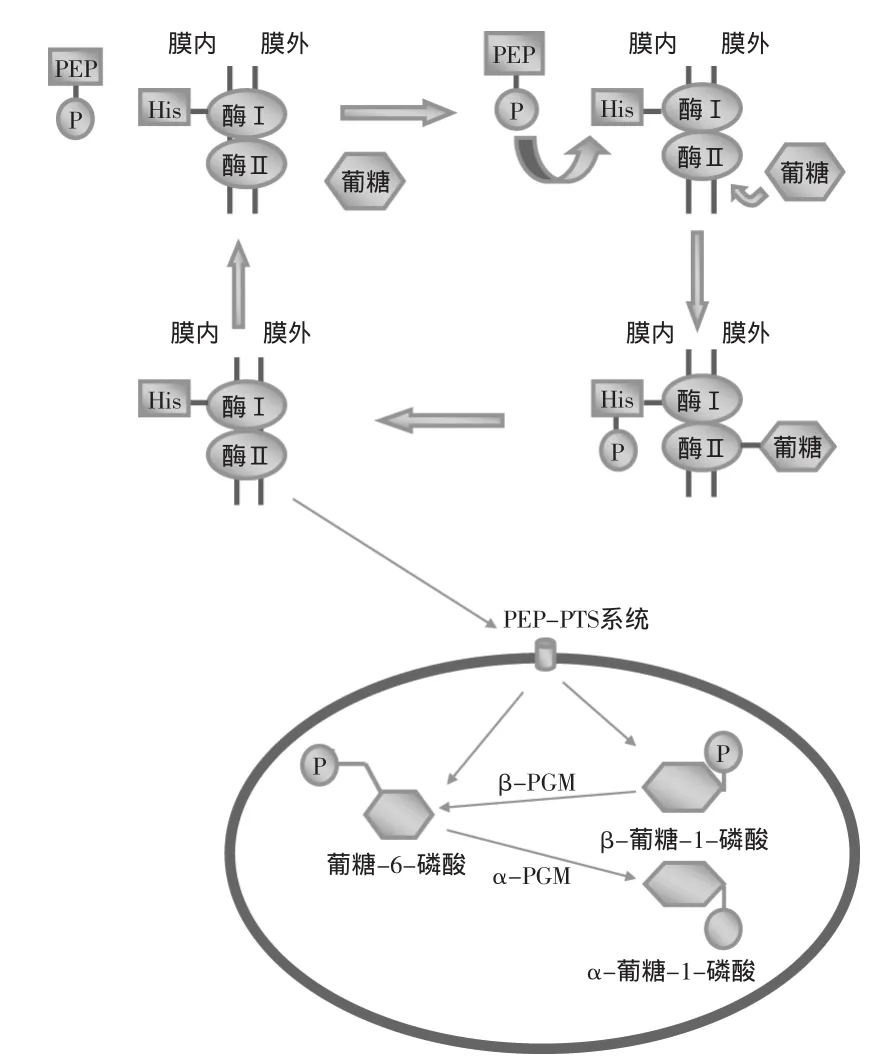
图1 PEP-PTS系统对糖单体的转运示意图Fig.1 PEP-PTS system diagram of sugar monomer transfer
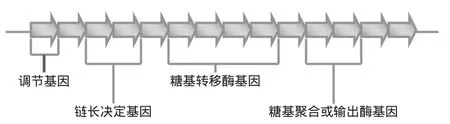
图2 EPS特异性基因分布示意图Fig.2 Distribution of EPS-specific genes
重复单元的合成是在细胞质膜的内表面完成的。首先在特异性糖基转移酶作用下,UDP-糖作为糖基供体将糖转移到脂质载体(十一异戊烯磷酸,Und-P)上形成脂载体焦磷酸化糖(Und-PP-糖)。然后通过各种不同的糖基转移酶将不同的糖依次转移到脂质载体上,形成脂载体焦磷酸化糖重复单元(Und-PP-糖重复单元)。最后在翻转酶作用下将Und-PP-糖重复单元转移到细胞外[39]。
2.1.4EPS的聚合及分泌自然界中多糖的种类及结构虽然千变万化,但它们的合成途径相对一致,即在特异性糖基转移酶作用下将单糖依次转移到脂质载体上形成寡糖重复单元,再转移到胞外并聚合为多糖。重复单元的起始、聚合和输出是多糖合成的主要步骤,根据这些步骤的差异可将这一过程分为三种类型,即Wzy依赖途径、ABC转运依赖途径和合成酶依赖途径[40]。
Wzy依赖途径是O-抗原合成的主要途径,并作为多糖合成的模型[41]。在糖基转移酶作用下,脂载体焦磷酸化糖(Und-PP-糖)不断延伸形成脂载体焦磷酸化重复单元(Und-PP-糖重复单元),Und-PP-糖重复单元在翻转酶Wzx作用下输出到细胞外;然后通过聚合酶Wzy形成一个长链的O-抗原;Wzz在O-抗原聚合过程中控制寡糖重复单元的数量,即决定了糖链的长度。
现在对重复单元的聚合方式以及参与整个过程的酶还不完全了解。但是可以概括出一个简单的EPS聚合和转运的模型。首先在翻转酶作用下,脂载体焦磷酸-糖重复单元(Und-PP-糖重复单元)从膜的内表面转移到外表面,然后聚合酶将重复单元连接为一个长链,最后酶催化脂载体和糖链的分离并起到控制链长的作用。
2.2乳酸菌同多糖(Homopolysaccharides,HoPS)的合成
在乳酸菌细胞中同多糖的合成要简单许多。同多糖的合成发生在细胞外[3],由特异性糖基转移酶即葡聚糖-蔗糖酶(Glycansucrases)或果聚糖-蔗糖酶(Fructansucrase)催化合成。在细胞外,乳酸菌通过Glycansucrases利用蔗糖作为底物来合成同多糖,多糖合成过程所需的能量也来自蔗糖的水解[13]。同多糖合成不需要其他能量,因为其过程中不需要将糖类转运进入细胞内,也不需要将单糖转化为活性核苷糖形式,因此乳酸菌能很容易将蔗糖转变为同多糖。由蔗糖转化为HoPS的反应如下[13]:

葡聚糖、果聚糖是最为常见的同多糖[13],其中果聚糖中主要包括β-2,6糖苷键和β-2,1糖苷键;葡聚糖中主要通过α-1,6糖苷键、α-1,3糖苷键、α-1,4糖苷键和α-1,2糖苷键与α-D-葡糖基团连接,而β-D-葡聚糖主要以β-1,3键和β-1,2键连接。由于HoPS的合成过程较为简单,对能量要求不高,所以HoPS既可以在培养基中合成,也可以在无细胞的酶液中合成[42]。
3 结论及展望
乳酸菌胞外多糖是一种安全的微生物次生代谢产物,不仅能够改善乳制品的口感和风味,还具有抗氧化、抗肿瘤、抗炎症和免疫调节等显著的益生作用,应用前景广阔。
乳酸菌EPS作为一种粘附因子有助于乳酸菌在肠道中粘附定植,进而竞争性抑制有害菌的附着增殖,减少有害菌数量,同时也减少了有害物质的产生。EPS具有很好的抗氧化作用,这种抗氧化特性与分子量的大小关系密切,同时乳酸菌EPS具有很低的细胞毒性的优点,是一种潜在的优良抗氧化物质。乳酸菌EPS还具有免疫调节作用,这种作用与EPS的来源,种类和结构有关。通常情况下含有负电荷侧链的酸性多糖以及分子量越小的EPS具有较好的免疫调节作用。不同的乳酸菌对免疫系统的激活也具有差异性,有些主要诱导先天性免疫,另一些则诱导后天免疫。这种免疫激活的菌种特异性的分子基础还有待进一步研究。乳酸菌EPS具有抗肿瘤活性,这一特性与乳酸菌的粘附作用、抗氧化作用和免疫调节作用紧密相连。抗氧化活性可以抑制机体的氧化性损伤,防止DNA损伤诱发的癌变;免疫调节作用可以提高机体免疫系统活性,抑制癌细胞的增殖和转移,促进癌细胞的凋亡;EPS的附着吸附作用一方面能够帮助菌体定植,增加EPS体内留存时间以加强其作用效果,另一方面也可以吸附有害有毒成分,减小对细胞的破坏。
乳酸菌EPS的生物合成机制是胞外多糖研究的一个重要方面,可以为EPS的定向改造提供理论依据和指导方向。当前乳酸菌EPS的合成机制还并不十分清楚完善,但可以确定乳酸菌合成胞外多糖的四个主要步骤,包括:单糖分子进入细胞质;葡糖-1-磷酸的合成转化;糖的活化与寡糖重复单元的合成;EPS的聚合与输出。其中杂多糖的聚合过程较为复杂,组分与结构多样,并且主要过程发生于细胞质中,需要ATP等提供能量;而同多糖的合成过程相对简单,主要分为葡聚糖和果聚糖,底物通常是蔗糖,在细胞外合成,能量来源于蔗糖中糖苷键的水解。因此,乳酸菌同多糖的产量一般要高于杂多糖的产量。鉴于国内外现状,多糖合成机理的研究有限,重点主要集中在合成条件的研究筛选。
现在人们对于乳酸菌和大型可食用真菌(如香菇、灵芝)的多糖类物质的研究不断深入,对它们的特性也更加了解。这促使更多的人将目光集中在多糖的研究上,大量的新型菌株不断被发现,更多的多糖被分离出来。陆地上的极端生境、海洋、极地以及太空都是具有巨大潜力的微生物种源地,其中大量有价值的微生物还有待开发。未来的研究重点在于多糖结构与功能的关系。多糖的结构复杂,包括单体的种类、单体数量、连键类型以及侧链的差异。目前多糖的结构与功能关系并不清楚,什么样的结构具有抗氧化作用,什么种类的侧链能够赋予多糖抗肿瘤特性等,弄清其结构功能的关系有助于多糖产品的开发、改性与定向改造。
[1]张筠,刘宁,孟祥晨.乳酸菌胞外多糖生物学活性[J].国外医学卫生学册,2004,31(4):227-230.
[2]Fanning S,Hall JL,Cronin M,et al.Bifidobacterial surfaceexopolysaccharide facilitates commensal-host interaction through immune modulation and pathogen protection[J].PNAS,2012,109(6):2108-2113.
[3]Ruas-Madiedo P,Hugenholtz J,Zoon P.An overview of the functionality of exopolysaccharides produced by lactic acid bacteria[J].Int Dairy J,2002,12:163-171.
[4]Costerton JW,Irvin RT,Cheng KJ,et al.The role of bacterial surface structures in pathogenesis[J].Crit Rev Microbiol,1981,8(4):303-338.
[5]Welman AD,Maddox IS.Exopolysaccharides from lactic acid bacteria:perspectives and challenges[J].Trends Biotechnol,2003,21(6):269-274.
[6]Laws A,Gu Y,Marshall V.Biosynthesis,characterization,and design of bacterial exopolysaccharides from lactic acid bacteria[J].Biotechnol Adv,2001,19(8):597-625.
[7]De Vuyst L,De Vin F,Vaningelgem F,et al.Recent developmentsinthebiosynthesisandapplicationsof heteropolysaccharides from lactic acid bacteria[J].Int Dairy J,2001(11):687-707.
[8]Badel S,Bernardi T,Michaud P.New perspectives for lactobacilli exopolysaccharides[J].Biotechnol Adv,2011,29(1):54-66.
[9]Nwodo UU,Green E,Okoh AI.Bacterial exopolysaccharides:functionality and prospects[J].Int J Mol Sci,2012,13(11):14002-14015.
[10]Vélez MP,De Keersmaecker SC,Vanderleyden J.Adherence factors of Lactobacillus in the human gastrointestinal tract[J]. FEMS Microbiol Lett,2007,276(2):140-148.
[11]Chong ES.A potential role of probiotics in colorectal cancer prevention:review of possible mechanisms of action[J].World J Microbiol Biotechnol,2014,30(2):351-374.
[12]Zivkovic AM,German JB,Lebrilla CB,et al.Human milk glycobiome and its impact on the infant gastrointestinal microbiota[J].PNAS,2011,108:4653-4658.
[13]Monsan P,Bozonnet S,Albenne C,et al.Homopolysaccharides from lactic acid bacteria[J].Int Dairy J,2001,11:675-685.
[14]Pan D,Mei X.Antioxidant activity of an exopolysaccharide purified from Lactococcus lactis subsp.lactis 12[J].Carbohyd Polym,2010,80:908-914.
[15]Chen H,Zhang M,Qu Z,et al.Antioxidant activities of different fractions of polysaccharide conjugates from green tea(Camellia Sinensis)[J].Food Chem,2008,106:559-563.
[16]Lai Y,Tsai S,Lee M.Isolation of Exopolysaccharide Producing Lactobacillus Strains from Sorghum Distillery Residues Pickled Cabbage and their Antioxidant Properties[J].Food Sci Biotechnol,2014,23(4):1231-1236.
[17]Xu R,Shen Q,Ding X,et al.Chemical characterization and antioxidant activity of an exopolysaccharide fraction isolated from Bifidobacterium animalis RH[J].Eur Food Res Technol,2011,232:231-240.
[18]Hidalgo-Cantabrana C,López P,Gueimonde M,et al. Immune modulation capability of exopolysaccharides synthesized by lactic acid bacteria and bifidobacteria[J].Probiotics Antimicro Prot,2012,4:227-237.
[19]Nakajima H,Hirota T,Toba T,et al.Structure of the extracellular polysaccharide from slime-forming Lactococcus lactis subsp.cremoris SBT 0495[J].Carbohyd Res,1992,224:245-253.
[20]Kitazawa H,Harata T,Uemura J,et al.Phosphate group requirement for mitogenic activation of lymphocytes by an extracellularphosphopolysaccharidefromLactobacillus delbrueckii ssp.bulgaricus[J].Int J Food Microbiol,1998,40(3):169-175.
[21]Kawashima T,Murakami K,Nishimura I,et al.A sulfated polysaccharide,fucoidan,enhances the immunomodulatory effects of lactic acid bacteria[J].Int J Mol Med,2012,29(3):447-453.
[22]Bleau C,Monges A,Rashidan K,et al.Intermediate chains of exopolysaccharides from Lactobacillus rhamnosus RW-9595M increase IL-10 production by macrophages[J].J Appl Microbiol,2010,108(2):666-675.
[23]Tsai YT,Cheng PC,Pan TM.The immunomodulatory effects of lactic acid bacteria for improving immune functions and benefits[J].Appl Microbiol Biotechnol,2012,96(4):853-862.
[24]Wells JM.Immunomodulatory mechanisms of lactobacilli[J]. Microb Cell Fact,2011,10:1-15.
[25]Lee do K,Jang S,Kim MJ,et al.Anti-proliferative effects of Bifidobacterium adolescentis SPM0212 extract on human colon cancer cell lines[J].BMC Cancer,2008,8:1-8.
[26]Saad N,Delattre C,Urdaci M,et al.An overview of the last advances in probiotic and prebiotic field[J].LWT-Food Sci Technol,2013,50:1-16.
[27]Canquil N,Villarroel M,Bravo S,et al.Behavior of the rheological parameters of exopolysaccharides synthesized by three lactic acid bacteria[J].Carbohyd Polym,2007,68:270-279.
[28]PostmaPW,LengelerJW,JacobsonGR.Phosphoenolpyruvate:carbohydrate phosphotransferase systems of bacteria[J].Microbiol Rev,1993,57(3):543-594.
[29]Viana R,Monedero V,Dossonnet V,et al.Enzyme I and HPr from Lactobacillus casei:their role in sugar transport,carbon catabolite repression and inducer exclusion[J].Mol Microbiol,2000,36(3):570-584.
[30]Sliz P,Engelmann R,Hengstenberg W,et al.The structure of enzyme IIAlactosefrom Lactococcus lactis reveals a new fold and points to possible interactions of a multicomponent system[J]. Structure,1997,5(6):775-788.
[31]Wang G,Louis JM,Sondej M,et al.Solution structure of the phosphoryl transfer complex between the signal transducing proteinsHPrandIIAGlucoseoftheEscherichiacoli phosphoenolpyruvate:sugar phosphotransferase system[J].EMBO J,2000,19(21):5635-5649.
[32]Van den Bogaard PT,Kleerebezem M,Kuipers OP,et al. Control of lactose transport,β-galactosidase activity,and glycolysis by CcpA in Streptococcus thermophiles:evidence for carbon catabolite repression by a non-phosphoenolpyruvate-dependent phosphotransferase system sugar[J].J Bacteriol,2000,182(21):5982-5989.[33]Suárez CA,Blancato VS,Poncet S,et al.CcpA represses the expression of the divergent cit operons of Enterococcus faecalis through multiple cre sites[J].BMC Microbiol,2011(11):1-13.
[34]Sjöberg A,Hahn-Hägerdal B.β-glucose-1-phosphate,a possible mediator for polysaccharide formation in maltoseassimilating Lactococcus lactis[J].Appl Environ Microbiol,1989,55(6):1549-1554.
[35]Degeest B,De Vuyst L.Correlation of activities of the enzymes α-phosphoglucomutase,UDP-galactose 4-epimerase,and UDP-glucose pyrophosphorylase with exopolysaccharide biosynthesis by Streptococcus thermophilus LY03[J].Appl Environ Microbiol,2000,66(8):3519-3527.
[36]Ramos A,Boels IC,De Vos WM,et al.Relationship between glycolysis and exopolysaccharide biosynthesis in Lactococcus lactis[J].Appl Environ Microbiol,2001,67(1):33-41.
[37]Grobben GJ,Smith MR,Sikkema J,et al.Influence of fructose and glucose on the production of exopolysaccharides and the activities of enzymes involved in the sugar metabolism and the synthesis of sugar nucleotides in Lactobacillus delbrueckii subsp. Bulgaricus NCFB 2772[J].Appl Microbiol Biot,1996,46:279-284.
[38]De Vos WM,Vaughan EE.Genetics of lactose utilization in lactic acid bacteria[J].FEMS Microbiol Rev,1994,15(2-3):217-237.
[39]丹彤,王俊国,张和平.乳酸菌胞外多糖的结构、生物合成及其应用[J].食品科学,2013,34(7):335-339.
[40]陈蕾蕾,王未名,祝清俊,等.细菌多糖的生物合成机制[J].微生物学报,2010,50(12):1583-1589.
[41]Whitfield C,Valvano MA.Biosynthesis and expression of cell-surface polysaccharides in gram-negative bacteria[J].Adv Microb Physiol,1993,35:135-246.
[42]姚晶,任婧,吴正钧,等.乳酸菌胞外多糖的生物合成及其遗传调控[J].食品与发酵工业,2011,37(2):11-16.
Progress on the physiological functions and synthesis pathways of lactic acid bacteria exopolysaccharide
QIN Xiao-meng,ZHANG Yuan-sen,LIU Chen-jian,ZHANG Kun-lin,YANG En*
(Kunming University of Science and Technology,Faculty of Life Science and Technology,Kunming 650500,China)
Lactic acid bacteria(LAB)were important probiotic microorganisms and critical industry bacteria,which were widely used in fermentation foods.The fermentation productions which produced by LAB were attracted great attention.This paper provided an overview of the main physiological and biochemical functions of exopolysaccharide(EPS)which produced by LAB.Besides,four metabolic pathways which related to exopolysaccharide biosynthesis also had been summarized,in order to provide evidence for the EPS applications,raising yields and structure modification.
lactic acid bacteria;exopolysaccharide;physiological functions;metabolic pathways
TS201.2
A
1002-0306(2015)14-0389-06
10.13386/j.issn1002-0306.2015.14.072
2014-10-28
秦晓萌(1990-),男,硕士研究生,研究方向:食品微生物。
杨恩(1982-),女,博士,讲师,研究方向:食品微生物。
云南省昆明理工大学人才培养项目(14118480);昆明理工大学分析测试基金(20140707)。
