春季东海有害藻华高发区仔稚鱼时空分布
2015-10-22陈渊戈钟俊生徐兆礼陈佳杰
陈渊戈,钟俊生,徐兆礼*,陈佳杰
(1.中国水产科学研究院东海水产研究所农业部海洋与河口渔业重点开放实验室,上海200090;2.上海海洋大学,上海201306)
春季东海有害藻华高发区仔稚鱼时空分布
陈渊戈1,钟俊生2,徐兆礼1*,陈佳杰1
(1.中国水产科学研究院东海水产研究所农业部海洋与河口渔业重点开放实验室,上海200090;2.上海海洋大学,上海201306)
根据2011年春季(3月29日—5月27日)海洋综合调查,对东海有害藻华高发水域(28°15′36″~30°30′16″N、121°52′36″~123°15′6″E)的仔稚鱼种类组成、时空分布以及主要优势种与环境因子的相关性进行了探讨。调查共采集到仔稚鱼27种,前几位优势种分别为日本鳀Engraulisjaponicus、鰕虎科sp.(Gobi idae sp.)、六丝钝尾鰕虎A mblychaeturichthyshexanema、小黄鱼Larimichthys polyactis,该4种占据了总丰度的91.23%。优势种丰度和发育阶段组成的时空变化显示,日本鳀在调查期间有两次产卵小高峰,其仔鱼孵化后向岸漂流;小黄鱼和六丝钝尾鰕虎在此期间只有一次产卵高峰,其仔鱼在舟山群岛邻近水域进行保育。可以看出,调查水域及邻近水域是部分鱼类产卵,仔稚鱼保育、向岸漂流的场所或途径,是鱼类早期生活史中重要生境,一旦暴发有害藻华,补充群体将受到不良影响。多元分析结果显示,所选环境因子对几位优势种仔稚鱼的分布影响不大,推测产卵场位置和保育方式可能是更重要的原因。
仔稚鱼;时空分布;东海;有害藻华
陈渊戈,钟俊生,徐兆礼,等.春季东海有害藻华高发区仔稚鱼时空分布[J].海洋学报,2015,37(2):44—54,doi:10.3969/j.issn. 0253-4193.2015.02.005
Chen Yuange,Zhong Junsheng,Xu Zhaol i,et al.Spatio-temporal distribution of larval and juveni le fishes at frequent harmful algal blooms(H A Bs)areas of the East China Sea in spring[J].Haiyang Xuebao,2015,37(2):44—54,doi:10.3969/j.issn.0253-4193.2015.02.005
1 引言
有研究运用多年的遥感数据[1]支持了Hjort的临界期假说[2]和Cushing的match/mismatch理论[3],认为在鱼类产卵季节藻华的发生能提高仔鱼成活率。但鱼类在早期生活史中较为脆弱,更多学者关注有害藻华对鱼类早期生活史的潜在威胁。W arlen发现短裸甲藻Gy mnodinium breve在河口引起的有害藻华对不同鱼类的补充量分别造成了持续性(白点牙鲆Paralichthysalbigutta,斑鳍副棘鲆Citharichthysspilopterus)、暂时性(暴油鲱Erevoortiatyrannus,漠斑牙鲆Paralichthyslethostig ma,黄尾平口石首鱼Leiostomu sxanthurus,鲻M ugilcephalus,斑纹油蛇鳗M yrophis punctatus,等)和不明显(波纹绒须石首鱼Micropogonia sundulatus,菱体兔牙鲷Lagodon rhomboids)影响,持续性影响表现为补充量即使在有害藻华结束后仍然维持在较低水平,暂时性影响则在有害藻华结束后出现高补充量[4]。有害藻华对鱼卵、仔稚鱼的危害大致有造成缺氧、物理损伤和藻毒素毒性损伤3种[5]。在这些威胁中,藻毒素可以通过食物链富集,对人类健康造成威胁,受到最为广泛的关注,其中又以PSP威胁最大。急性试验证明了PSP对仔鱼的致死作用[6—8],在亚急性试验中斑马鱼Canio rerio仔鱼出现了麻痹、反应减弱、身体水肿和生长速度减慢等症状[9]。
近年来东海有害藻华频发且呈上升趋势,每年5、6月于浙江省沿岸水域频发,尤其是舟山群岛水域更为严重[10]。研究表明,东海大规模赤潮可能通过改变微型浮游动物的群落结构,进而影响大型浮游动物的种群数量和群落结构[11]。而大中型浮游动物是仔稚鱼主要的饵料生物,仔稚鱼在初次开口和变态着底(食性转换)的两个临界期如果不能获取足够的适口饵料,会因为饥饿导致死亡率进一步升高[2],这将对鱼类群体补充造成负面影响,进而可能在局部地区对渔业资源量造成损害。有害藻华对东海仔稚鱼群体的威胁评估尚未见报道,相关基础资料也较为缺乏。
学者们对长江口、杭州湾及浙江近海水域鱼类浮游生物的组成和分布特征进行了调查研究[11—14],分析了环境因子对鱼类浮游生物组成和分布的影响[15—16]。这些调查根据各自的研究目的按季节或者逐月进行采样,如果要评估有害藻华频发月份内仔稚鱼受到的影响,在更小时间尺度上仔稚鱼群体的时空变化的数据支持略显不足。本研究根据2011年春季对东海有害藻华高发海域的密集走航调查,分析在有害藻华易发的月份里仔稚鱼种类组成和分布的时空特征,以期为评估东海有害藻华对鱼类早期生活史的影响提供基础依据。
2 材料方法
2.1调查站位及时间
M C2011航次在浙江省近海水域设置了5个断面(R A、RB、Z A、ZB、ZC),每个断面预设了8个站位,调查范围为28°15′36″~30°30′16″N、121°52′36″~123°15′6″E(图1)。从2011年3月29日至2011年5月27日,“润江1”海洋调查船共进行了6航次(A PR1、A PR2、A PR3、M A Y1、M A Y2、M A Y3)调查,RB、Z A、ZB断面开展了6次调查,其余两个断面开展了4次调查(表1)。
2.2采样方法和样品测定
在每一站位使用大型浮游生物网由底至表拖曳采集仔稚鱼,网口直径80 cm,网长280 cm,网衣孔径0.505 m m,所得样品保存于5%海水福尔马林溶液。
所得样品带回陆上实验室鉴定到尽可能低的分类阶元,根据Kendall等[17]划分仔稚鱼发育阶段,测量全长或标准体长。受目微尺量程限制,不足14 m m的个体在体视镜(Zeiss Stemi2000)下通过目微尺测量,超过14 m m个体使用数显游标卡尺(Mitutoyo CD-6"CSX)测量。科名按Nelson[18],中文名和学名按伍汉霖等[19],同属种名按英文字母顺序排列。
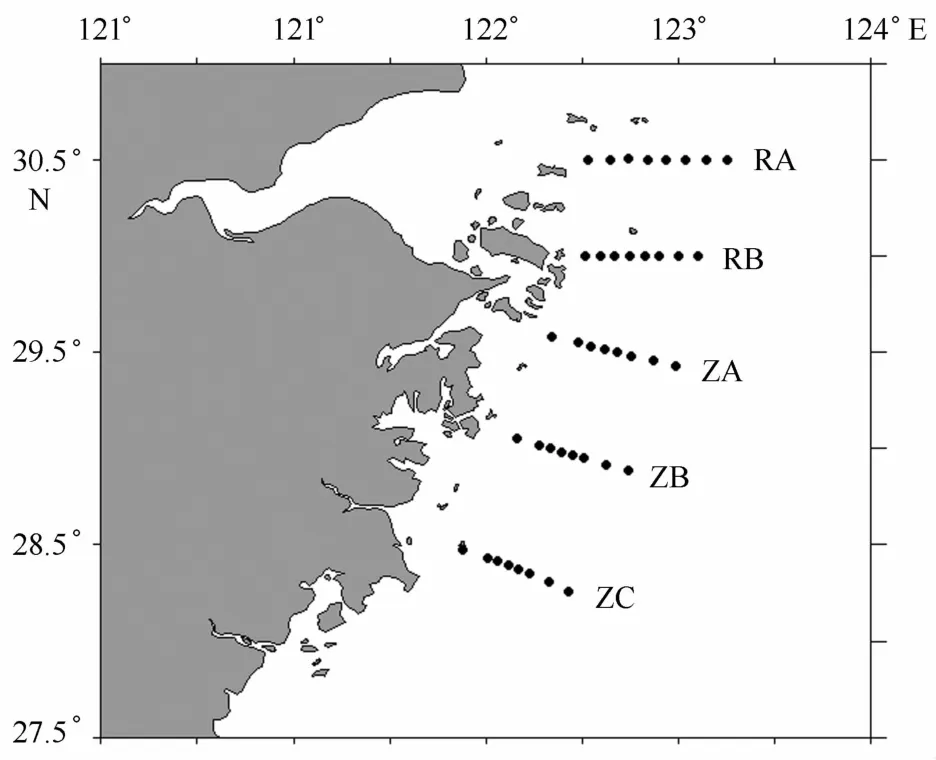
图1 调查站位Eig.1 Locations of sampl ing sites

表1 各航次调查时间及调查断面Tab.1 Periods and sampling sites of each cruise
2.3环境数据
环境数据使用航次共享的水文数据,包括水深、水温、盐度、溶解氧、p H、叶绿素a、透明度。由于仔稚鱼早期多营浮游生活,连续测量的数据(如水温、盐度等)选择5 m水深读数进行分析。从大型浮游生物网中挑出浮游动物,晾干后进行称量得到生物量,作为近似的饵料生物量参数加入分析。
2.4数据处理
在Surfer8.0中绘制仔稚鱼的丰度分布图,使用多元线性回归方法分析环境因子和仔稚鱼丰度的相关性。对仔稚鱼站位-种类丰度数据进行对数变换,计算各站位间的Bray-Curtis距离,进行层次聚类和非度量多维尺度分析。
3 结果
3.1种类组成
共采集到仔稚鱼1 239尾(包括幼鱼1尾),除1种未定种外,其余样品经鉴定隶属于18科27种。石首鱼科和舌鳎科的物种丰富度相对较高,各有4种(表2)。
日本鳀Engraulis japonicus丰度最高,共计832尾,占所有航次总丰度的67.15%。其次是鰕虎科sp.(Gobi idae sp.),共154尾,占总丰度的12.43%。丰度前几位的种类中,小黄鱼Larimichthys polyactis经济价值较高,共采到55尾,占总丰度的4.44%。总样本数不足10尾的偶见种有20种,隶属于15科(表2)。
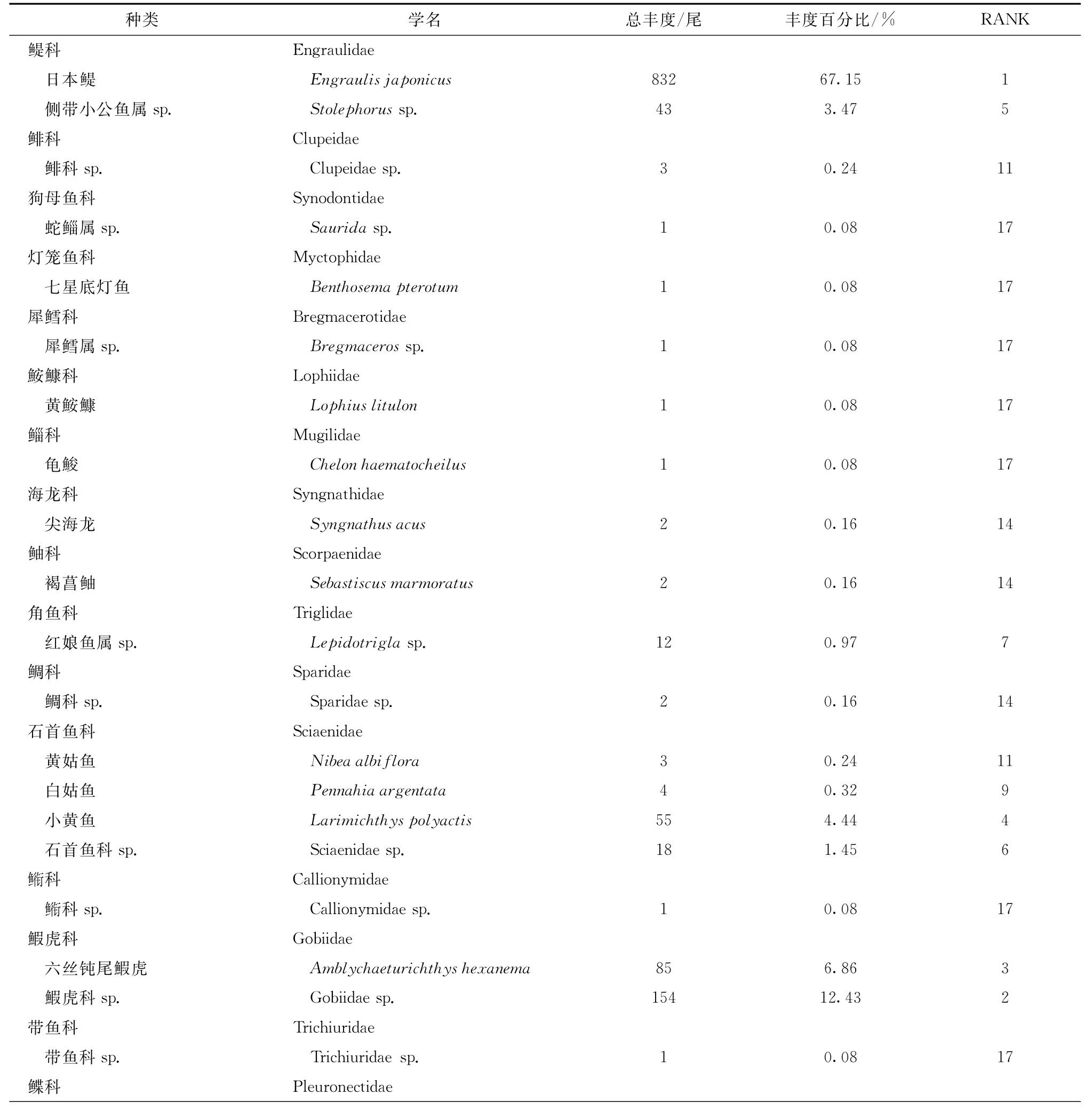
表2 仔稚鱼种类组成及丰度Tab.2 Species list and abundance of larval and juvenile fishes
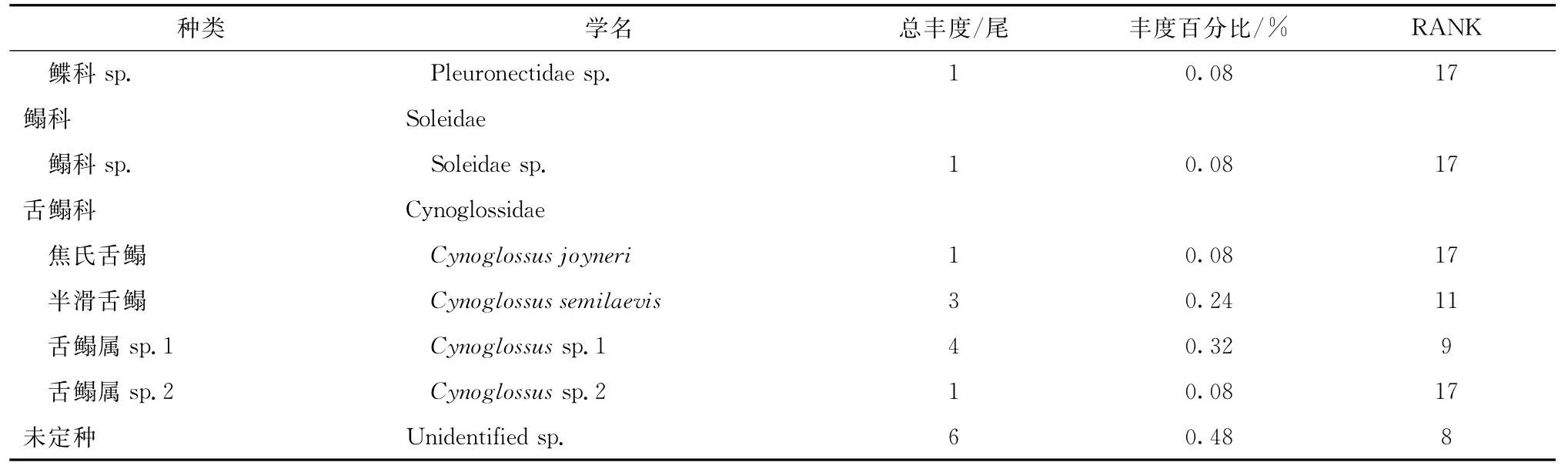
续表2
3.2优势种仔稚鱼的时空分布特征
调查期间日本鳀的前弯曲期、弯曲期、后弯曲期仔鱼各占53.41%、44.73%、1.86%。3月底至4月上半月(APR1~APR2),日本鳀仔鱼大量分布在离岸最远的外侧站位,发育阶段组成以前弯曲期仔鱼为主;从4月下半月至5月初(APR3~M A Y1),日本鳀较多地在相对近岸的站点出现,并且发展成为以弯曲期仔鱼为主;随后(M A Y2~M A Y3)观察到日本鳀重新在远岸外侧站位大量出现,前弯曲期仔鱼在数量上重新占据了主导地位(图2)。体长分布变化与此发展趋势相吻合,从3月底至5月初,日本鳀仔鱼体长呈增长态势,随后又回到与4月初相近的体长分布。
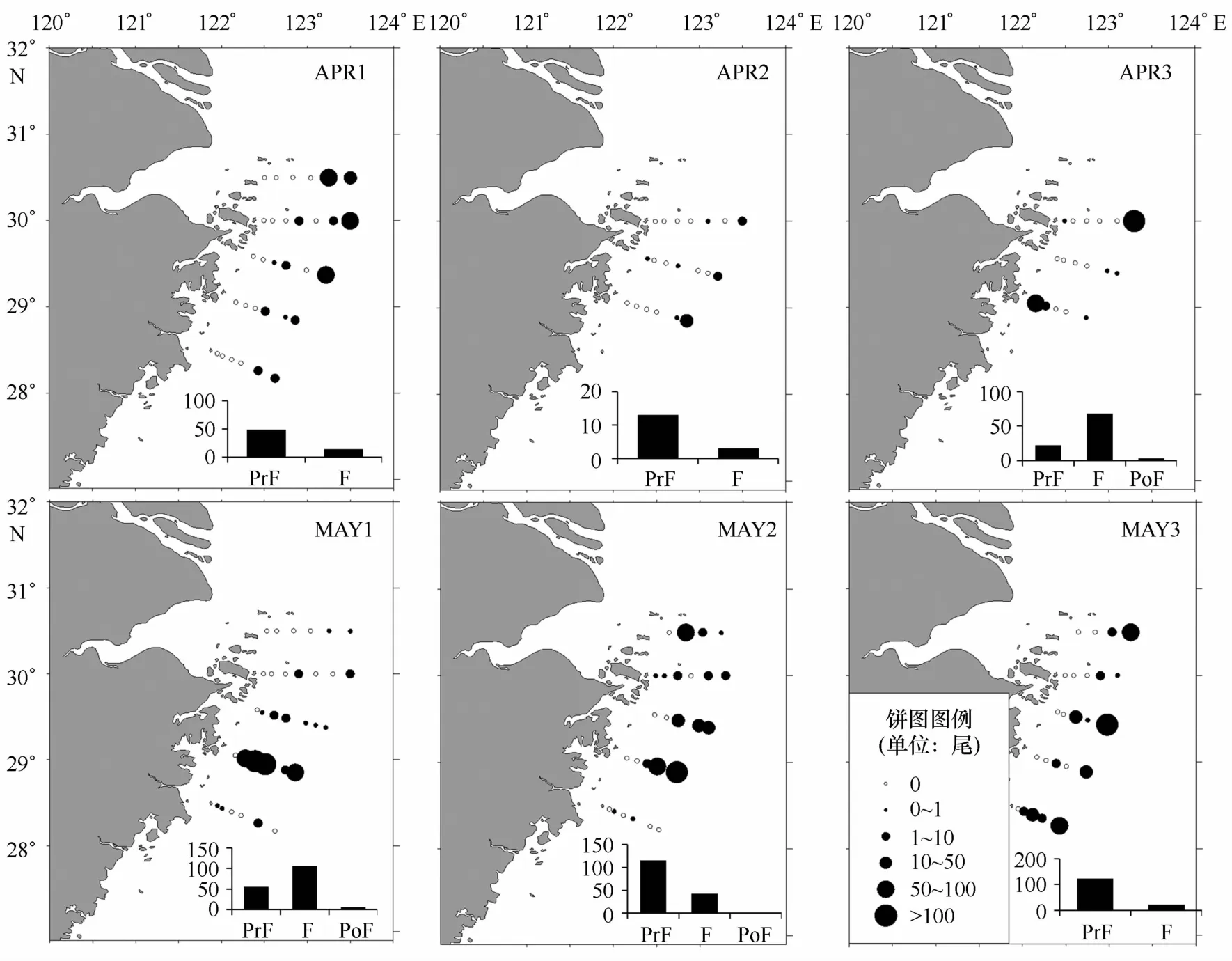
图2 日本鳀仔稚鱼丰度分布和发育阶段组成Eig.2 Developmental stage composition and abundance distribution ofE.japonicus
所获小黄鱼仔稚鱼以早期仔鱼为主,卵黄囊期仔鱼和前弯曲期仔鱼占总数的80.00%,而且基本上都出现在北面的R A、RB、Z A断面,外侧站位丰度略高于内侧站位。总体上看,空间分布无明显的移动趋势。在时间序列上,总丰度逐渐减低,后期仔鱼(弯曲期和后弯曲期)比例在4月底至5月中增高(图3)。到5月下半所获小黄鱼仔鱼丰度骤减,最后两个航次仅采集到的2尾,前4航次的小黄鱼仔鱼体长分布显现出明显的增长。
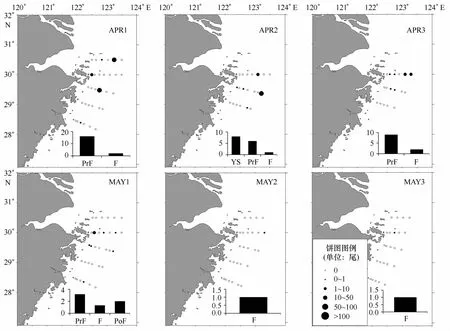
图3 小黄鱼丰度分布和发育阶段组成Eig.3 Developmental stage composition and abundance distribution ofL.polyactis
鰕虎科sp.总体以前弯曲期仔鱼为主,占总数的85.43%。鰕虎科sp.较为均匀的分布在整个调查水域范围内,每航次的丰度在整体上呈减少的趋势,前弯曲期仔鱼在所有航次中占比均最高(见图4)。整体来看鰕虎科sp.体长变化相对不明显,只在A PR3和M A Y1采集到少量后期仔鱼,体长略大。
与前3种相比,六丝钝尾鰕虎的后弯曲期仔鱼占比最高,达30.49%,总体看来北面断面丰度高于南面断面。后弯曲期仔鱼占比在4月上半还不及前弯曲期仔鱼,从4月下半开始占据优势地位(见图5),体长也有较大幅的增长(见图6)。5月下半丰度下降,到5月底(M A Y3)航次已经没能采到六丝钝尾鰕虎。
综合这4种鱼类仔稚鱼的丰度分布和生长发育趋势,可以略微探讨它们早期生活史的异同。在调查期间日本鳀体长分布呈现先增再减的格局,并且早期仔鱼前后两次在外侧站位大量出现,意味着在3月下半和5月下半调查水域外侧可能有两次产卵小高峰,孵化后向岸漂流的特征比较明显。同理,小黄鱼和六丝钝尾鰕虎在此期间可能有一次产卵高峰,鰕虎科sp.较为平稳地产卵繁殖。小黄鱼在舟山近海外侧站位出现较多,六丝钝尾鰕虎较为随机地出现在舟山渔场附近,都显示出了明显的生长发育趋势,表明了这一水域对它们二者的保育功能。鰕虎科sp.均匀地分布在整个调查水域,其早期生活史可能更为复杂,尚待进一步发现。
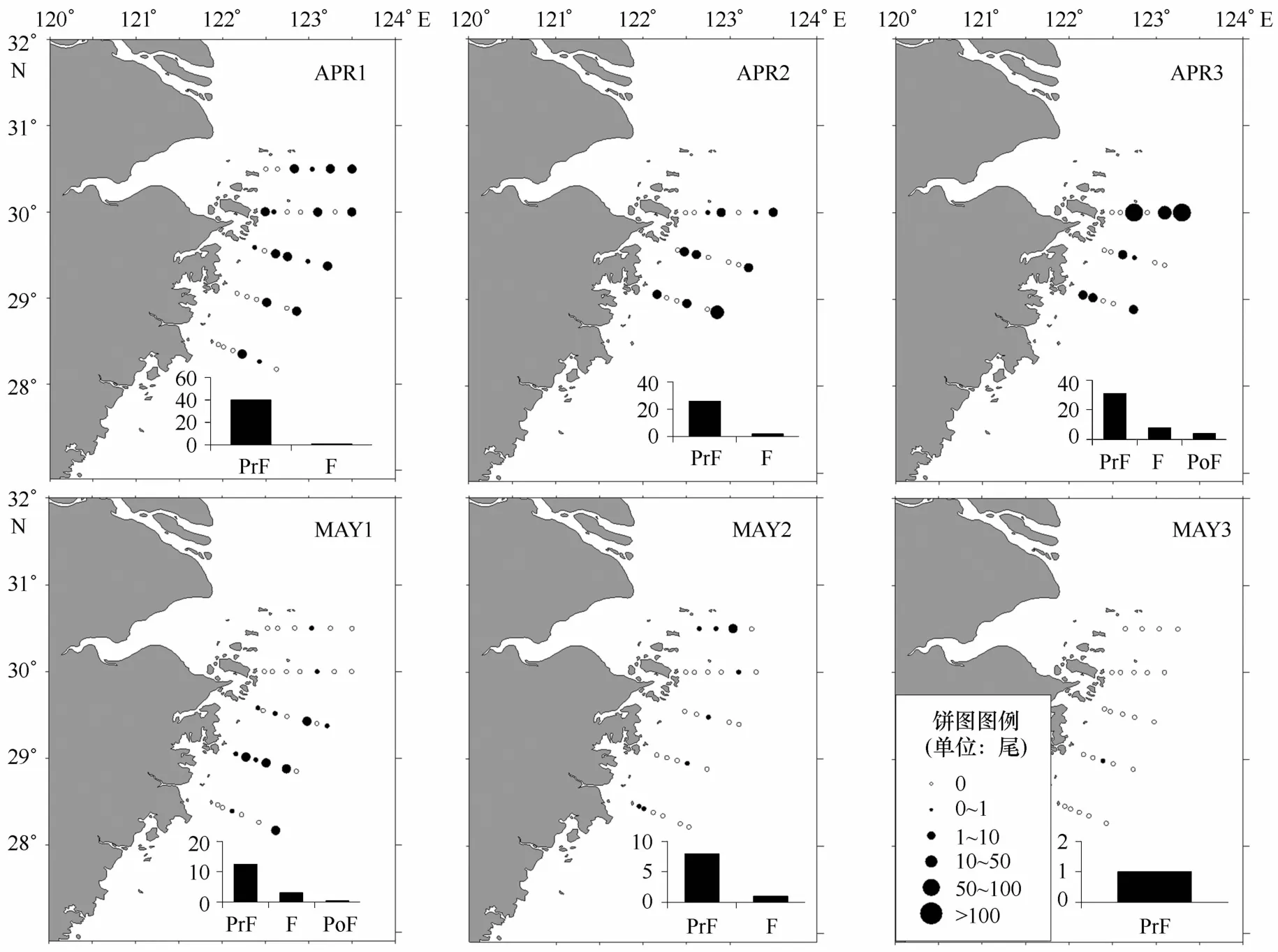
图4 鰕虎科sp.丰度分布和发育阶段组成Eig.4 Developmental stage composition and abundance distribution of Gobi idae sp.
3.3多元分析
根据每个站位的仔稚鱼丰度进行层级聚类分析,4月的3个航次(A PR1~A PR3)和5月的3个航次(M A Y1~M A Y3)各被聚为一支(见图7)。A N OSIM显示这两支在0.1的置信水平上通过显著性检验(R =0.78,P=0.09)。
日本鳀、鰕虎科sp.和六丝钝尾鰕虎贡献的相异性累积百分比超过70%,前五位的相异性累积百分比超过90%(表3)。日本鳀4月平均丰度不到5月平均丰度的一半,鰕虎科sp.、六丝钝尾鰕虎和小黄鱼在4月的平均丰度均远高于5月,可以看出它们的贡献主要是来自丰度的差异。除侧带小公鱼属sp.之外,还有16种仔稚鱼仅在4月或5月出现,但由于丰度极低,对总相异性贡献很小。
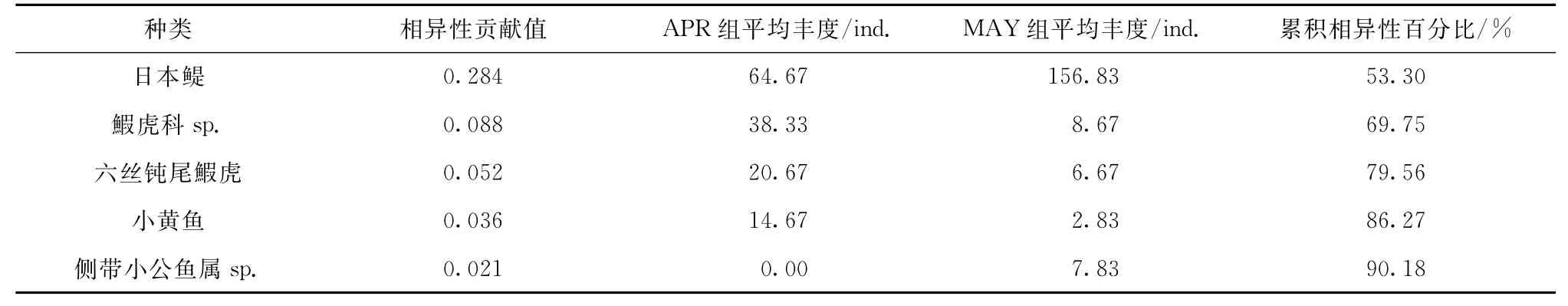
表3 4月航次和5月航次间相异性贡献度分析Tab.3 Results of SIM PER analyses between group APR and group M A Y
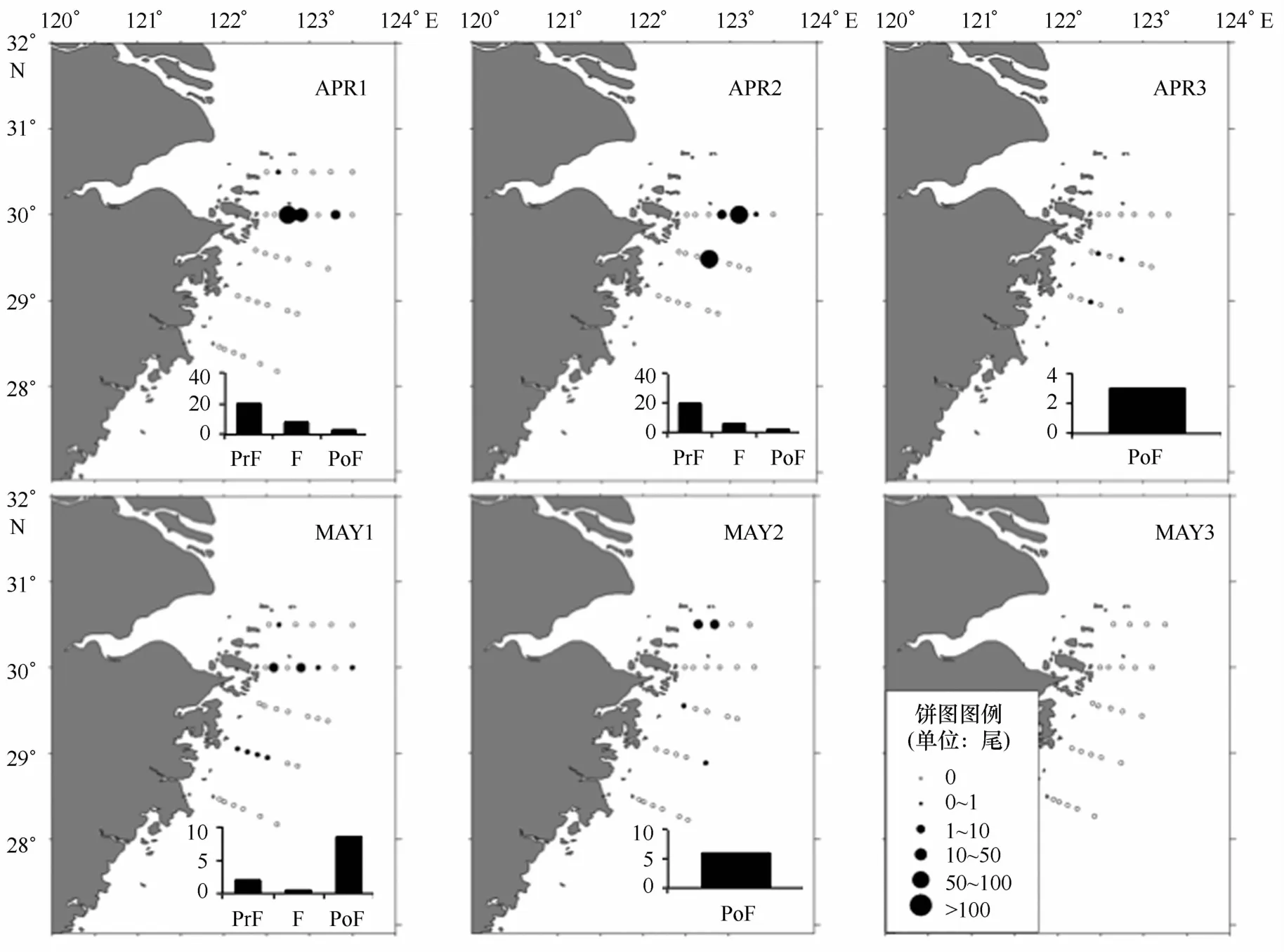
图5 六丝钝尾鰕虎丰度分布和发育阶段组成Eig.5 Developmental stage composition and abundance distribution ofA.hexanema
对每一航次的站位进行聚类和排序分析发现,各航次站位的聚类在空间位置上不具有良好的解释性(图略),站位间差异大,分组间的差异多不能通过显著性检验(α=0.05)。
逐步多元回归分析结果显示,在本研究的时空尺度下,选取的7个环境因子对六丝钝尾鰕虎、鰕虎科sp.的分布变化的解释量很小(调整R方低)。日本鳀的丰度分布与透明度极显著负相关,与水深显著正相关;小黄鱼的丰度分布与盐度极显著正相关,与水温和透明度显著负相关(表4)。
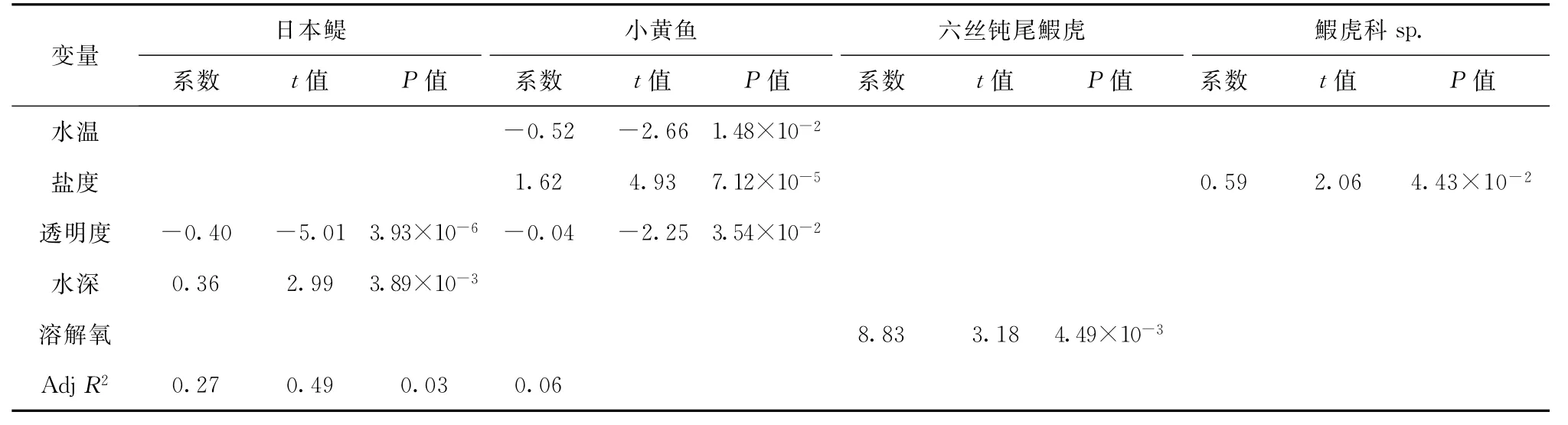
表4 主要优势种与环境因子回归分析结果Tab.4 Results of regression analyses between dominantlarval fishes and environmental factors
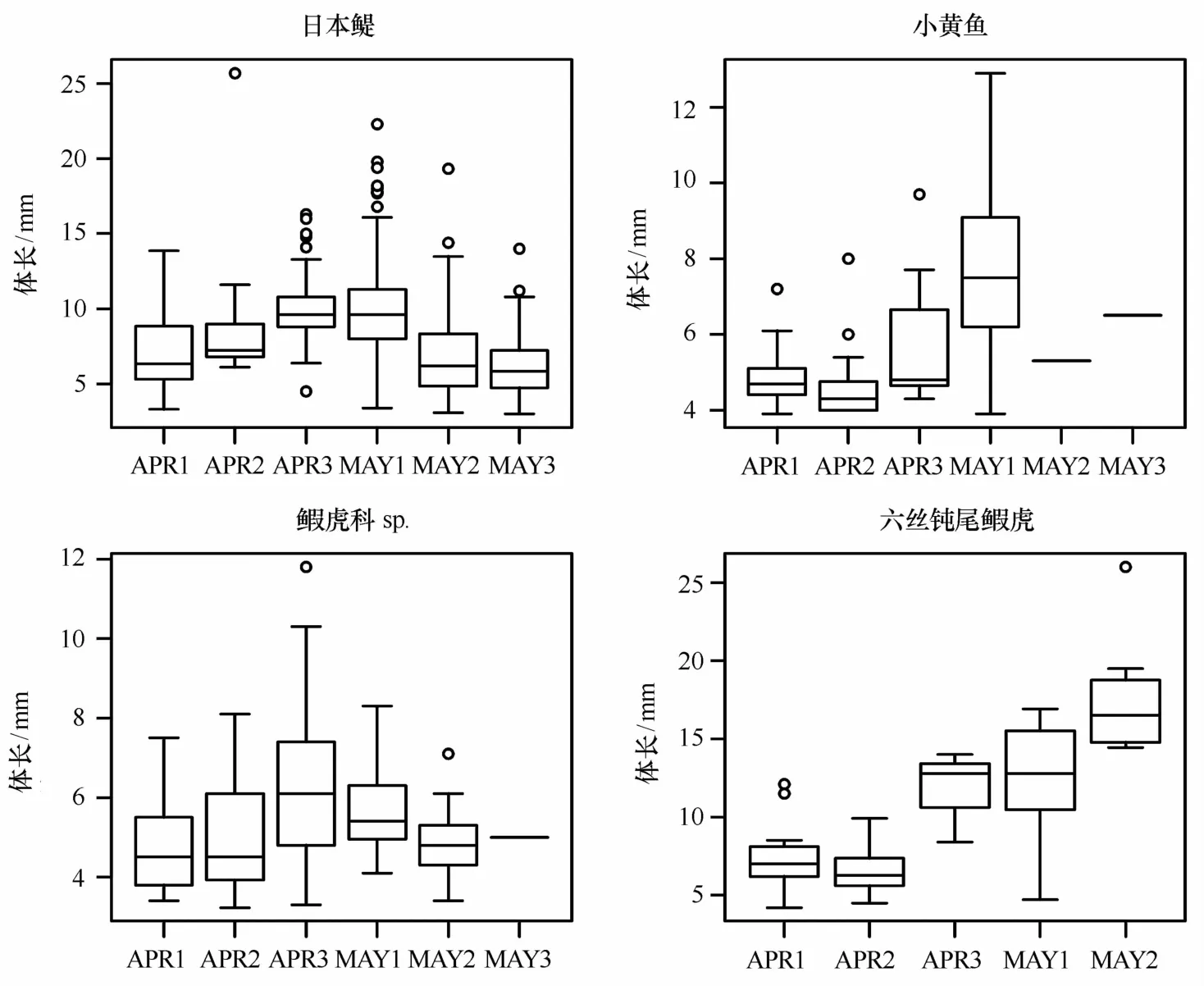
图6 优势种仔稚鱼的体长分布Eig.6 Body length distribution of dominant larval and juveni le fishes
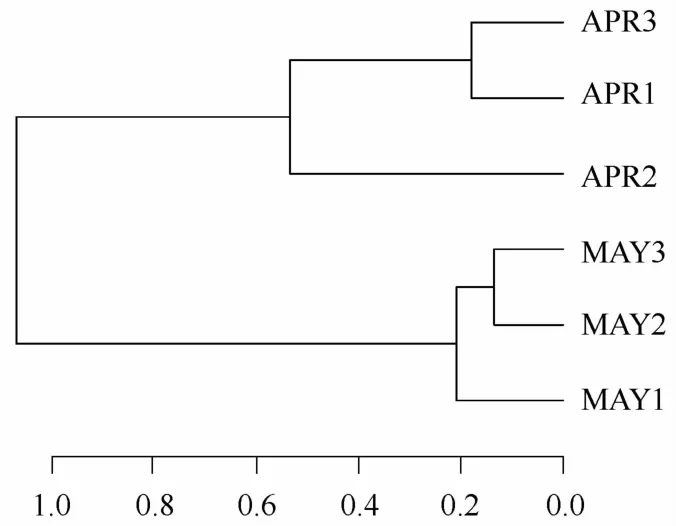
图7 航次层次聚类Eig.7 Hierarchical Cluster of cruises
4 讨论
万瑞景2001年春季的调查中东海北部(30°N以北,124°30′E以东)水域日本鳀鱼卵丰度很高,其仔稚鱼主要分布123°30′E以西的近岸水域[20]。胡芬2003年春季(4月上半)调查中发现长江口外123°E附近水域是日本鳀鱼卵仔鱼的集中分布区[21]。本研究的时空尺度较前人的研究小,结合他们的研究成果,可以进一步完善对日本鳀在东海北部近海的早期生活史的认识。日本鳀2月开始大量进入浙江近海产卵[22],鱼卵和前弯曲期仔鱼主要集中在123°E附近;在从前弯曲期向后弯曲期的生长发育过程中,日本鳀仔鱼向岸漂流,并有可能继续向海岸带漂流;5月中后期,在123°E附近出现另一次产卵高峰,大量鱼卵和早期仔鱼出现;6月份以后,由于水温升高,日本鳀开始游向外海继续产卵或开始索饵洄游[22—23],6月以后的日本鳀仔鱼的漂流方式还有待进一步研究发现。
陈佳杰等根据1971—1982年渔捞数据分析指出,小黄鱼春季在禁渔线内沿岸水域产卵,东海北部沿岸产卵场主要在长江口及邻近水域,产卵期为4月上旬到5月中旬[24]。林龙山等发现近年在外海海域也存在小黄鱼产卵场,其水温和水深分布范围较过去扩大[25—26]。本研究中的小黄鱼仔鱼以早期仔鱼为主,从4月初至5月初出现较多,分布在舟山群岛近岸水域,附近应该存在具一定规模的小黄鱼产卵场。小黄鱼仔鱼从4月至5月有明显的体长生长,仔鱼的平面分布具有较大的随机性且没有明显的移动趋势,显示出此水域在小黄鱼早期生活史中的保育作用。
几种优势种仔稚鱼占总渔获的大部分比例,不同航次间、断面或者区域间的种类组成均无大的差别。对前后航次的聚类只能在90%的置信水平上通过显著性检验,前几位贡献种的差别主要是前后丰度的变化,空间分布的聚类分析更是无法进行,因而在本研究的时空尺度上可以视调查水域的仔稚鱼为一个总体,没有分化出不同类群。
以往在长江口、东海北部和黄海南部的众多研究中,环境因子与鱼类浮游生物分布的相关性分析结果各不相同。在相对较大的时间或/与空间尺度上,水温、盐度对分布的作用明显,如水深、溶解氧、叶绿素、浮游动物等因子对鱼类浮游生物的影响则有所不同[15,23,27—31]。不同鱼类的繁殖生态习性、分布范围不同,本研究将几位优势种独立分析。在4月—5月,东海北部近海的日本鳀仔鱼在禁渔线附近孵化,大量早期仔鱼出现在此,本研究外侧站点临近日本鳀产卵水域,大量早期仔鱼在此聚集,因此回归分析结果表现出仔鱼丰度与水深正相关。较低的调整R方也暗示还有别的重要影响因子,可能即是成鱼产卵场位置。有学者提出过潮流导致日本鳀鱼卵聚集的假说[32],可能也是较为重要的影响因子。Clark等认为一定程度的浑浊度能帮助仔稚鱼有更大的几率躲避捕食者[33],日本鳀和小黄鱼仔鱼的丰度与透明度的负相关关系可能与此有关。本研究中小黄鱼在前期出现得多,水温较5月低,所以得出与温度负相关的结果。北面的断面受长江口冲淡水影响,外侧站位盐度较高,小黄鱼仔鱼也在外侧站位分布较多,所以显示出与盐度正相关。
2010年东海发现赤潮39次,累计面积6 374 k m2,5、6月发生次数最多,毒害作用较大的东海原甲藻引发的赤潮次数最多(18次)、面积最大(4 539 k m2),主要集中在浙江和福建沿岸海域[34]。本研究结果显示4、5月正是东海北部沿岸的日本鳀仔鱼向岸漂流,小黄鱼仔鱼在舟山群岛外侧进行保育,以及作为许多经济鱼类重要饵料的鰕虎仔稚鱼出现的季节。W hite等的实验中給初次开口的日本鳀和真鲷Pagrus major投喂一种有毒膝沟藻,日本鳀仔鱼的实验组和饥饿对照组的死亡率没有表现出显著差异;两者较大的仔鱼在摄食含有膝沟藻毒素的浮游动物后均表现出PSP中毒症状,20%~30%的仔鱼死亡[35]。Lefebvre等的实验则显示了仔鱼直接受溶解在水体内的藻毒素影响,体形和运动能力都会受影响[9]。在长江口外、舟山群岛邻近海域,有毒赤潮一旦暴发,既有可能直接导致仔稚鱼中毒或缺氧死亡,也可能在导致仔稚鱼身体麻痹不能顺利漂流到适宜继续生长发育的水域而大量死亡,日本鳀、小黄鱼、鰕虎科种类补充群体将受到不同程度的不良影响。
[1]Platt T,Euentes-Yaco C,Erank K T.M arine ecology:Spring algal bloom and larvalfish survival[J].Nature,2003,423(6938):398-399.
[2]Hjort J.Eluctuationsin the great fisheries of northern Europe viewed in thel ight of biologicalresearch[J].Rapp P-V Réun Cons Int Explor M er,1914,20:1-228.
[3]Cushing D H.Plankton production and year-class strength in fish populations——an update of the match/mismatch hypothesis[J].Advancesin Marine Biology,1990,26:249-293.
[4]W arlen S M,Tester P A,Colby D R.Recruitment oflarvalfishesinto a north carol ina estuary during a bloom ofthe red tide dinoflagel late,Gy mnodinium breve[J].Bul letin of M arine Science,1998,63(1):83-95.
[5]殷名称.鱼类早期生活史阶段的自然死亡[J].水生生物学报,1996,20(4):363-372.
Yin Mingcheng.Natural mortal ity of early l ife stages of fish[J].Acta H ydrobiologica Sinica,1996,20(4):363-372.
[6]Gossel in S,Eortier L,Gagné J A.Vulnerabi l ity of marine fish larvae to the toxicdinoflagel lateProtogonyaulax tamarensis[J].Mar Ecol Prog Ser,1989,57(1):1-10.
[7]Robineau B,Gagné J A,Eortier L,et al.Potentialimpact of a toxic dinoflagel late(Alexandrium excavatum)bloom on survival offish and crustacean larvae[J].Marine Biology,1991,108(2):293-301.
[8]蒋玫,缪宇平,袁骐,等.微小亚历山大藻对黑鲷早期生长发育的毒性效应[J].环境科学学报,2009,29(10):2152-2156.
Jiang Mei,Miu Yuping,Yuan Qi,et al.The effect ofAlexandrium minutumon development of eggs and larvae offishSparus macrocphalus[J].Acta Scientiae Circu mstantiae,29(10):2152-2156.
[9]Lefebvre K A,Trainer V L,Scholz N L.M orphological abnormal ities and sensorimotor deficitsin larvalfish exposed to dissolved saxitoxin[J].A-quatic Toxicology,2004,66(2):159-170.
[10]Wang J H,W u J Y.Occurrence and potentialrisks of harmful algal bloomsin the East China Sea[J].Science ofthe Total Environ ment,2009,407(13):4012-4021.
[11]张利永.东海大规模赤潮对微型浮游动物群落结构影响的研究[D].青岛:中国科学院海洋研究所,2007.
Zhang Liyong.The effects oflarge-scale harmfulalgal bloom in the East China Sea on microzooplankton com munity structure[D].Qingdao:Institute of Oceanology,Chinese Academy of Sciences,2007.
[12]蒋玫,沈新强.长江口及邻近水域夏季鱼卵、仔鱼数量分布特征[J].海洋科学,2006,30(6):92-97.
Jiang M ei,Shen Xiqiang.Abundance distributions of pelagicfish eggs and larvain the Changj iang River estuary and vicinity watersin su m mer[J]. Marine Sciences,2006,30(6):92-97.
[13]蒋玫,沈新强,王云龙,等.长江口及其邻近水域鱼卵、仔鱼的种项组成与分布特征[J].海洋学报,2006,28(2):171-174.
Jiang Mei,Shen Xinqiang,Wang Yunlong,et al.Species of fish eggs and larvae and distribution in Changj iang Estuary and vicinity water[J]. Haiyang Xuebao,2006,28(2):171-174.
[14]周永东,金海卫,蒋日进,等.浙江中北部沿岸春、夏季鱼卵和仔稚鱼种类组成与数量分布[J].水产学报,2011,35(6):881-889.
Zhou Yongdong,Jin Haiwei,Jiang Ri j in,et al.The category composition and abundance distributions ofichthyoplankton along the north-central coast of Zhej iang Province in spring and su m mer[J].Journal of Eisheries of China,2011,35(6):881-889.
[15]蒋玫,沈新强,陈莲芳.长江口及邻近水域春季鱼卵仔鱼分布与环境因子的关系[J].海洋环境科学,2006,25(2):37-44.
Jiang M ei,Shen Xinqiang,Chen Lianfang.Relationship between with abundance distribution offish eggs,larvae and environ mentalfactorsin the Changj iang Estuary and vicinity waters in spring[J].M arine Environmental Science,2006,25(2):37-44.
[16]W an R J,Zhou E,Shan X J,et al.Impacts of variabi l ity of habitatfactors on species composition ofichthyoplankton and distribution offish spawning ground in the Changj iang River estuary and its adjacent waters[J].Acta Ecologica Sinica,2010,30(3):155-165.
[17]Kendal l A W J,Ahlstrom E H,M oser H G.Early l ife history stages offishes and their characters[M]//M oser H G,Richards W J,Cohen D M,et al.Ontogeny and Systematics of Eishes.Lawrence:The A merican Society of Ichthyologists Herpetologists,1984.
[18]Nelson J S.Eishes of the W orld[M].H oboken,N J:John Wi ley,2006.
[19]伍汉霖,邵广昭,赖春福,等.拉汉世界鱼类系统名典[M].基隆:水产出版社,1999.
W u Hanl in,Shao Guangzhao,Lai Chunfu,et al.Latin-Chinese Dictionary of Eish Names by Classification System[M].Keelung:The Sueichan Press,1999.
[20]万瑞景,孙珊.黄、东海生态系统中鱼卵、仔稚幼鱼种类组成与数量分布[J].动物学报,2006,52(1):28-44.
W an Rui j ing,Sun Shan.The category composition and abundance ofichthyoplankton in the ecosystem of the Yel low Sea and the East China Sea[J].Acta Zoologica Sinica,2006,52(1):28-44.
[21]胡芬.2003年春季东海区浮性鱼卵和仔稚鱼种类组成及数量分布[J].海洋渔业,2004,26(2):79-85.
H u Een.The species composition and quantity distribution of pelagic eggs,fish larvae and juveni lesin the East China Sea regionin spring 2003[J]. Marine Eisheries,2004,26(2):79-85.
[22]薛利建,刘子藩.东海日本鳀数量分布和生物学特性的研究[J].浙江海洋学院学报(自然科学版),2005,24(4):312-317.
Xue Li j ian,Liu Zifan.Study on the biomass distribution&biological characteristics ofEngraulisjaponicusin the East China Sea[J].Journal of Zhej iang Ocean U niversity(Natural Science),2005,24(4):312-317.
[23]蒋玫,王云龙.东海夏季日本鲭(Scomber japonicus)和鳀鱼(Engraulis japonicus)鱼卵仔鱼分布特征[J].海洋与湖沼,2007,38(4):351-355.
Jiang Mei,Wang Yunlong.Su m mer distributions of eggs and larvae ofScomber japonicusandEngraulis japonicusin the East China Sea[J]. Oceanologia et Limnologia Sinica,2007,38(4):351-355.
[24]陈佳杰,徐兆礼,陈雪忠.我国沿海小黄鱼渔场的空间格局[J].水产学报,2010,34(2):236-244.
Chen Jiaj ie,Xu Zhaol i,Chen Xuezhong.The spatial distribution pattern of fishing ground for small yellow croaker in China Seas[J].Journal of Eisheries of China,2010,34(2):236-244.
[25]林龙山,程家骅,姜亚洲,等.黄海南部和东海小黄鱼(Larimichthys polyactis)产卵场分布及其环境特征[J].生态学报,2008,28(8):3486-3494.
Lin Longshan,Cheng Jiahua,Jiang Yazhou,et al.Spatial distribution and environmental characteristics of the spawning grounds of smal l yellow croakerin the southern Yellow Sea and the East China Sea[J].Acta Ecologica Sinica,2008,28(8):2386-3494.
[26]林龙山,姜亚洲,严利平,等.黄海南部和东海小黄鱼产卵亲体分布特征与繁殖力的研究[J].上海海洋大学学报,2009,18(4):453-459.
Lin Longshan,Jiang Yazhou,Yan Liping,et al.Study on the distribution characteristics and fecundity ofspawning stock ofLarimichthys polyactisin the southern Yel low Sea and the East China Sea[J].Journal of Shanghai Ocean University,2009,18(4):453-459.
[27]万瑞景,黄大吉,张经.东海北部和黄海南部鳀鱼卵和仔稚幼鱼数量、分布及其与环境条件的关系[J].水产学报,2002,26(4):321-330.
W an Rui j ing,H uang Daj i,Zhang Jing.Abundance and distribution of eggs and larvae ofEngraulisjaponicusin the Northern part of East China Sea and the Southern part of Yellow Sea and its relationship with environmental conditions[J].Journal of Eisheries of China,2002,26(4):321-330.
[28]刘淑德,线薇微.长江口及其邻近水域鱼类浮游生物群落的时空格局[J].生物多样性,2009,17(2):151-159.
Liu Shude,Xian W eiwei.Temporal and spatial patterns of the ichthyoplankton com munity in the Yangtze Estuary and its adjacent waters[J]. Biodiversity Science,2009,17(2):151-159.
[29]刘淑德,线薇微,刘栋.春季长江口及其邻近海域鱼类浮游生物群落特征[J].应用生态学报,2008,19(10):2284-2292.
Liu Shude,Xian W eiwei,Liu Dong.Characteristics ofichthyoplankton assemblagesin Yangtze Estuary and adjacent watersin spring[J].Chinese Journal of Appl ied Ecology,2008,19(10):2284-2292.
[30]李建生,胡芬,程家骅.长江口水域春季鱼卵仔鱼分布及其与温度盐度的关系[J].海洋科学,2007,31(4):16-21.
Li Jiansheng,H u Een,Cheng Jiahua.Distributions offish eggs and juveni les and relations between surfacelayer watertemperature and sal inity of the Yangtze estuary water area in spring[J].M arine Sciences,2007,31(4):16-21.
[31]蒋玫,李磊,沈新强,等.2010-2011年长江河口及邻近水域春夏季仔稚鱼群落结构及其多样性的研究[J].海洋学报,2014,36(6):131-137.
Jiang Mei,Li Lei,Shen Xinqiang,et al.Structure of com munity and biodiversity oflarvaefish in spring and autu mn 2010-2011 in Yangtze River Estuary and its adjacent waters[J].Haiyang Xuebao,2014,36(6):16-21.
[32]魏皓,王玉衡,万瑞景,等.黄海锋区环流与鳀鱼卵的聚集[J].中国海洋大学学报,2007,37(3):512-516.
W ei Hao,Wang Yuheng,Wang Rui j ing,et al.Tidalfront and the convergence of anchovy(Engraulisjaponicus)eggsin the Yel low Sea[J].Periodical of Ocean U niversity of China,2007,37(3):512-516.
[33]Clark B M,Bennett B A,Lamberth S J.Eactors affecting spatial variabi l ity in seine net catches of fish in the surf zone of Ealse Bay,South Africa[J].Marine Ecology Progress Series,1996,131(8):17-34.
[34]国家海洋局.2010年中国海洋环境质量公报[R].北京:国家海洋局,2010.
State Oceanic Administration.Report on the M arine Environ mental Qual ity in P.R.C,2010[R].Bei j ing:State Oceanic Administration,2010.
[35]W hite AW,Eukuhara O,Anraku M.M ortal ity of fish larvae from eating toxic dinoflagel lates or zooplankton containing dinoflagel late toxins[M]//Okaichi T,Anderson D M,Nemoto T.Red Tides:Biology,Environmental Science and Toxicology.Cal ifornia:Elsevier,1989:395-398.
Spatio-temporal distribution oflarval and juvenile fishes at frequent harmful algal blooms(H ABs)areas of the East China Sea in spring
Chen Yuange1,Zhong Junsheng2,Xu Zhaol i1,Chen Jiaj ie1
(1.Key and Open Laboratory of M arine and Estuary Eisheries,Ministry of Agriculture of China,East China Sea Eisheries Research Institute,Chineses Academy of Eisheries Sciences,Shanghai200090,China;2.Shanghai Ocean University,Shanghai201305,China)
Based on an oceanographic comprehensive survey at the frequent H A Bs area in the East China Sea from M arch 29th 2011 to 27th May 2011,species composition,spatio-temporal distribution and relationship with environmentalfactors were discussed.A total of 27 species were collected.The top abundant species includeEngraulis japonicas,Gobi idae sp.,A mblychaeturichthys hexanema,Larimichthys polyactis,comprising 91.23%of the total catches.According to the spatio-temporal distribution of abundance and develop mental stages composition,E.japonicasshowed two spawning peaks and indicated a trend ofinshore drifting,whileA.hexanemaandL.polyactis had only one spawning peak and uti l ized Zhoushan archipelago as nursery ground.These larval and juveni le fishes uti l ize this area for spawning,nursing and inshore drifting respectively,once H A Bs occur,the recruitment would be negatively affected.M ultivariate analysis showed no significant relationship between larval and juvenile fish assemblages and involved environmentalfactors,the location of spawning ground and nursing pattern may be more important reasons.
larvalfishes;spatio-temporal distribution;East China Sea;H A Bs
S931.1
A
0253-4193(2015)02-0044-11
2014-05-28;
2014-09-02。
国家重点基础研究发展计划(973计划)项目(2010CB428705)。
陈渊戈(1986—),男,四川省乐山市人,助理研究员,从事鱼类早期生活史研究。E-mai l:yg.chan.shou@gmai l.com
徐兆礼,研究员,主要研究浮游动物对全球气候变化的响应以及海洋灾害对海洋浮游生态系统的影响。E-mai l:xu_zhaol i@126.com
