春季季风间期巽他陆架和马六甲海峡表层海水浮游植物群落结构研究
2015-10-22王磊冷晓云孙庆杨王东晓张翠霞孙军
王磊,冷晓云,孙庆杨,王东晓,张翠霞,孙军*
(1.天津科技大学海洋科学与工程学院,天津300457;2.中国科学院南海海洋研究所热带海洋环境国家重点实验室,广东广州510301)
春季季风间期巽他陆架和马六甲海峡表层海水浮游植物群落结构研究
王磊1,2,冷晓云1,孙庆杨2,王东晓2,张翠霞1,孙军1*
(1.天津科技大学海洋科学与工程学院,天津300457;2.中国科学院南海海洋研究所热带海洋环境国家重点实验室,广东广州510301)
于2013年3-5月通过走航取样分别对巽他陆架和马六甲海峡表层海水浮游植物叶绿素a生物量和群落结构进行了观测和研究。结果表明:巽他陆架生物量较低,叶绿素a浓度平均值为(0.083 ±0.043)μg/L,爪哇海的SS4站位生物量最低,仅为0.014μg/L,浮游植物粒级组成上主要以Pico-级为优势,占80%以上;马六甲海峡自西北至东南存在明显的盐度梯度,在盐度最低的S M 5站,叶绿素a生物量最高,达到1.080μg/L;马六甲海峡站位叶绿素a浓度平均值为(0.433±0.315)μg/L,同时浮游植物群落结构变动较大。在海峡西北的S M 1-S M4站与巽他海峡类似,主要以聚球藻为优势类群,Pico-级浮游植物占60%~80%;在生物量最高的S M 5站,同样以聚球藻为优势类群,而在海峡东南段的S M6和S M7站,虽然叶绿素a浓度相对于S M 5略有降低,但仍明显高于其他马六甲海峡站位和巽他陆架站位,此两个站位硅藻比例明显升高,均可达20%以上。从优势类群生物量与环境因子和营养浓度的相关性可以看出,研究海区叶绿素a生物量与水体盐度呈现显著负相关(p<0.050),说明陆源输入对研究海区生物量具有明显的影响。另外,硅藻生物量也与磷酸盐浓度(p<0.050)和硅酸盐(p<0.010)浓度均呈现显著正相关;聚球藻在浮游植物群落中的优势度会受到陆源营养盐输入的影响而降低,但仍然是整个研究区域最优势的浮游植物类群。
浮游植物;群落结构;巽他陆架;马六甲海峡
王磊,冷晓云,孙庆杨,等.春季季风间期巽他陆架和马六甲海峡表层海水浮游植物群落结构研究[J].海洋学报,2015,37(2):120—129,doi:10.3969/j.issn.0253-4193.2015.02.013
Wang Lei,Leng Xiaoyun,Sun Qingyang,et al.The distribution of phytoplankton com munity structurein the Sunda Shelf and the Strait of M alacca during spring intermonsoon[J].Haiyang Xuebao,2015,37(2):120—129,doi:10.3969/j.issn.0253-4193.2015.02.013
1 引言
南海是典型的热带-亚热带陆架边缘海,位于其西南部的巽他陆架(Sunda Shelf)面积约为1.85×106k m2,平均水深基本低于50 m,是全球岛屿最多的陆架生态系统[1]。马六甲海峡(Strait of M alacca)在南海与印度洋的表层水交换上所起作用虽远不如Karimata海峡和Gelasa海峡[2],但因其狭长具有漏斗形状且作为海上交通要道的特殊性,一直以来也是备受瞩目的重要研究区域。巽他陆架和马六甲海峡受到典型的季风气候影响,一般5-9月盛行西南季风,潮湿多雨;11月至翌年3月盛行东北季风,气候干燥,因而在3-5月属于季风转换期[3]。由于巽他陆架及外围区域处于太平洋与印度洋海水交换的关键区域,印度尼西亚贯穿流等会受到季风影响[4],因此在季风期和季风转换期对于该区域的生物地球化学研究具有特别高的关注度[5]。
关于巽他陆架[6—8]及马六甲海峡[9—11]浮游植物群落结构和生物量时空变化已有大量的研究工作。柯志新等[6]对巽他陆架的叶绿素a粒级结构研究表明,在春末夏初叶绿素a浓度较低,在0.05~0.58μg/L之间,其中0.7~3μm粒级的叶绿素a约占75%。研究者通过对环境因子的分析认为盐度和硝酸盐含量是影响该区域叶绿素a浓度的主要因素。Ke等[7]的研究中指出,夏初巽他陆架浮游植物优势物种主要是原甲藻属(Prorocentrum)、膝沟藻属(Gonyaulax)、环沟藻属(Gyrodinium)、斯氏藻属(Scrippsiella)和角毛藻属(Chaetoceros)的种类,硅藻种类主要分布在低温、高营养盐的区域,而甲藻种类反之。由此可见在巽他陆架即使是在季风转换期,浮游植物生物量和物种组成上也存在较大的空间差异性。对马六甲海峡的浮游植物研究主要集中于新加坡近岸。据Gin等[9]研究报道,新加坡近岸周年平均叶绿素a浓度约为1.7μg/L,高生物量主要出现在西南季风期,其中Pico-级及小型的Nano-粒级的叶绿素a约占40%左右。Gin等[10]通过特征色素的方法发现,新加坡近岸区域硅藻支持了约72%的叶绿素a生物量,而聚球藻(Synechococcus)约占18%。通过上述研究结果可以看出,在巽他陆架和马六甲海峡,浮游植物群落结构和生物量存在着较大的时空变动。对于巽他陆架全粒级浮游植物群落组成和生物量的研究尚显不足,对于马六甲海峡的研究目前还缺乏跨整个海峡的环境梯度下的浮游植物群落结构变化的数据。本文正是基于以上设想开展了现场观测和研究。
2 材料与方法
2.1航次与站位
于2013年3-5月期间,乘中国科学院南海海洋研究所“实验一号”船参加国家自然科学基金委员会东印度洋科学考察航次,分别于3月16-19日在巽他陆架、5月3-4日在马六甲海峡进行走航表层海水取样观测,共11个测站(图1)。
2.2水文及营养盐参数
走航海表面温度(T)、盐度(S)等基本水文学参数均由SBE 21(SE A-BIR D ELEC T R O NIC,IN C.)于现场进行测定(见表1)。表层海水营养盐样品采用QuA Atro连续流动分析仪进行测定,具体参照Strickland和Parsons的方法进行[12]。

表1 采样站位信息及各测站表层海水温度、盐度Tab.1 Sampling date,time,location,the sea surface temperature and the sea surface salinity of all stations
2.3浮游植物特征光合色素
走航过程中采表层海水2~4 L,以直径25 m m的W hatman GE/E滤膜过滤后液氮保存。应用反相高效液相色谱法(H PLC)对滤膜样品进行定性、定量分析[13—14],光合色素标准品购于丹麦D HI公司(D K-2970 H ørsholm,Den mark),参照标准光合色素扫描图谱对环境样品进行定性定量分析[13]。根据Uitz等[15]给出的公式进行基于特征色素的浮游植物粒级结构的分析。
2.4C H E M TAX反演浮游植物群落结构
选取13种浮游植物光合色素,利用化学分类法软件[16]对浮游植物功能群进行反演,共分成甲藻、硅藻、定鞭金藻_8型、定鞭金藻_6型、绿藻、隐藻、原绿球藻、聚球藻和青绿藻9大浮游植物类群。
3 结果
3.1走航表层温度和盐度的分布特征
走航表层温度和盐度分布如图2所示,其中右侧轨迹为3月份从北至南的走航,左侧轨迹为5月份从西至东的走航。可以看出,3月和5月两次走航,巽他陆架表层海水温度相差2~3℃左右,而盐度基本相近,5月份略高。而在两次走航期间,马六甲海峡和爪哇海(the Java Sea)的盐度均明显低于巽他陆架。
通过对两段航迹温度-盐度道具(T-S properties)进行分析(表1和图3),可以看出,巽他陆架采样点温度在29.02~30.26℃之间,平均为(29.47±0.49)℃;盐度在30.54~32.84之间,平均为(32.14±0.93),以爪哇海的SS4站的高温低盐特征最为明显。相比而言,马六甲海峡各站位温度差别不大,平均为(30.81±0.13)℃;盐度在30.11~31.67之间,平均为(31.00±0.50),马六甲海峡西北部的S M 1-S M 4号站盐度明显较高,而S M 5 -S M 7号站盐度均低于31。从T-S图可以看出,巽他陆架除SS4站位外,与马六甲海峡站位水体温、盐性质差异较大。SS4站与马六甲海峡东侧的S M 5-S M 7站位水体性质较为接近。马六甲海峡呈现自东、西出口向海峡中部(S M 5站)盐度逐渐降低的趋势。
3.2观测站位的营养盐浓度
各站位营养盐浓度如图4所示。巽他陆架站位表层海水硝酸盐浓度平均值为(0.340±0.077)μmol/L;亚硝酸盐浓度平均值为(0.090±0.009)μmol/L;磷酸盐浓度平均值为(0.109±0.008)μmol/L;硅酸盐浓度平均值为(4.177±2.040)μmol/ L,尤以SS4站位硅酸盐含量最高,达到7.520μmol/ L。相比而言,马六甲海峡站位的营养盐浓度变化幅度较大,其硝酸盐浓度平均值为(1.185±0.672)μmol/L;亚硝酸盐浓度平均值为(0.312±0.230)μmol/L;磷酸盐浓度平均值为(0.084±0.071)μmol/ L;硅酸盐浓度平均值为(4.584±2.913)μmol/L,马六甲海峡的S M 1-S M 4号站硝酸盐浓度明显较高,但磷酸盐和硅酸盐浓度明显低于S M 5-S M7号站,而在S M 7站几乎所有营养盐浓度均为最高。
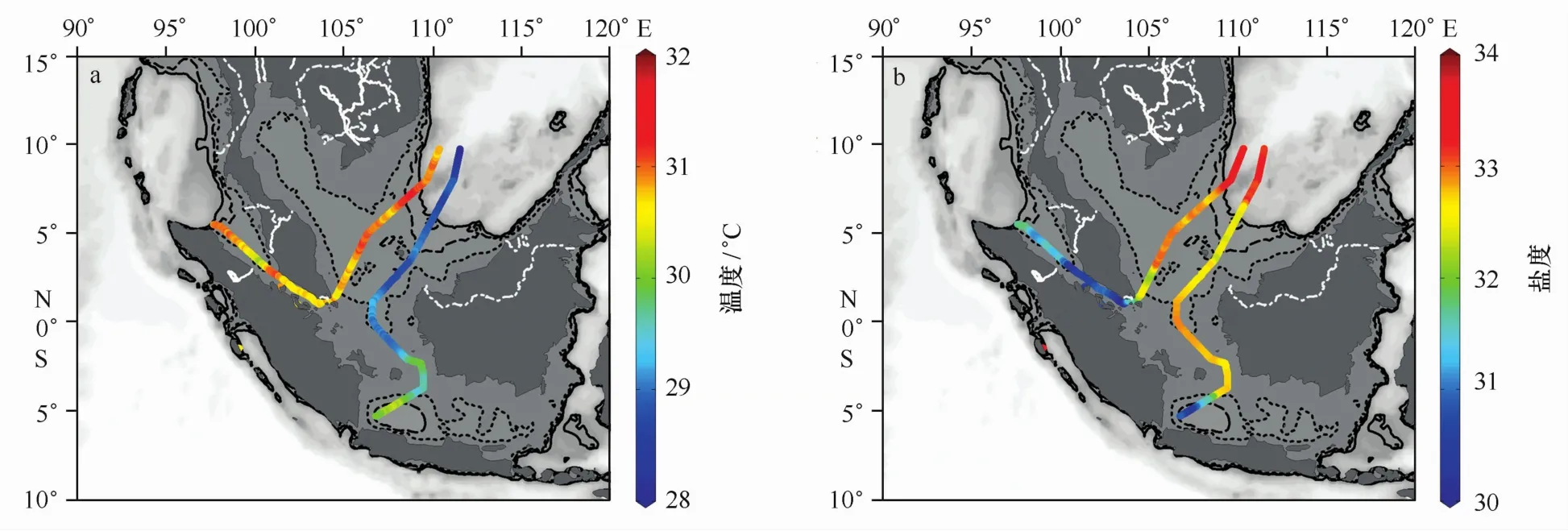
图2 巽他陆架和马六甲海峡的走航温度(a)和盐度(b)Eig.2 The underway sea surface temperature(a)and the sea surface sal inity(b)in the Sunda Shelf and the Strait of M alacca

图3 走航观测的T-S道具图及采样站位的密度分布Eig.3 TheT-Sdiagram for the underway surface watermass
通过各站位氮磷比和硅氮比可以看出,除马六甲海峡的S M4站氮磷比为17.6,接近Redfield比值外,S M 1-S M 3站氮磷比均远高于Redfield比值,而巽他陆架及S M 5-S M7站的氮磷比却明显低于16∶1;而对于硅氮比来说,除S M 1-S M 3站硅氮比接近1外,其余站位的硅氮比均远高于15∶16,其中尤以爪哇海的SS4站为最高,达到14.2。
3.3浮游植物粒级结构
各站位叶绿素a生物量及主要特征色素浓度如图5所示。可以看出,巽他陆架站位生物量明显低于马六甲海峡站位,巽他陆架站位叶绿素a生物量在0.014~0.127μg/L之间,平均值为(0.083±0.043)μg/L,其中爪哇海的SS4站位生物量最低,为0.014 μg/L;而马六甲海峡叶绿素a生物量在0.176~1.080μg/L之间,平均值为(0.433±0.315)μg/L,S M 5-S M 7站生物量明显较高,叶绿素a浓度均高于0.5μg/L。
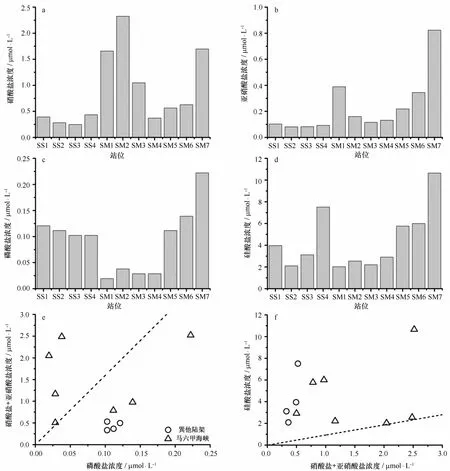
图4 各观测站位的硝酸盐(a)、亚硝酸盐(b)、磷酸盐(c)、硅酸盐(d)、氮磷比(e)和硅氮比(f)Eig.4 The nitrate(a),nitrite(b),phosphate(c),si l icate(d),N∶P ratio(e)and Si∶N ratio(f)of all stations
通过采用Uitz等的粒级计算方法[15],获得各站位Micro-(大于20μm)、Nano-(2~20μm)和Pico-(0.2~2μm)粒级的组成百分比如图6所示。在生物量较低的巽他陆架站位,除SS4站以外,其余各站Pico-级生物量均在80%以上;在水体温、盐性质相近的SS4和S M 1-S M 3站,粒级组成结构也较近似,其中Micro-级不到20%,Nano-级略高于20%,而Pico-级约在60%左右。S M 6和S M 7站具有较高的Micro-级百分比,约占40%~45%,同时Pico-级也占近50%,Nano-级所占百分比则较低,分别占6%和10%。而在生物量最高的S M 5站位,其Micro-和Nano-级则都较低,仅占约20%左右,Pico-级为最优势类群,占81%。

图5 各观测站位的叶绿素a(a)、多甲藻素(b)、19′-丁酰基氧化岩藻黄素(c)、岩藻黄素(d)、19′-己酰基氧化岩藻黄素(e)、别藻黄素(f)、玉米黄素(g)和叶绿素b(h)浓度Eig.5 The concentration of the Chla(a),peridinin(b),19′-butanoyloxyfucoxanthin(c),fucoxanthin(d),19′-hexanoyloxyfucoxanthin(e),alloxanthin(f),zeaxanthin(g)and Chlb(h)of all stations
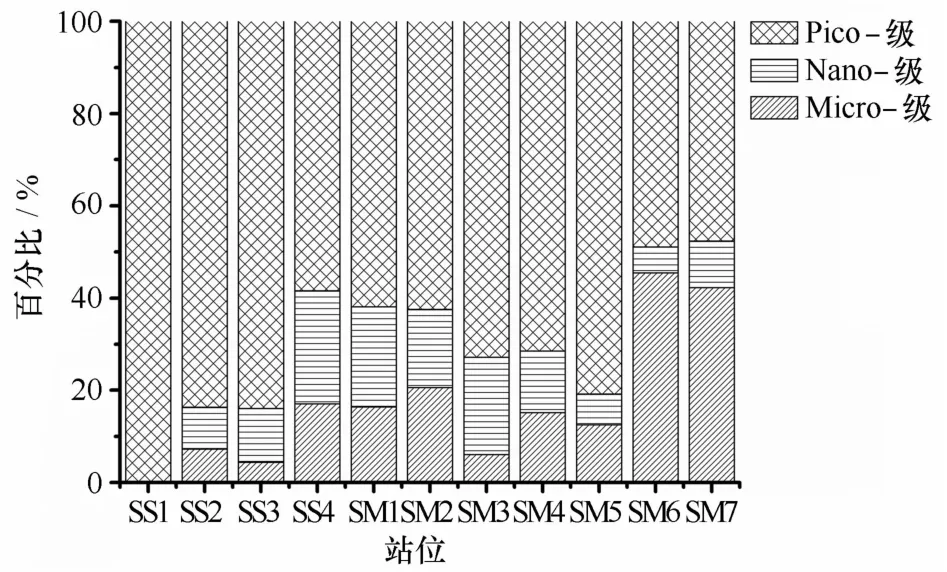
图6 各观测站位基于特征色素的浮游植物各粒级百分比组成Eig.6 The size-fractional percentage of the phytoplankton in all stations
3.4浮游植物的群落结构
通过C H E M TA X软件反演9类浮游植物群落结构百分比如图7所示。可以看出,聚球藻是研究区域的最主要优势类群,在巽他陆架,聚球藻占总生物量百分比在62%~81%之间,平均值为(71±7)%;而在马六甲海峡,聚球藻所占百分比在58%~77%之间,平均值为(67±7)%;另一类Pico-级浮游植物原绿球藻在巽他陆架的SS1-S M3站比例较高,占总生物量的13%~27%之间,其余站位较低。硅藻只在SS4、S M 6和S M7站具有较高百分比,在爪哇海的SS4站,硅藻百分比达到16%,而在S M6和S M 7站硅藻所占比例超过了20%。在S M1和S M 2站位,甲藻比例较低,仅占5%~7%左右,而其他站位并未检测到。类似地,在S M 2、S M4、S M6和S M7站位,青绿藻百分比也明显高于其他站位,均超过了10%。各站两类定鞭金藻比例均较低,仅在SS4站,定鞭金藻_6所占比例达到12%左右,而其他站位则多数以定鞭金藻_8为主,占6%左右。
4 讨论
4.1巽他陆架浮游植物群落结构及影响因子
柯志新等[6]报道春季巽他陆架叶绿素a浓度在0.05~0.58μg/L之间,其中0.7~3μm粒级的叶绿素a约占75%。而本文中巽他陆架站位叶绿素a生物量在0.014~0.127μg/L之间,除爪哇海的SS4站以外,其余各站Pico-级生物量均在80%以上。而针对生物量上的这一差异可以从营养盐浓度得以解释。柯志新等[6]研究中巽他陆架区域的硝酸盐浓度平均为0.72μmol/L,约为本研究中的2倍,同样的亚硝酸盐和磷酸盐也分别是本研究中的2.9倍和1.4倍,而本研究中硅酸盐含量却是其报道的1.5倍,由此推断本研究中这种低生物量主要是由营养盐浓度相对较低所致。较低的氮、磷营养盐会造成硅藻等较大型浮游植物生长受到限制,从而耐寡营养盐的Pico-级浮游植物成为优势[17]。同时,硅藻生长受到限制会影响到对硅酸盐的利用,从而导致硅酸盐浓度相对较高,产生超额硅酸盐剩余的现象[18]。
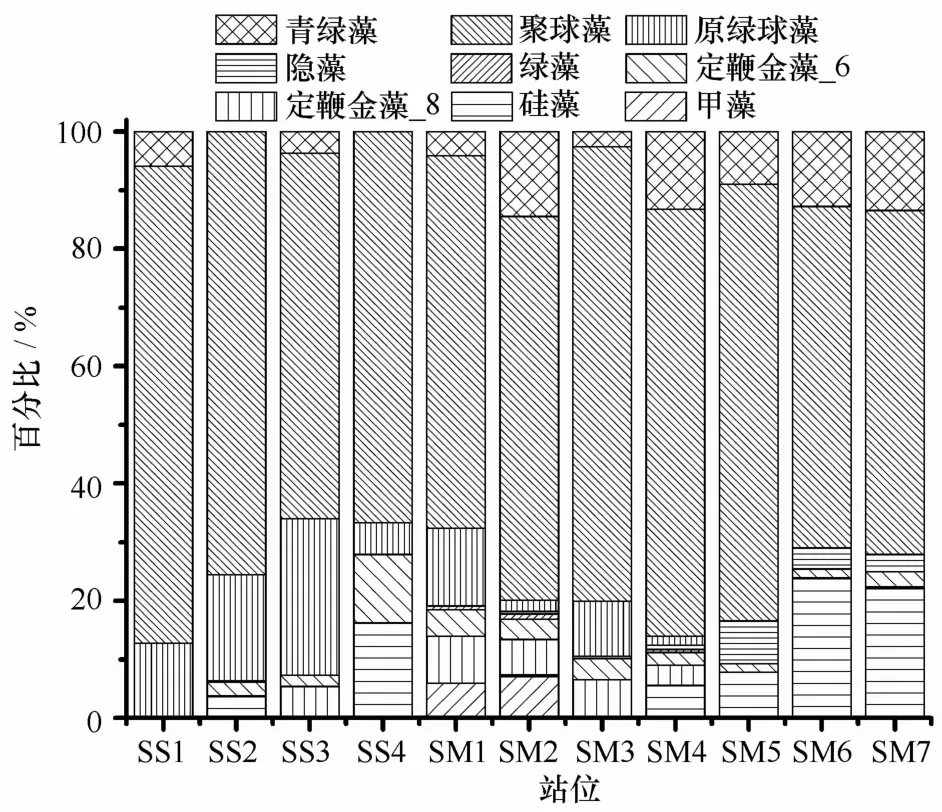
图7 各观测站位基于C H E M TA X反演的浮游植物群落百分比组成Eig.7 The group-specific contribution to the phytoplankton com munity in all stations
位于爪哇海的SS4站无论是在海水温、盐性质还是在浮游植物生物量上都表现出区别于巽他陆架其他站位的状态。相比于SS1-SS3站来说,SS4站具有更为明显的高温、低盐特征。在营养盐方面除硅酸盐较高外,其余营养盐浓度相差无几,SS4站硅酸盐浓度达到7.520μmol/L,加之较低的表层盐度,可以推断其受到陆源输入的影响较大。柯志新等[6]研究指出,爪哇海相对于南海南部具有更高的硝酸盐和硅酸盐浓度,同时叶绿素a浓度也明显较高,Pico-级浮游植物百分比接近76%。参考其研究中表层盐度平均为32.18,硅酸盐浓度仅为本研究中的一半左右,分析可能由于本次研究中SS4站受到爪哇岛或苏门答腊岛陆源径流影响更大,在降低盐度、提高营养盐浓度的同时,加大了海水浊度,使浮游植物可利用光强降低,从而导致如此低的生物量。另外,在群落结构上,SS4站表现出较高的硅藻和定鞭金藻_6的百分比,进一步证明了该站位可能存在由陆源径流引起的浮游植物群落演替。
4.2马六甲海峡浮游植物群落结构及影响因子
本研究中马六甲海峡站位体现出明显的盐度梯度特征,最低盐度出现在S M5站位,这主要是由于苏门答腊岛的Barumun河和马来半岛的Perak河河口分列S M5站南北所致,使该站表层盐度仅有30.11。Gin等[9]报道新加坡近岸的叶绿素a浓度周年平均值约为1.7μg/L,高生物量主要出现在西南季风期。而本研究中该区域叶绿素a浓度平均值为0.433 μg/L,最高值的S M5站为1.080μg/L,这可能是由于季风转换期水体层化所致。在其报道中,Pico-级及小型的Nano-粒级(小于8μm)的叶绿素a约占40%左右,而本研究中Pico-级浮游植物百分比变动在50%~80%左右,同时营养盐浓度也比其报道的平均值低,可见在季风转换期马六甲海峡同样存在由Pico-级取代较大粒级浮游植物的群落演替特征。值得注意的是,在盐度最低的S M5站,Pico-级浮游植物百分比超过80%,而聚球藻所占比例接近75%。S M 5站叶绿素a浓度分别为S M6和S M 7站的1.7倍和2.0倍,而营养盐浓度却还略低于S M6和S M7站,这说明S M5站浮游植物对陆源输入的营养盐存在着大量的消耗。同时可能由于氮限制(氮磷比约为7),硅藻并未形成较大生物量,致使硅酸盐并不能有效地被利用而随表层流流向S M6和S M 7站[3]。而在S M6和S M 7站处马六甲海峡宽度达到最低值,无论是来自陆源的还是因航速较慢引起的船舶污染都会更多,营养盐浓度更高,从而使硅藻达到较大的百分比。而自海峡中部向东南直至新加坡,浮游植物群落这种自聚球藻向硅藻的演替现象逐步加剧,据Gin等[10]研究结果表明,在新加坡近岸硅藻约占叶绿素a生物量的72%,而聚球藻约占18%。
通过对观测站位测定到的浮游植物各优势类群生物量与环境因子进行相关性分析(表2)可以看出,总叶绿素a浓度仅与盐度呈现出显著负相关性(p<0.050),说明各调查站位叶绿素a生物量受到多重因素共同作用的影响。叶绿素a生物量和盐度呈现负相关主要是由于陆源输入增加了研究区域的营养盐浓度,使硅藻等适高营养的类群提高生物量,从而总体生物量有所提高;硅藻生物量与磷酸盐(p<0.050)和硅酸盐(p<0.010)呈现显著正相关性;而研究海区主要以Pico-粒级浮游植物如聚球藻为优势类群,其仅与亚硝酸盐呈显著负相关(p<0.050)。众所周知表层海水聚球藻为优势属于热带海区的普遍共性,主要由其喜好高温、高光强和寡营养盐的特性所致[19],而陆源输入引起的盐度的降低会导致聚球藻生长受到影响,硅藻等类群更易生长,虽然浮游植物总生物量增加,但聚球藻优势度下降,本研究结果也符合这一现象。而定鞭金藻_8与磷酸盐的显著负相关性(p<0.010)及与氮磷比的正相关性(p<0.010)体现了其与硅藻等占据不同的生态位,虽在研究海区表层并未形成优势,但符合其在叶绿素最大层分布的特征[20]。

表2 浮游植物主要类群及生物量与主要环境因子相关性Tab.2 The correlation analysis between the TChlabiomass of the major phytoplankton groups and the environmental parameters
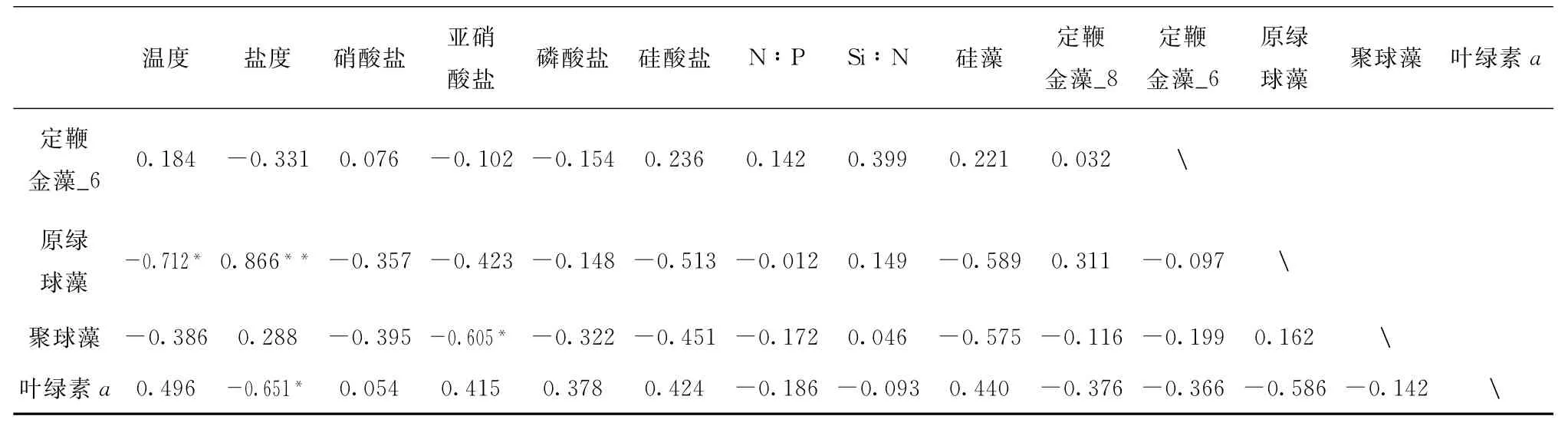
续表2
5 结论
春季季风转换期巽他陆架和马六甲海峡表层浮游植物生物量和群落结构分布表现出明显的空间差异,高叶绿素a生物量主要集中在马六甲海峡东南海区,而巽他陆架生物量略低,最低值出现在爪哇海的SS4站。在巽他陆架区,叶绿素a生物量较低,类群上主要以Pico-级浮游植物为优势,聚球藻生物量较高,也有部分原绿球藻;而在马六甲海峡西北段浮游植物生物量略高于巽他陆架站位,群落结构仍然主要以聚球藻为优势,但体现出了相对较高的类群组成上的多样性。马六甲海峡东南段受到陆源淡水输入的影响,具有较高的浮游植物生物量,群落结构上也存在着自聚球藻向硅藻的演替特征,但在生物量最高的S M 5站位仍然以聚球藻为绝对优势类群。总体来看,巽他陆架和马六甲海峡在春季季风转换期生物量相对较低,陆源营养盐的输入会导致受影响区域生物量的升高。不同类群浮游植物的演替主要由各自区域营养盐浓度和氮磷比所致。由于陆源输入的影响,接近河口海区耐寡营养盐的聚球藻在总浮游植物生物量贡献上的优势度会降低。
致谢:感谢国家自然科学基金委员会印度洋科学考察航次项目的资助;感谢中国科学院南海海洋研究所“实验一号”船队全体工作人员的协作;感谢厦门大学环境与生态学院黄邦钦教授、国家海洋环境监测中心翟惟东研究员在样品测定方面的协助和支持。
[1]Tomascik T,Mah A J,Nontj i A,et al.The Ecology of the Indonesian Seas PartⅠ[M].H ong Kong:Periplus Editions Ltd,1996.
[2]Chen C T A,Wang S L,Wang B J,et al.Nutrient budgets for the South China Sea basin[J].M arine Chemistry,2001,75(4):281-300.
[3]W yrtki K.Physical oceanography of the southeast Asian waters.Scientific results of marine investigations of the South China Sea and the Gulf of Thai land[R].La Jol la,C A:Scripps Institution of Oceanography,1961:195.
[4]Bray N A,Hautala S,Chong J,et al.Large-scale sealevel,thermocl ine,and wind variationsin the Indonesian throughflow region[J].Journal of Geophysical Research,1996,101(C5):12239-12254.
[5]Martinez J I,Taylor L,Deckker P D,et al.Planktonic foraminifera from the eastern Indian Ocean:distribution and ecology in relation to the W estern Pacific W arm Pool(W P W P)[J].M arine Micropaleontology,1998,34(3/4):121-151.
[6]柯志新,黄良民,李刚,等.春末夏初巽他陆架表层水体的叶绿素粒级结构及其和营养盐的关系[J].海洋学报,2012,34(3):190-196.
Ke Zhixin,H uang Liangmin,Li Gang,et al.Size structure of chlorophyl l ain relation to environ mentalfactorsin surface waters of Sunda Shelf during late spring and early su m mer[J].Haiyang Xuebao,2012,34(3):190-196.
[7]Ke Z X,Tan Y H,H uang L M,et al.Relationship between phytoplankton composition and environ mentalfactorsin the surface waters of southern South China Sea in early su m mer of 2009[J].Acta Oceanologica Sinica,2012,31(3):109-119.
[8]Ooi S H,Samah A A,Braesicke P.Primary productivity and its variabi l ity in the equatorial South China Sea during the northeast monsoon[J].Atmospheric Chemistry and Physics Discussions,2013,13(8):21573-21608.
[9]Gin K Y H,Lin X,Zhang S.Dynamics and size structure of phytoplankton in the coastal waters of Singapore[J].Journal of Plankton Research,2000,22(8):1465-1484.
[10]Gin K Y-H,Zhang S,Lee Y K.Phytoplankton com munity structurein Singapore's coastal waters using H PLC pigment analysis and flow cytometry[J].Journal of Plankton Research,2003,25(12):1507-1519.
[11]Natarajan R,Ross S A,Chua T E.Analysis of the state ofthe marine environment ofthe Straits of Malacca and Singapore[J].Singapore Journalof International&Comparative Law,1998,2:323-349.
[12]Strickland J D H,Parsons T R.A practical handbook of seawater analysis[R].Ottawa:Eisheries Research Board of Canada,1968.
[13]Roy S,Llewel lyn C A,Egeland E S,et al.Phytoplankton Pigments:Characterization,Chemotaxonomy and Appl ications in Oceanography[M]. New York:Cambridge University Press,2011.
[14]王磊,钟超,柳欣,等.夏季南海东北部和东海陆架浮游植物群落结构昼夜变化的比较研究[J].海洋学报,2013,35(6):170-177.
Wang Lei,Zhong Chao,Liu Xin,et al.The comparative study on the diurnal variations of phytoplankton com munity between the northeastern South China Sea and the East China Sea in su m mer[J].Haiyang Xuebao,2013,35(6):170-177.
[15]Uitz J,Claustre H,M orel A,et al.Vertical distribution of phytoplankton com munitiesin open ocean:An assessment based on surface chlorophyl l[J].Journal of Geophysical Research,2006,111:C08005.
[16]M ackey M D,M ackey D J,Higgins HW,et al.C H E M TA X-a program for estimating class abundances from chemical markers:appl ication to H PLC measurements of phytoplankton[J].M arine Ecology Progress Series,1996,144:265-283.
[17]Turpin D H,Harrison P J.Limiting nutrient patchiness and its rolein phytoplankton ecology[J].Journal of Experimental Marine Biology and E-cology,1979,39(2):151-166.
[18]Tréguer P J,Rocha C L D L.The world ocean si l ica cycle[J].Annual Review of Marine Science,2013,5(1):477-501.
[19]Chen B Z,Liu H B,Landry M R,et al.Close coupl ing between phytoplankton growth and microzooplankton grazing in the western South China Sea[J].Limnology and Oceanography,2009,54(4):1084-1097.
[20]Schoemann V,Becquevort S,Stefels J,et al.Phaeocystisbloomsin the global ocean and their control l ing mechanisms:a review[J].Journal of Sea Research,2005,53(1/2):43-66.
The distribution of phytoplankton com munity structure in the Sunda Shelf and the Strait of M alacca during spring intermonsoon
Wang Lei1,2,Leng Xiaoyun1,Sun Qingyang2,Wang Dongxiao2,Zhang Cuixia1,Sun Jun1
(1.College of M arine Science&Engineering,Tianjin University of Science&Technology,Tianjin300457,China;2.State Key Laboratory of Tropical Oceanography,South China Sea Institute of Oceanology Chinese Academy of Sciences,Guangzhou510301,China)
Surface water samples were collected in order to study the spatial distribution of phytoplankton com munity in the Sunda Shelf and the Strait of M alacca during M arch and M ay 2013.Significant spatial variations of phytoplankton total Chla(T Chla)biomass and com munity structure were observed using the photosynthetic pigments method.The T Chlabiomass was low to some extent in the Sunda Shelf during the intermonsoon season,when and where the minimum value was only 0.014μg/L at Sta.SS4 in the Java Sea,and the average was(0.083± 0.043)μg/L in the Sunda Shelf.The picophytoplankton dominated in the Sunda Shelf and contributed more than 80%of the total biomass.The sal inity gradient was distinct along the Strait of M alacca from northwest to the southeast.And the lowest sal inity appeared at Sta.S M 5 with the highest T Chlaconcentration(1.080μg/L),which was outstanding from the average value of allthe strait stations[(0.433±0.315)μg/L].TheSynechococcus dominated 60%-80%ofthe total phytoplankton biomass atthe stationsin the Sunda Shelf and S M 1-S M4.Even though the T Chlaconcentrations was less than it at Sta.S M 5,they were sti ll apparently higher at Sta.S M 6 and S M7 than the biomass at the other stations.And the Diatoms contributed more than 20%at these two stations as a result of the higher nutrient concentration carrying by the river runoff,especially for phosphate and si l icate.On the contrary,the superiority ofSynechococcusmight be weakened by the intrusion of low-sal inity,high-nutrients water.
phytoplankton;com munity structure;Sunda Shelf;Strait of M alacca
S963.21+3
A
0253-4193(2015)02-0120-10
2014-04-11;
2014-06-18。
国家自然科学基金面上项目“印度洋浮游植物功能群初级生产调控因子与碳输出效率研究”(41270491);热带海洋环境国家重点实验室开放课题“南海浮游植物群落结构在涡旋影响下的亚中尺度响应”(L T O1307);国家自然科学基金青年基金“东印度洋营养盐上行效应调控下的浮游植物群落水平初级生产力研究”(41406158);“海洋酸化对东印度洋微型浮游动物群落及摄食影响研究”(41306119);天津市高等学校创新团队培养计划(T D12-5003)。
王磊(1981—),男,辽宁省阜新市人,博士,主要从事海洋浮游植物生理、生态学研究。E-mai l:lwangbor@gmai l.com
孙军,教授。E-mai l:phytoplankton@163.com
