根域体积限制对芹菜幼苗生长和气体交换及叶绿素荧光参数的影响
2015-07-04曲继松张丽娟冯海萍杨冬艳
曲继松,张丽娟,冯海萍,杨冬艳
(宁夏农林科学院 种质资源研究所,银川750002)
柠条(Caraganakorshinskii)是蝶形花科(Papilionaceae)锦鸡儿属(Caragana)植物栽培种的通称,落叶灌木。柠条作为中国“三北”地区一种广泛分布的乡土灌木树种,由于其根系发达、耐旱性强已成为水土保持和防风固沙的主要灌木树种[1-2]。对于多年生长的柠条,必须进行平茬抚育。如果不进行平茬,柠条就会出现严重的木质化现象。木质化柠条输送养分和水分的能力会越来越弱,然后逐渐干枯死亡。“三北”地区柠条种植面积广,占地面积大,估计全国柠条的生长面积至少在133.3万hm2以上,每年需要平茬的面积大约有33.3万hm2,这样将会产生大量的平茬枝条。前人对柠条的研究主要集中在生理特性[3-6]、环境条件对生长发育的影响[7-11]及 柠 条 利 用[12-13]等 方 面;本 课 题 组 在2009~2013年间以柠条平茬后的枝条经过粉碎的柠条粉作为育苗基质的探索性试验已经取得了初步成功,尤其是在西瓜[14]、甜瓜[15]、茄子[16]、辣椒[17]等育苗上取得较好表现。目前国外开发了椰子壳、锯末等替代草炭基质,并应用于商业化生产[18-19],国内在以木糖渣、芦苇末、油菜秸秆、蚯蚓粪等工农业废弃物为原料开发草炭替代基质方面也作了较为深入的研究[20-23];容器苗根域体积是固定的,容易对地下部分生长造成限制,进而影响地上部分的生长,基质与容器的筛选一直是国内外容器育苗研究的重要内容[24-30]。
目前柠条基质配型筛选研究已经基本确定了柠条基质的复混配比类型,但是使用柠条复混基质进行多种蔬菜育苗过程中的穴盘选择存在一定不确定性,因此本研究以芹菜为样本,不同穴数标准穴盘为试材,测定分析不同根域体积限制对芹菜幼苗生长、气体交换和叶绿素荧光动力学诱导曲线的影响,以判断何种规格穴盘更适宜柠条基质芹菜育苗,为柠条资源合理利用和工厂化育苗生产提供理论依据和技术支撑,提高沙产业的经济效益和生态效益。
1 材料和方法
1.1 试验地点
试验地点设在宁夏银川市宁夏农林科学院园林场试验基地育苗专用温室内,位于106°09′00.55″E,38°38′57.89″N,海拔1 117m。银川市属典型的中温带大陆性气候,四季分明,春迟夏短,秋早冬长,昼夜温差大,雨雪稀少,蒸发强烈,气候干燥,风大沙多等。年平均气温8.5℃左右,年平均日照时数2 800~3 000h,年平均降水量200mm 左右,无霜期185 d左右。
1.2 试验材料
供试芹菜品种为‘皇后’,引自于法国Tezier公司,供试柠条粉购自宁夏回族自治区盐池县源丰草产业有限公司,1m3柠条粉加入2.0kg尿素、5kg商品有机肥(N∶P∶K=12∶8∶9),高温静态发酵90d后,加入珍珠岩和蛭石(柠条粉∶珍珠岩∶蛭石=7∶2∶1,体积比),作为育苗基质使用。
1.3 试验设计
试验时间为2013年10月20日~2014年1月10日,试验在10m 大跨度育苗专用日光温室内进行。试验采用29cm×58cm 标准穴盘,设置7个不同规格的标准穴盘,以确保每个处理的根域体积、秧苗密度不同(具体规格见表1),每个处理重复3次。所有穴盘在育苗床架上摆放,每个穴盘按照南北方向摆放,所有穴盘横向并排一字排开,与温室前沿平行,距离温室前沿3m。将配制后的柠条粉基质装填入7种不同规格的育苗穴盘进行芹菜育苗试验,水分管理采用见干见湿方法管理,待处理Ⅲ的90%幼苗第五片真叶完全展开后进行幼苗的植株生长、气体交换及叶绿素荧光参数的测定。

表1 各处理基本状况Table 1 The basic situation of each treatment
1.4 测定项目及方法
1.4.1 生长指标 生长指标于出苗后第60 天测定。幼苗株高、根长用直尺测量,根系体积采用排水法测定,根系活力测定采用氯化三苯基四氮唑(TTC)法[31],根冠比=地下部干重(g)/地上部干重(g)[14],每个处理测量5株,样株随机选择。
1.4.2 气体交换参数 净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)、气孔限制值(Ls)、水分利用效率(WUE)等光合参数采用TPS-2便携式光合作用测定系统测定,测定时育苗温室内部光照强度为(1 000±50)μmol·m-2·s-1,CO2浓度为(400±20)μmol·mol-1。测定叶片为秧苗最高一片完全展开功能叶,每个处理测量3片叶片,随机选择取样。
1.4.3 叶绿素荧光参数 使用英国Hansatech公司生产的连续激发式荧光仪Handy PEA 测定芹菜幼苗叶片的荧光参数,测定前暗适应30 min,利用配套软件对数据进行处理分析。其中,初始荧光(F0)、最大荧光(Fm)、可变荧光(Fv)、PSⅡ最大光化学效率(Fv/Fm)、性能指数(PI)直接从系统导出,其余参数根据公式计算。主要包括:单位反应中心吸收的光能(ABS/RC)=Mo·(1/Vj)·(1/φPo),单位反应中心捕获的用于还原QA的能量(TRo/RC)=Mo·(1/Vj),单位反应中心捕获的用于电子传递的 能量(ETo/RC)=Mo·(1/Vj)·ψo,单位反应中心耗散掉的能量(DIo/RC)=(ABS/RC)-(TRo/RC);初始最大光化学效率(φPo)=TRo/ABS,捕获的激子中用来推动电子传递到电子传递链中超过QA的其它电子受体的激子占用来推动QA还原激子的比率(ψo)=ETo/TRo,用于电子传递的量子产额(φEo)=ETo/ABS,用于热耗散的量子比率(φDo)=1-φPo;单位面积吸收的光能(ABS/CS)≈F0,单位面积内有活性的反应中心数目(RC/CS)=φPo(Vj/Mo)·(ABS/CS),单位面积捕获的光能(TRo/CS)=φPo·(ABS/CS),单位面积内用于电子传递的光能(ETo/CS)=φEo·(ABS/CS),单位面积内热耗散的光能(DIo/CS)=(ABS/CS)-(TRo/CS)[32]。测定于晴天上午10:00~12:00之间进行,测定叶片为秧苗最高一片完全展开功能叶,每个处理测量3片叶片,样品随机选择。
1.5 数据分析
每个样本测量3次,结果取平均值。数据处理和作图采用DPS 3.01软件Duncan新复极差法和Excel软件进行统计分析。
2 结果与分析
2.1 根域体积限制对芹菜幼苗生长和生物量积累的影响
从表2可以看出,芹菜幼苗株高、叶片数、根长、根系体积和根系活力均随着根域体积的减小而逐渐降低,且处理之间大多达到显著差异水平,如处理Ⅶ的各指标分别比处理Ⅰ显著降低38.68%、42.86%、63.83%、77.14%、58.00%。可见,根域体积限制显著抑制了芹菜幼苗的生长,并且对幼苗根系指标的影响更大,尤其是根系体积。
同时,根域体积限制也对芹菜幼苗干物质积累产生了显著的影响(表3)。其中,幼苗地上部干鲜质量、地下部干鲜质量、全株干鲜质量基本都随根域体积的减小而逐渐降低,而其根冠比却表现出现增加后降低的趋势,并在处理Ⅲ时达到最大值,且各处理之间大多达到显著差异水平。如处理Ⅶ以上各生物量比处理Ⅰ显著降低79.01%、70.9%、79.67%、71.15%、79.15%、70.95%,以上结果说明,根域体积限制也显著抑制了芹菜幼苗生物量的积累。
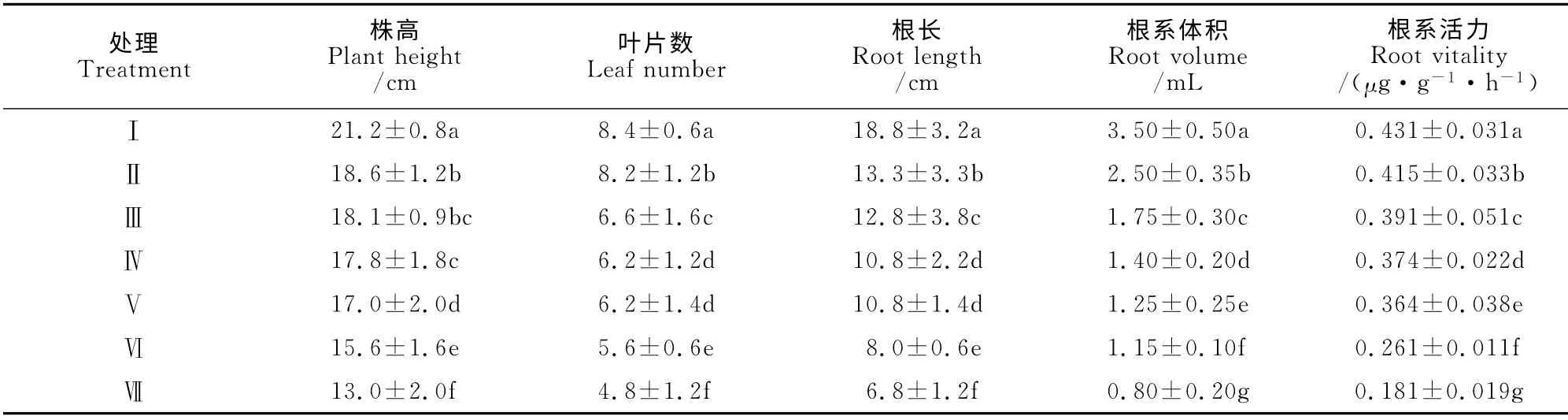
表2 根域体积限制对芹菜幼苗生长的影响Table 2 Effect of root-zone volume on growth of A.graveolens L.seedling in Caragana substrate
2.2 根域体积对芹菜幼苗叶片气体交换参数影响
根域体积大小对芹菜幼苗叶片气体交换参数的部分参数影响达到差异显著水平(表4)。在净光合速率方面总体变化趋势均为随着根域体积的减少而各参数减少,各处理之间达到差异显著水平,处理Ⅶ以上各净光合速率比处理Ⅰ显著降低39.07%、26.21%、22.20%、17.78%、21.08%和1.82%;各处理蒸腾速率差异显著,变化规律与净光合速率类似,处理Ⅶ的蒸腾速率比处理Ⅰ降低了15.87%。蒸腾速率亦随着根域体积的减小而降低,在气孔导度也是如此,总体趋势为随着根域体积的减少而减少。
在胞间CO2浓度方面,处理Ⅲ、处理Ⅴ、处理Ⅵ之间差异不显著,处理Ⅱ和处理Ⅳ差异不显著,处理Ⅰ至处理Ⅶ总体变化趋势无显著规律;在气孔限制值方面,各处理之间差异显著,总体变化趋势不规律;在各处理之中,处理Ⅱ的水分利用效率值最高,为2.628 6,处理Ⅶ为1.816 9,为处理Ⅰ的69.12%,其它处理均在2.000 0~2.599 9之间,与根域体积不构成相关关系。
2.3 根域体积对芹菜幼苗叶绿素荧光参数的影响
2.3.1 基本参数 不同根域体积限制下芹菜幼苗叶片叶绿素荧光基本参数F0、Fm和Fv变化较大(表5),且各处理达到差异显著水平,其中处理Ⅲ的F0值最大,达到546.75,比处理Ⅶ高出7.63%,但与根域体积不构成相关关系。Fm和Fv在处理Ⅳ时值最高,分别为2 608.25和2 074.5,当根域体积大于或小于26.82cm3/穴(处理Ⅳ)时,Fm和Fv均呈现逐渐降低的趋势,各处理之间达到差异显著水平。
Fv/Fm是表明光化学反应状况的1 个重要参数,Fv/Fm反映了荧光诱导动力学曲线上升过程的O-P段的PSⅡ光合电子传递能力,从其大小关系可以发现根域体积过大或偏小都会影响PSⅡ光合电子传递,而且根域体积过小(处理Ⅶ,7.28cm3/穴)对PSⅡ光合电子传递的影响要明显大于根域体积过大(处理Ⅰ,110.07cm3/穴)的影响;PI可以准确反映植物光合机构的状态,适当根域体积(处理Ⅳ,26.82cm3/穴)芹菜幼苗光合原初反应显著高于根域体积过大(处理Ⅰ,110.07cm3/穴)或偏小(处理Ⅶ,7.28cm3/穴)。
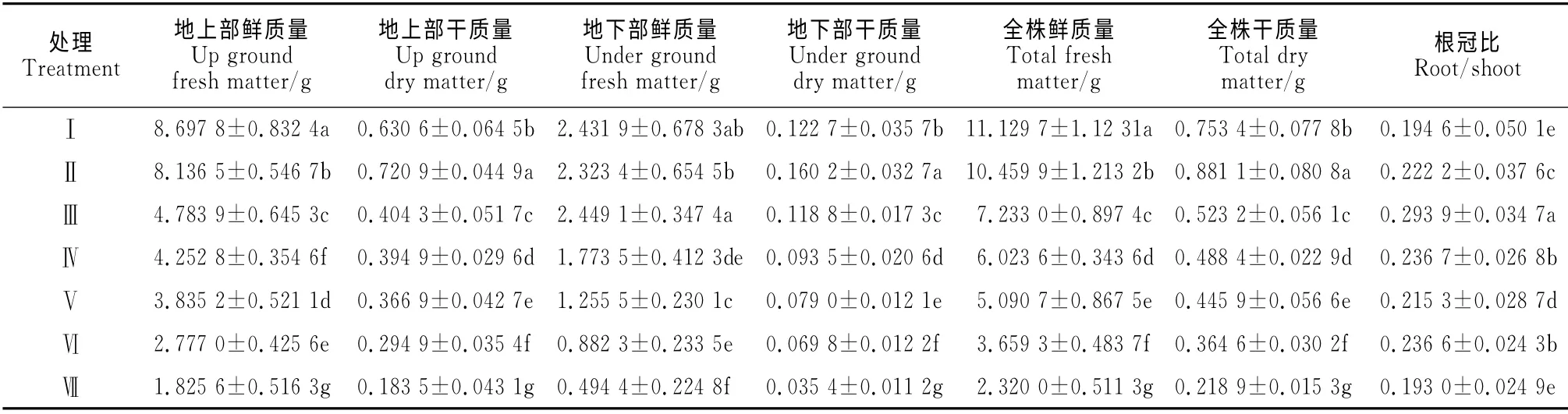
表3 根域体积限制对芹菜幼苗干物质积累的影响Table 3 Effect of root-zone volume on accumulation of dry matter of A.graveolens L.seedling in Caragana substrate

表4 根域体积限制对芹菜幼苗叶片气体交换参数的影响Table 4 Effect of root-zone volume on gas exchange parameters in leaves of A.graveolens L.seedling in Caragana substrate
2.3.2 光系统Ⅱ反应中心活性参数 由表6可以看出,芹菜幼苗叶片单位反应中心吸收(ABS/RC)、捕获的用于还原QA(TRo/RC)、捕获的用于电子传递(ETo/RC)及热耗散掉(DIo/RC)的能量的4个活性参数与根域体积既不呈现线性相关,也不呈现非线性相关,说明光系统Ⅱ(PSⅡ)反应中心活性参数受根域限制的影响不大。但无论是何种根域体积条件下都呈现出ABS/RC>TRo/RC>ETo/RC的趋势,这表明随着电子传递链的延伸热耗散增加,光能利用率降低。而且处理Ⅰ和处理Ⅶ的ETo/RC均比DIo/RC小,其 余 各 处 理 均 为ETo/RC>DIo/RC,说明根域体积过大或偏小均使得PSⅡ反应中心用于热耗散的能量高于用于电子传递的能量。
2.3.3 光系统Ⅱ能量分配比率 在PSⅡ受体侧的几个指标中(表7),初始最大光化学效率(φPo)、用于热耗散的量子比率(φDo)与根域体积既不呈线性相关,也不呈现非线性相关,PSⅡ的功能活性受根域体积影响不大;用于电子传递的量子产额(φEo)和捕获的激子中用来推动电子传递到电子传递链中超过QA的其它电子受体的激子占用来推动QA还原激子的比率(ψo)主要反映了PSⅡ受体侧的变化,φEo与根域体积既不呈线性相关,也不呈现非线性相关,ψo与φEo变化规律极其相似;说明根域体积过大(处理Ⅰ,110.07cm3/穴)或偏小(处理Ⅶ,7.28cm3/穴)使得芹菜幼苗叶片用于QA下游电子传递的量子不断减少,PSⅡ反应中心捕获的激子中用于QA下游电子传递的激子占捕获激子总数的比例不断减少,PSⅡ受体侧QA下游的电子传递接收的能量占总能量的比例值都是不断降低的。
2.3.4 光系统Ⅱ光能利用效率 对叶绿素快速荧光诱导动力学曲线的数据分析表明,处理Ⅴ的单位面积内反应中心数目(RC/CS)值最高,且各处理达到差异显著水平,比最低的处理Ⅶ高出31.68%,同时处理Ⅴ的单位面积捕获的光能(TRo/CS)比最低值处理Ⅶ高出8.67%;而单位面积内用于电子传递的光能(ETo/CS)处理Ⅴ值仍然为最高值,同样处理Ⅶ最低,仅为处理Ⅴ的74.90%;单位面积内热耗散的光能(DIo/CS)方面,处理Ⅴ最小,比最高值处理Ⅶ降低9.26%。
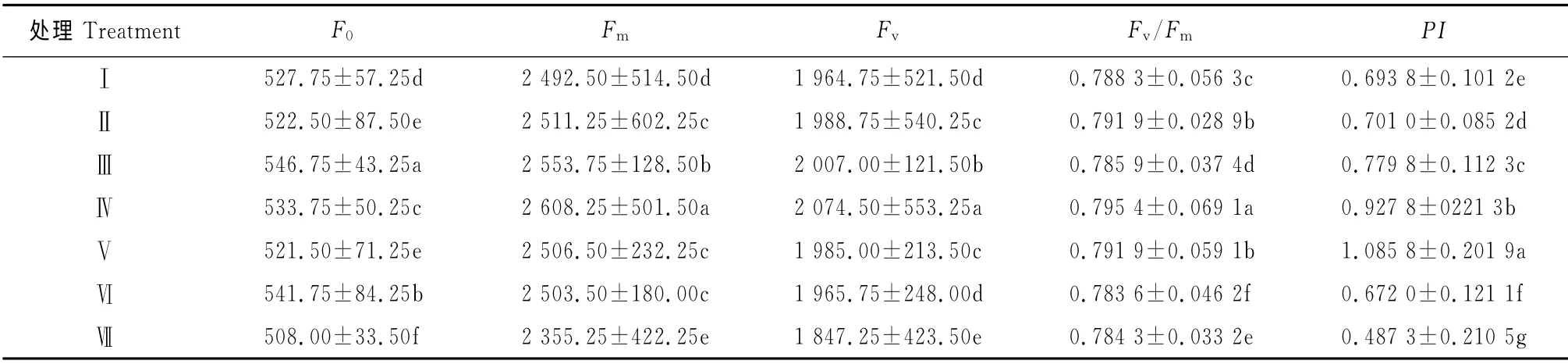
表5 根域体积限制对芹菜幼苗叶绿素荧光参数的影响Table 5 Effect of root-zone volume on chlorophyll fluorescence parameters in leaves of A.graveolens L.seedling in Caragana substrate

表6 根域体积限制对芹菜幼苗叶片PSⅡ反应中心活性参数的影响Table 6 Effect of root-zone volume on specific energy fluxes of PSⅡreaction center of A.graveolens L.seedling in Caragana substrate

表7 根域体积限制对芹菜幼苗叶片光系统Ⅱ能量分配比率的影响Table 7 Effect of root-zone volume on flux ratios of PSⅡreaction center of A.graveolens L.seedling in Caragana substrate
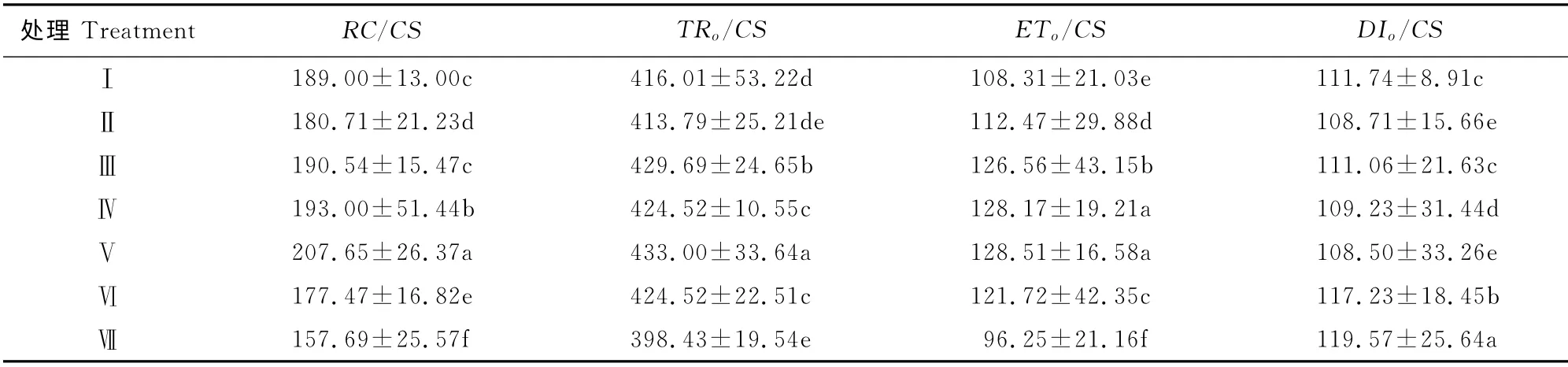
表8 根域体积限制对芹菜幼苗叶片单位面积叶片光能利用效率的影响Table 8 Effect of root-zone volume on light use efficiency per unit leaf area of maize ear leaf of A.graveolens L.seedling in Caragana substrate
3 讨 论
3.1 根域体积限制对柠条基质芹菜幼苗生长和干物质积累的影响
无论是何种苗龄定植,均表现为穴盘孔数越少,蔬菜幼苗生长势越强[33],孙磊玲等认为高密度低根域体积的栽培,虽然可以使经济学产量较高,但商品性较差[34];本研究得出:株高随着根域体积的减小而降低;叶片数、根长、根系体积表现结果与株高相同,说明根域体积大小对芹菜幼苗株高、叶片数、根长、根系体积等形态指标的影响大于秧苗密度的影响,而且从试验结果分析得出,根域体积越大对芹菜幼苗形态指标越有利,这与张海利等的结果相一致[35]。
3.2 根域体积限制对柠条基质芹菜幼苗气体交换参数的影响
根域体积限制对芹菜幼苗叶片气体交换参数的影响差异显著,孙磊玲等研究认为净光合速率随着单株根域体积的减小呈现递减趋势,高密度栽培会影响普通幼苗对光能的吸收、营养的分配,进而影响其叶绿素的合成和叶面积的增大[34],这与本试验研究结果相一致。
植物通过光合作用合成碳水化合物,积累干物质,积累量的大小直接反映在植株的生长量上。光合作用是作物形成生物学产量和经济产量的基础。光合强度不但与叶片的生理状况有关,而且和根系的发育密切相关。虽有限根对植株光合无影响的报道[36],但极度限根会使光合速率下降[37]。
3.3 根域体积限制对柠条基质芹菜幼苗叶绿素荧光参数的影响
叶绿素荧光测定技术作为一种无损伤的快速探针用于植物的抗逆生理研究已有大量报道[38-41]。Fv/Fm反映了荧光诱导动力学曲线上升过程的O-P段的PSⅡ光合电子传递能力(表5),根域体积过大(处理Ⅰ,110.07cm3/穴)或 偏 小(处 理Ⅶ,7.28 cm3/穴)都会影响PSⅡ光合电子传递,而且根域体积过小(处理Ⅶ,7.28cm3/穴)对PSⅡ光合电子传递的影响要明显大于根域体积过大(处理Ⅰ,110.07 cm3/穴)的影响,这可能是由于根域体积较小时,根域体积与秧苗密度共同作用的结果,但其影响的主效因子的确定有待于进一步研究。
根据Strasser等[42]的能量流动模型,植物叶片吸收的总能量(ABS),一部分以荧光的形式释放,大部分被反应中心(RC)捕获(TR),被反应中心捕获的能量中有一部分通过QA的还原氧化导致电子传递(ET);另一部分以热耗散的形式释放(DI)。ABS/RC、TRo/RC和DIRo/RC的值不断增加,说明叶片受胁迫时单位反应中心承担的光能转换任务更多[43]。本试验得出芹菜幼苗叶片PSⅡ反应中心活性 参 数ABS/RC、TRo/RC、ETo/RC及DIo/RC受根域限制的影响不大,但无论是何种根域体积条件下都呈现出ABS/RC>TRo/RC>ETo/RC的趋势,这表明随着电子传递链的延伸热耗散增加,光能利用率降低;而且处理Ⅰ和处理Ⅶ的ETo/RC均比DIo/RC小,其它各处理均为ETo/RC>DIo/RC,说明根域体积过大或偏小均使得PSⅡ反应中心用于热耗散的能量高于用于电子传递的能量,这也体现了热耗散对PSⅡ具有较强的保护能力[44]。
用于电子传递的量子产额(φEo)和捕获的激子中用来推动电子传递到电子传递链中超过QA的其它电子受体的激子占用来推动QA还原激子的比率(ψo)主要反映了PSⅡ受体侧的变化,ψo是对PSⅡ电子传递的综合评价之一,受PSⅡ供体侧的电子供应能力和受体侧(包括PSⅠ)接收电子的能力制约[45]。当根域体积为25.68cm3/穴时,φEo值达到最大;当根域体积达到14.34cm3/穴时,ψo值达到最大。说明根域体积超过25.68cm3/穴或小于14.34cm3/穴时,使得芹菜幼苗叶片用于QA下游电子传递的量子不断减少,PSⅡ反应中心捕获的激子中用于QA下游电子传递的激子占捕获激子总数的比例不断减少,PSⅡ受体侧QA下游的电子传递接收的能量占总能量的比例值都是不断降低的。
当根域体积达到17.32cm3/穴时,TRo/CS值达到最大;当根域体积达到19.21cm3/穴时,ETo/CS值达到最大。说明根域体积超过19.21cm3/穴或小于17.32cm3/穴时,芹菜叶片单位面积叶片从光能捕获(TRo/CS)和用于电子传递(ETo/CS)的能力开始下降,并导致光能过剩及活性氧浓度上升,损害OEC[46]。
在穴盘规格(29cm×58cm)确定的前提下,根域体积、穴盘规格、秧苗密度三者相互制约,本研究结果是在三者同时作用下得到的结论,而三者对芹菜幼苗生长发育、光合及叶绿素荧光参数方面的单效影响,以及互作关系过程中主效因素的确定有待于进一步研究。
4 结 论
根域体积大小对芹菜幼苗株高、叶片数、根长、根系体积、地上部鲜质量、地上部干质量、地下部鲜质量和地下部干质量等指标的影响较大。在净光合速率随着根域体积的减少而各参数减少,净光合速率与根域体积呈线性正相关,蒸腾速率变化规律与净光合速率相似,气孔导度总体变化趋势为随着根域体积的减少而减少,胞间CO2浓度总体变化受根域体积影响不大。根域体积限制会影响PSⅡ光合电子传递,适当根域体积时(26.82cm3/穴)性能指数(PI)值较高。根域体积过大(110.07cm3/穴)或偏小(7.28cm3/穴)均使得PSⅡ反应中心用于热耗散的能量高于用于电子传递的能量。当根域体积超过25.68cm3/穴或小于14.34cm3/穴时,使得芹菜幼苗叶片用于QA下游电子传递的量子不断减少,PSⅡ反应中心捕获的激子中用于QA下游电子传递的激子占捕获激子总数的比例不断减少,PSⅡ受体侧QA下游的电子传递接收的能量占总能量的比例值不断降低。根域体积超过19.21cm3/穴或小于17.32cm3/穴时,芹菜叶片单位面积叶片从光能捕获(TRo/CS)和用于电子传递(ETo/CS)的能力开始下降。通过对芹菜幼苗生长、气体交换和叶绿素荧光参数的综合分析得出,建议在使用柠条基质穴盘培育芹菜时使用128穴/盘,即根域体积为19.76 cm3/穴的穴盘进行育苗操作。
[1]ZHANG ZH W(张振万),YANG SH X(杨淑性).Plant ofCaraganaFavr.in Shaanxi[J].ActaBotanicaBoreali-OccidentaliaSinica(西北植物研究),1983,3(1):21-31(in Chinese).
[2]ZENG CH(曾 辰),SHAO M A(邵明安).Soil moisture variation of youngCaraganakorshinskiiartificial shrubland in the wind-water erosion crisscross region of the Loess Plateau[J].AgriculturalResearchintheAridAreas(干旱地区农业研究),2006,24(6):155-158(in Chinese).
[3]MA C C,GAO Y B,GUO H Y,etal.Photosynthesis,transpiration,and water use efficiency ofCaraganamicrophylla,C.intermedia,andC.korshinskii[J].Photosynthetica,2004,42(1):65-70.
[4]ZHENG Y R,RIMMINGTON G M,XIE Z X,etal.Responses to air temperature and soil moisture of growth of four dominant species on sand dunes of central Inner Mongolia[J].J.PlantRes.,2008,121(5):473-482.
[5]FANG X W,LI Y B,XU D H,etal.Activities of starch hydrolytic enzymes and starch mobilization in roots ofCaraganakorshinskiifollowing above-ground partial shoot removal[J].Trees,2007,21(1):93-100.
[6]ZHANG Z S,LI X R,LIU L C,etal.Distribution,biomass,and dynamics of roots in a revegetated stand ofCaraganakorshinskiiin the Tengger Desert,northwestern China[J].J.PlantRes.,2009,122(1):109-119.
[7]CHENG X R,HUANG M B,SHAO M G,etal.A comparison of fine root distribution and water consumption of matureCaraganakorshinkiiKom grown in two soils in a semiarid region,China[J].PlantSoil,2009,315(1/2):149-161.
[8]ALAMUSAANDl,JIANG D M.Characteristics of soil water consumption of typical shrubs(Caraganamicrophylla)and trees(Pinus sylvestris)in the Horqin Sandy Land area,China[J].FrontForChina,2009,4(3):330-337.
[9]WANG Z Q,LIU B Y,LIU G.Soil water depletion depth by planted vegetation on the Loess Plateau[J].Sci.ChinaSerD-EarthSci.,2009,52(6):835-842.
[10]ZHENG Y R,XIE Z X,GAO Y,etal.Germination responses ofCaraganakorshinskiiKom.to light,temperature and water stress[J].EcologicalResearch,2004,19(5):553-558.
[11]YIN J,HE F,QIU G Y,etal.Characteristics of leaf areas of plantations in semiarid hills and gully loess regions[J].FrontForChina,2009,4(3):351-357.
[12]LI X R,KONG D S,TAN H J,etal.Changes in soil and vegetation following stabilization of dunes in the southeastern fringe of the Tengger Desert,China[J].PlantSoil,2007,300(1/2):221-231.
[13]AWANG Y,ISMAILl M.The growth and flowering of some annual ornamentals on coconut dust[J].ActaHort.,1997,450(2):31-38.
[14]QU J S(曲继松),GUO W ZH(郭文忠),ZHANG L J(张丽娟),etal.Influence on the growth and accumulation of dry matter of watermelon seedlings based onCaragana-straw as nursery substrate[J].TransactionsoftheCSAE(农业工程学报),2010,26(8):291-295(in Chinese).
[15]ZHANG L J(张丽娟),QU J S(曲继松),FENG H P(冯海萍),etal.Influence on the quality of muskmelon seedlings utilize caraganastraw as nursery substrate[J].NorthernHorticulture(北方园艺),2010,(15):165-167(in Chinese).
[16]QU J S(曲继松),ZHANG L J(张丽娟),FENG H P(冯 海萍),etal.Influence of caragana-straw as component of mixed substrate on growth of eggplant seedlings[J].ActaAgriculturaeBoreali-OccidentalisSinica(西北农业学报),2012,21(11):162-167(in Chinese).
[17]QU J S(曲继松),ZHANG L J(张丽娟),FENG H P(冯海萍),etal.Caragana-straw as component of mixed substrate for pepper seedling growth[J].JiangsuJ.ofAgr.Sci.(江苏农业学报),2012,28(4):846-850(in Chinese).
[18]OSTOS J C,LóPEZ G R,MURILLO J M,etal.Substitution of peat for municipal solid waste and sewage sludge-based composts in nursery growing media:Effects on growth and nutrition of the native shrubPistacialentiscusL.[J].BioresourceTechnology,2008,99(6):1 793-1 800.
[19]GRUDA N,SCHNITZLER W H.Suitability of wood fiber substrates for production of vegetable transplantsⅡ[J].ScientiaHorticulturae,2004,100:333-340.
[20]LAN SH L(兰时乐),CAO X ZH(曹杏芝),DAI X Y(戴小阳),etal.The changes of nutrition elements during the composting chicken manure and rape straw under higher temperature[J].JournalofAgro-EnvironmentScience(农业环境科学学报),2009,28(3):564-569(in Chinese).
[21]CHENG F(程 斐),SUN ZH H(孙朝晖),ZHAO Y G(赵玉国),etal.Analysis of physical and chemical properties of reed residue substrate[J].JournalofNanjingAgriculturalUniversity(南京农业大学学报),2001,24(3):19-22(in Chinese).
[22]SHANG X H(尚秀华),XIE Y J(谢耀坚),PENG Y(彭 彦).Rice-husk composting as seedling medium boosted by sugar refinery wastewater[J].JournalofCentralSouthUniversityofForestry&Technology(中南林业科技大学学报),2009,29(2):78-81(in Chinese).
[23]SHANG Q M(尚庆茂),ZHANG ZH G(张志刚).Experimental studies on fertilizer-adding amount in eggplant plug seedling production with vermicom post-based media[J].TransactionsoftheCSAE(农业工程学报),2005,21(S):129-132(in Chinese).
[24]DENG Y(邓 煜),LIU ZH F(刘志峰).Study on growth medium and growth law for containerized seeding stocks grown in Greenhouse[J].ScientiaSilvaeSinicae(林业科学),2000,36(5):33-39(in Chinese).
[25]GINWAL H S,RAWAT D S,SHARMA S,etal.Standardization of proper volume/size and type of root trainer for raisingAcacianiloticaseedlings:Nursery evalution and field trail[J].IndianForestry,2001,127:920-928.
[26]BASHIR A,QAISAR K N,KHAN M A,etal.Standardization of growing media for raisingPinuswallichianaseedlings under root trainer production system in nursery[J].EnvironmentandEcology,2009,27(1A):381-384.
[27]AGBOGIDI O M,ENUJEKE E C,ESHEGBEYI O F.Germination and seedling growth of African pear(DacryodesedulisDon.G.Lam.H.J.)as affected by different planting media[J].AmericanJournalofPlantPhysiology,2007,2(4):282-286.
[28]GOPAL S,SUMIT C,DEY A N.Effect of growing media on germination and initial seedling growth ofAlbiziaprocera(Roxb)Benth.in Terai zone of West Bengal[J].EnvironmentandEcology,2007,25(S2):406-407.
[29]WANG Y SH(王月生),ZHOU ZH CH(周志春),JIN G Q(金国庆),etal.Growth ofTaxuschinensisvar.maireifor container seedlings in different media mixtures and for bare-root versus container seedlings in a young stand[J].JournalofZhejiangForestryCollege(浙江林学院学报),2007,24(5):643-646(in Chinese).
[30]ZHANG J M(张纪卯).Studies on effect of different medium and container size on growth ofKeteleeriafartuneicontainer seedling?[J].JournalofFujianCollegeofForestry(福建林学院学报),2001,21(2):176-180(in Chinese).
[31]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000:207-208.
[32]LI P M(李鹏民),GAO H Y(高辉远),RETO J S.Application of the chlorophyll fluorescence induction dynamics in photosynthesis study[J].JournalofPlantPhysiologyandMolecularBiology(植物生理学与分子生物学学报),2005,31(6):559-566(in Chinese).
[33]CHEN H(陈 慧),LIANG ZH H(梁朝晖),XIE Y Q(谢燕青),etal.Effects of different specification plug seedling on growth and yield of Chinese cabbage[J].JournalofChangjiangVegetables(长江蔬菜),2011,12:38-40(in Chinese).
[34]SUN L L(孙磊玲),HUANG D F(黄丹枫),ZHANG K(张 凯),etal.Effects of rooting-zone volume on packchoi growth in greenhouse[J].ChinaVegetables(中国蔬菜),2012,18:116-121(in Chinese).
[35]ZHANG H L(张海利),SUN J(孙 娟),PANG Z Q(庞子千).Effects of different specifications of tray on growth and development of tomato seedlings[J].JournalofChangjiangVegetables(长江蔬菜),2012,8:42-43(in Chinese).
[36]KHARKINA T G,ROSENQVIST E,OTTOSEN C O.Effects of root restriction on the growth and physiology of cucumber plants[J].PhysiologiaPlantarum,1999,105(3):434-441.
[37]WILL R E,TESKEY R O.Effect of elevated carbon dioxide concentration and root restriction on net photosynthesis,water relations and foliar carbohydrate status of Ioblolly pine seedlings[J].TreePhysiol.,1997,17(10):655-661.
[38]GUO Y P(郭延平),ZHOU H F(周慧芬),ZENG G H(曾光辉),etal.Effects of high temperature stress on net photosynthetic rate and photosystem activity inCitrus[J].ChineseJournalofAppliedEcology(应用生态学报),2003,14(6):867-870(in Chinese).
[39]WU H Y(吴韩英),SHOU S Y(寿森炎),ZHU ZH J(朱祝军),etal.Effects of high temperature stress on photosynthesis and chlorophyll fluorescence in sweet pepper[J].ActaHorticultureSinica(园艺学报),2001,28(6):517-521(in Chinese).
[40]YAMANE Y,KASHINO Y,KOIKE H,etal.Increases in the fluorescence level andF0level and reversible inhibition of photosystem Ⅱreaction center by high temperature treatment in higher plants[J].PhotosynthesisResearch,1997,52:57-64.
[41]YOSHIHIRO Y,YASUHIRO K,HIROYUKI K,etal.Effects of high temperatures on the photosynthetic systems in spinach:Oxygenevolving activities,fluorescence characteristics and the denaturation process[J].PhotosynthesisResearch,1998,57:51-59.
[42]STRASSER R J,TSIMILLI M M,SRIVASTAVA A.The Fluorescence Transient as A Tool to Characterise and Screen Photosynthetic Samples[M]//YUNUS M,PATHRE U,MOHANTY E.Probing Photosynthesis:Mechanisms,Regulation and Adaptation.London:Taylor &Francis,2000.
[43]HAN B(韩 彪),CHEN G X(陈国祥),GAO ZH P(高志萍),etal.The changes of PSⅡchlorophyll fluorescence dynamic characteristic during leaf senescence of ginkgo[J].ActaHorticulturaeSinica(园艺学报),2010,37(2):173-178(in Chinese).
[44]WANG M(王 梅),GAO ZH K(高志奎),HUANG R H(黄瑞虹),etal.Heat stress characteristics of photosystemⅡin eggplant[J].ChineseJournalofAppliedEcology(应用生态学报),2007,18(1):63-68(in Chinese).
[45]KRAUSE G H,WEIS E.Chlorophyll fluorescence and photosynthesis:The basis[J].Annu.Rev.PlantPhysiol.PlantMol.Biol.1991,42:313-349.
[46]LI G(李 耕),GAO H Y(高辉远),ZHAO B(赵 斌),etal.Effects of drought stress on activity of photosystems in leaves of maize at grain filling stage[J].ActaAgronomicaSinica(作物学报),2009,35(10):1 916-1 922(in Chinese).
