澳洲杨(大戟科)胚珠及其种子附属结构的个体发育研究
2015-07-04THOMASSttzel张志翔于海燕王利兵
张 鑫,THOMAS Stützel,张志翔,于海燕,王利兵
(1 西北农林科技大学 林学院,陕西杨陵712100;2波鸿鲁尔大学 植物多样性与进化系 生物科学与技术学院,德国波鸿44780;3北京林业大学 自然保护区学院,北京100083;4 中国林业科学研究院 林业研究所,北京100091)
种子附属物的结构和发育呈现出各种不同的模式和生物学功能[1]。如果这些结构被用来进行进化生物学研究,一个清晰的限定和区别不同类型的种子附属物是关键,特别是一些类群中存在描述不完整,甚至是相互矛盾的情况。澳洲杨属植物就是一个非常好的例子(图1)。
Corner[2]记载了Homalanthuspopulneus(Geiseler.)Pax的胚珠和种子形态,描述其种子没有附属结构,而Esser[3]在关于马来西亚的澳洲杨属植物修订时,记载澳洲杨属植物种子具有类似于假种皮的种子附属结构。后来Gardner[4]描述澳大利亚的澳洲杨(HomalanthuspopulifoliusGraham)的果实及种子结构时认为这个属的种子有乳凸状的种皮和发育良好的假种皮,同时他还强调Corner与Esser相互矛盾的观察结果仍然需要继续的研究才能确定。他指出详细的个体发育研究是解决这个矛盾所必须的。Tokuoka[5]报道了澳洲杨属植物的种子有肉质化的假种皮(或种阜),这样的报道使得区别种阜和假种皮更加混乱。Boesewinkel指出为了能理解种子的结构,关于胚珠发育的知识是关键[6],然而这些信息通常通过干燥的材料来获得是非常困难的[4]。
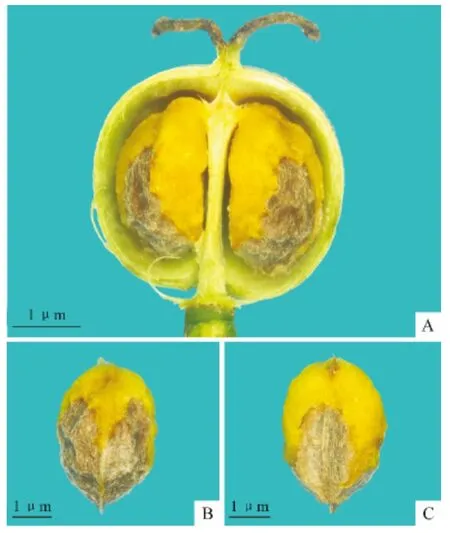
图1 澳洲杨的成熟果实和种子(黄色肉质化结构为种子附属结构)Fig.1 Mature fruit and seeds of H.populifolius Graham(showing the fleshy seed appendage)
澳洲杨或血色杨,属大戟科植物,灌木或小乔木,分布于澳大利亚的东北和北部,所罗门群岛和新圭亚那[3]。其生长速度快,能够被用作热带雨林恢复的先锋树种。在澳大利亚的植被景观恢复工程中常被用作先锋树种[7]。本文通过研究澳洲杨从胚珠到种子成熟的整个发育过程来揭示本属植物的种子附属物结构,同时通过研究鸭脚西番莲(Pasiflora citrinaJ.M.MacDougal)的假种皮结构,并与澳洲杨的肉质化结构进行比较,以明确种子附属物的不同发育模式及其生物学功能。
1 材料和方法
1.1 材 料
本实验材料是澳洲杨和鸭脚西番莲的不同发育时期花芽、花序、花及果实和种子;采样时间为2011年的1月份到2012年的1月份,花芽、果实和种子时期每周取样1次,花期每周取样2次,每次取样3~5个样本;材料采自于德国波鸿鲁尔大学植物园。
1.2 方 法
采集的新鲜材料立即用FAA 固定液固定,之后到实验室抽真空最少30min,使固定液完全浸入组织,然后室温保存2d。之后用70%的酒精替换FAA 固定液进行永久保存,在电镜扫描之前,利用光学解剖镜将胚珠从花中解剖出来;通过FDA(formaldeyde demethyl acetal)经 过4℃冰箱下24 h脱水;利用BALZERS CPD 030进行临界点干燥;将干燥的样品通过导电胶,粘贴到铝制样台上;在42~43 mA 强度的电流下喷金200s;然后利用ZEISS DSM 950进行扫描电镜观察。所有的图片经过“Digital Image System 2.2”进行储存,像素设定为2 000×2 000,以防原始图片的信息有所损失。原始图片经过Adobe Photoshop?整理为图版,模式图利用CorelDRAW X4绘制。
2 结果与分析
2.1 澳洲杨的胚珠及种阜发育
澳洲杨的珠心椭球形(图版Ⅰ,A)形成一个喙状结构,延伸到内珠被外(图版Ⅰ,A)。外珠被在内珠被的基部外沿发育(图版Ⅰ,B)。随着外珠被的生长,胚珠渐渐向腹面弯曲。接着珠心纵向膨大,形成为一个舌状结构在两层珠被外(图版Ⅰ,C),最终珠心顶端碰到心皮内面才被挤压而停止向外生长(图版Ⅰ,D)。之后两层珠被快速地生长,速度快过珠心将其包裹。外珠被的生长速度又快过内珠被,如此胚珠形成外珠孔类型。外珠被在珠心和内珠被外形成一个“连衣帽”结构(图版Ⅰ,D~F)。最终外珠被将整个胚珠包裹起来只留下一个缝隙——珠孔(图版Ⅱ,A)。在外珠孔形成后,外珠被的边缘增厚,增厚的组织逐渐扩展到珠柄,形成一个围绕珠孔和珠柄的环(图版Ⅱ,B、C)——种阜。随着种阜的生长,其慢慢向下包裹部分种子(图版Ⅱ,D)。这个结构在种子成熟时会发育成一个肉质化结构并且包裹一半的种子(图版Ⅱ,D~F)。

图2 鸭脚西番莲假种皮发育Fig.2 P.citrinaJ.M.MacDougal:developmental stages of aril
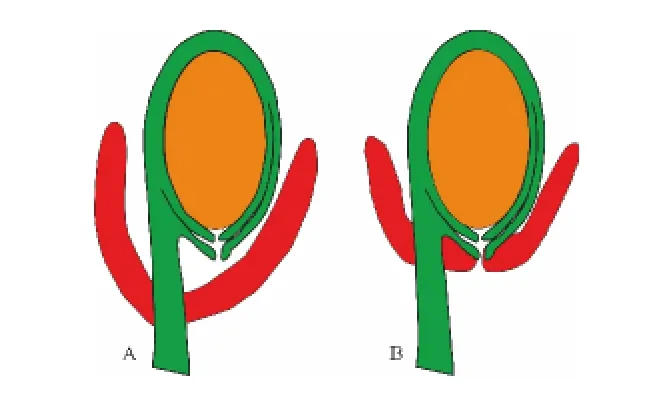
图3 假种皮与种阜结构模型Fig.3 Two models of aril and caruncula
2.2 鸭脚西番莲的假种皮发育
鸭脚西番莲的珠孔由内珠被和腹侧外珠被构成,为内珠孔类型(图2,A)。其肉质结构起源于珠柄,且开始就是珠柄上的一个环形结构。其发育在花粉管进入胚珠之后,随着发育而将珠孔覆盖(图2,B)。研究发现不止一个花粉管同时进入珠孔(图2,B),大戟科假种皮与种阜结构模型见图3。
3 讨 论
Gaertner[8]是第一个使用假种皮这个概念来描述种子的附属结构[1,9]。之后不同的研究者使用各自的概念来命名同样的结构,或者将不同的结构置于同一个概念下[10-15]。如此一来,对于种子附属结构的认识非常混乱,特别是对于一些种,因为材料有限而描述不完整或者相互矛盾的情况时有发生。直到Kapil等[1]基于生长模式与起源位置的不同将各种不同种子附属结构进行了分类。
澳洲杨属的种子附属结构曾被记载为泡状外种皮(似假种皮)[3],发育良好的假种皮[4]和肉质化假种皮(种阜)[5]。然而,以上研究者都没进行个体发育研究,多是通过干燥蜡叶标本进行推断性描述。本研究结果可以清楚显示,澳洲杨种子的附属结构从外珠孔发育而来,所以可以明确定义为种阜而非其他结构。这个结果部分支持了Tokuoka的观察,但同时也反映出Tokuoka没有清楚地将假种皮与种阜区别开来。典型假种皮是一个由珠柄发育而来的环状肉质结构,其发育会将珠孔与胚珠包裹起来,所以不能在假种皮上发现珠孔。而种阜是一个小的、碟形的结构,其起源为外珠孔的环形附属物,在种阜中心可以识别出珠孔。相对于是否起源于外珠孔及能否识别出珠孔,大小和形状都不是诊断种阜的关键特征[16]。
关于假种皮与种阜的区别,本文作者在有关卫矛科植物肉质化种子附属结构的文章中有过详细讨论,并指出了卫矛科植物具有种阜而非假种皮[17]。Kloos指出西番莲属植物的假种皮起源于配子体时期,且全部是表皮细胞发育而来[18]。然而本文的结果显示,假种皮的发生是在花粉管进入珠心之后。从功能考虑,如果假种皮起源于配子体时期,其过早的发育很可能会阻碍花粉管进入珠心的唯一通道——珠孔,因为假种皮的发育会覆盖珠孔。或者假种皮的发育本身就是需要受精作用来启动,通过覆盖珠孔来阻挡其他花粉管继续进入珠心。然而种阜并没有关闭珠孔的功能,其发生是否早于受精,要解答这些问题还需要更进一步的详细研究。
致谢:感谢德国波鸿鲁尔大学植物园为本研究提供实验材料;感谢英国皇家植物园的Simon Mayo教授对于本文撰写过程中提供的宝贵建议;感谢波鸿鲁尔大学Sabine Adler女士在参考文献查阅与翻译上的帮助;感谢国家留学基金委对本研究的支持。

图版Ⅰ 澳洲杨胚珠的连续发育过程PlateⅠ The successive developmental stages of the ovule in H.populifolius Graham

图版Ⅱ 澳洲杨种阜的连续发育过程PlateⅡ The successive developmental stages of the caruncula in H.populifolius Graham
[1]KAPIL R N,BOR J,BOUMAN F.Seed appendages in angiosperms[J].BotanischeJahrbücherfürSystematik,Pflanzengeschichteund Pflanzengeographie,1980,101:555-573.
[2]CORNER EJH.The Seeds of Dicotyledons(Vols.1and 2)[M].Cambridge:Cambridge University,1976.
[3]ESSER H J.A revision of omalanthus(Euphorbiaceae)in Malesia[J].Blumea,1997,42:421-466.
[4]GARDNER RHO.Notes on the fruit and seed ofHomalanthus(Euphorbiaceae)[J].Adansonia,1999,21(2):301-305.
[5]TOKUOKA T,TOBE H.Ovule and seed in Euphorbiodeae(Euphorbiaceae):structure and systematic implications[J].JournalofPlant Research,2002,115:361-374.
[6]BOESEWINKEL E D,BOUMAN E.The seed:structure[M]//JOHRI B M.Embryology of Angiosperms.Berlin:Springer-Verlag,1984:567-610.
[7]FLOYD A G.Rainforest Trees of Mainland South-Eastern Australia(Second Edition)[M].Terania:Rainforest Publishing,b&w illustrations,2008.
[8]GAERTNER J.De Fructibus et Seminibus Plantarum[M].Stuttgart,1788.
[9]RODRIGUEZ-RIAN O,FRANCISCO J,VALTUEN A,etal.Megasporogenesis,megagametogenesis and ontogeny of the aril inCytisus striatusandC.multiflorus(Leguminosae:Papilionoideae)[J].AnnalsofBotany,2006,98:777-791.
[10]PLANCHON M J E.Développements et caractères des vrais et des faux arilles[J].AnnalesdesSciencesNaturelles;Botanique,1845,3:275-312.
[11]BAILLON H.Sur I′origine de I′arille des Carumbium[J].Adasonia,1867,6:348-351.
[12]PFEIFFER A.Die Arillargebilde der Pflanzensamen[J].BotanischeJahrbücherfürSystematik,PflanzengeschichteundPflanzengeographie,1891,13:492-540.
[13]CORNER E J H.The durian theory or the origin of the modern tree[J].AnnalsofBotany,1949,13(4):367-414.
[14]CORNER E J H.The durian theory extended I[J].Phytomorphology,1953,3:465-476.
[15]VAN DER P I J L L.Principles of dispersal in higher plants[M].Springer,Berlin,Heidelberg,New York,1972.
[16]BAILLON H.Etude générale du groupe des Euphorbiacées[M].Paris,1858.
[17]ZHANG X,ZHANG Z,STÜTZEL T.Aril development in Celastraceae[J].FeddesRepertorium,2012,122:445-455.
[18]KLOOS A,BOUMAN F.Case studies in aril developmentPassiflorasuberosaandTurneraulmifolia[J].BeitraegeZurBiologieDer Pflanzen,1980:49-66.
