蜡蚧菌的遗传多样性及其对柑橘木虱的致病性
2015-06-05鹿连明程保平杜丹超胡秀荣蒲占湑陈国庆
鹿连明,程保平,杜丹超,胡秀荣,蒲占湑,陈国庆*
(1.浙江省柑桔研究所,浙江台州318026;2.广东省农业科学院植物保护研究所,广州510640)
蜡蚧菌的遗传多样性及其对柑橘木虱的致病性
鹿连明1,程保平2,杜丹超1,胡秀荣1,蒲占湑1,陈国庆1*
(1.浙江省柑桔研究所,浙江台州318026;2.广东省农业科学院植物保护研究所,广州510640)
为探究蜡蚧菌(Lecanicillium lecanii)的遗传多样性、对柑橘木虱的致病性及其影响因素,对19个不同寄主种类和地理来源的蜡蚧菌菌株进行随机引物扩增多态性DNA(randomly amplified polymorphic DNA,RAPD)分析,并对其ITS1-5.8S rDNA-ITS2和β-tubulin基因进行测序分析及构建系统发育树,同时测定菌株的生长速度、产孢量、孢子萌发率等生物学特性及对柑橘木虱的致病性.结果显示:蜡蚧菌种内存在丰富的遗传多样性,不同菌株的生长速度、产孢量和孢子萌发率等生物学特性及对柑橘木虱致病性均表现出明显的差异,但未发现彼此之间及其与菌株的寄主种类和地理来源之间具有相关性.在显微镜下观察菌株ZJVL-A对柑橘木虱的侵染过程发现,吸附在寄主体表的分生孢子可在接种后16 h萌发入侵,42 h后菌丝侵入血腔并在其中增殖,54 h后可引起寄主死亡,60 h后菌丝侵染并破坏寄主的内部组织器官,最后覆盖在虫体表面的菌丝又产生大量的分生孢子.本研究为进一步了解蜡蚧菌对柑橘木虱的致病机制,为开发用于柑橘木虱田间防治的生物制剂提供了依据.
蜡蚧菌;遗传多样性;柑橘木虱;致病性
SummaryDiaphorina citri may be the most serious pest of citrus worldwide,primarily because it is the vector of huanglongbing(HLB),one of the most destructive citrus diseases,and can damage citrus directly by feeding fresh shoots and cause citrus sooty mold disease.Currently,control of D.citri mainly depends on the chemical pesticides.However,pesticides abuse affects workers and food safety,causes the development of insecticideresistant D.citri populations,and reduces populations of natural enemies in citrus orchards.Therefore,new approaches are needed to complement the existing management strategies against D.citri.
Lecanicillium lecanii,formerly named Verticillium lecanii,is an entomopathogenic fungus with a remarkably wide host range,which can infect aphids,whiteflies,trips,plant hoppers and so on.Infection of L.lecanii against D.citri was firstly reported in 1980s.However,from then on,there is no further research such as biologicalcharacteristic and pathogenicity difference about this fungus.Therefore,in this study,genetic diversities of 19 L.lecanii strains from different hosts and geographical origins as well as their biological characteristics and pathogenicities against D.citri were determined and analyzed.This research aimed to understand the genetic diversity,pathogenicity difference and their relationships with geographical origins,host varieties and biological characteristics.
Firstly,19 L.lecanii strains from different hosts and geographical origins were characterized using randomly amplified polymorphic DNA(RAPD)analysis.Based on the RAPD results,a dendrogram was constructed using unweighted pair group method with arithmetic mean(UPGMA)by software NTSYS-pc version 2.1.Then,the ITS1-5.8S r DNA-ITS2 andβ-tubulin genes of these L.lecanii strains were amplified and sequenced using primer sets ITS5/ITS4 and Bt2a/Bt2b,respectively.These sequences were aligned with software Clustal X 2.0,edited with software BioEdit 7.2.5,and finally a phylogenic tree was constructed using the neighbor-joining method by software MEGA 5.05.Besides,the biological characteristics of these strains such as mycelial growth rates,sporulation quantities,conidial germination rates and conidial sizes were also determined.Furthermore,the pathogenicities of these strains against D.citri were assayed,and the invasion process of strain ZJVL-A was observed under a dissecting microscope,a scanning electron microscope and a light microscope.
The results showed that a total of 96 polymorphic bands were amplified using 7 random primers,and the size of the bands ranged f rom 0.2-3 kb.Al l the 19 L.lecanii st rains were divided into 2 groups in the UPGMA dendrogram.Strain CGVL-11 was located alone in one group,while the other strains were clustered into another group.In the latter group,18 strains were then divided into 2 subgroups.The ITS1-5.8S r DNA-ITS2 andβ-tubulin gene sequences had been deposited in GenBank and their accession numbers were KJ598810-KJ598828 and KJ598829-KJ598847,respectively.The similarities of ITS1-5.8S r DNA-ITS2 andβ-tubulin gene sequences were 98%-100%and 96%-100%with each other,and molecular variations existed within these nucleotide sequences. The phylogenetic tree based on ITS1-5.8S rDNA-ITS2 andβ-tubulin sequences was consistent with the dendrogram generated through RAPD analysis.The mycelial growth rates,sporulation quantities,conidial germination rates and conidial sizes of these strains were remarkably dif ferent.The colony diameter was 4.25-2.85 cm after 10 d incubation,and the sporulation quantity was 0.10×108-9.75×108conidia/m L after 3 d incubation in PDB medium,and the conidial germination rate was 3.55%-68.38% after 8 h incubation at 25℃,and the size of conidia was 4.0-5.0 μm in length and 1.4-1.7μm in width.Conidia of L.lecanii strain ZJVL-A mainly distributed in the intersegmental fold,setal alveolus,anus,genital,and other fold or sunken regions on every tagmata.At 16 h post-inoculation,the conidia began to germinate and penetrated the cuticle of host.At 42 h post-inoculation,the hyphae invaded into haemocoel,proliferated abundantly and filled the entire haemocoel.Diaphorina citri was killed by the fungus after 54 h postinoculation.After 60 h post-inoculation,the internal organs of host such as fat body,muscle tissue,digestive tract and ovary were invaded and progressively degraded.Finally,a large number of newly-developed conidia could be observed on the hyphae which covered the body surface of host.
In conclusion,genetic diversities and pathogenicity differences exist in populations of L.lecanii.Moreover,their biological characteristics also show significant differences.However,they have no relationship with each other and with host varieties or geographical origins.The strain ZJVL-A maybe kill D.citri through secreting some toxins before the hyphae degrade the internal organs of host.This research lays a good foundation for further study on the pathogenic mechanism of L.lecanii against D.citri,and takes an important step towards developing and utilizing these entomopathogenic fungi for their potential to suppress D.citri populations.
柑橘木虱(Diaphorina citri)是柑橘生产上的一种重要害虫,除作为唯一自然虫媒传播柑橘上的毁灭性病害——柑橘黄龙病外,还可直接取食危害新梢嫩叶及产生分泌物诱发煤烟病等.目前对柑橘黄龙病尚缺乏有效的防治药剂和理想的抗病品种,因此加强对柑橘木虱的防治,无论对于控制柑橘黄龙病的流行还是降低虫体本身对柑橘的危害都具有重要的意义.目前对于柑橘木虱的防治多采用化学防治措施,然而化学农药的频繁使用造成了农药残留、环境污染、生物多样性破坏和柑橘木虱产生抗药性等诸多问题,而生物农药以其高效、低毒、低残留、无污染、不易产生抗药性等特性已逐渐引起人们的重视.已有报道[1-5]表明,对柑橘木虱具有致病活性的真菌有檬形被毛孢(Hirsutella citriformis)、宛氏拟青霉(Paecilomyces varioti)、玫烟色拟青霉(P. fumosoroseus)、球孢白僵菌(Beauveria bassiana)、蚜笋顶孢霉(Acrostalagmus aphidium)、黄色镰刀菌(Fusarium culmorum)等.但目前对于柑橘木虱生防菌的研究多处于实验室阶段,在农业生产中利用生防菌制剂成功防治柑橘木虱的实例尚未见报道.
蜡蚧菌(Lecanicillium lecanii)原命名为蜡蚧轮枝菌(Verticillium lecanii),是一种寄主范围极为广泛的昆虫病原真菌,可寄生介壳虫、蚜虫、温室白粉虱、飞虱、蓟马等多种昆虫.蜡蚧菌具有侵染柑橘木虱的活性最早由我国谢佩华等[6]于20世纪80年代发现,但之后国内外学者对该菌的生物学特性和对柑橘木虱的致病活性均未再进行深入研究.蜡蚧菌或因地理来源和寄主品种的不同,种内表现出丰富的遗传多样性,且不同菌株对相同的寄主表现出致病性分化.而关于蜡蚧菌不同菌株是否对柑橘木虱存在致病性差异,以及影响菌株致病性的因素,目前尚未见相关的研究报道.基于此,本研究对不同寄主种类和地理来源的蜡蚧菌菌株的遗传多样性进行分析,以探讨蜡蚧菌不同菌株之间的亲缘关系及其与寄主种类和地理来源之间的关系.同时测定菌株对柑橘木虱的致病活性,了解不同菌株之间的致病性差异及影响因素,为今后筛选或改造用于防治柑橘木虱的优良菌株提供依据.
1 材料与方法
1.1 材料
1.1.1 供试菌株 蜡蚧菌CIVL-1和CIVL-2购自中国工业微生物菌种保藏管理中心,ACVL-3、ACVL-4、ACVL-5、ACVL-6、ACVL-7、ACVL-8、ACVL-9和ACVL-10购自中国农业微生物菌种保藏管理中心,CGVL-11、CGVL-12和CGVL-13购自中国普通微生物菌种保藏管理中心,AHVL-14、AHVL-15、AHVL-16、AHVL-17和AHVL-18购自安徽农业大学,ZJVL-A为本实验室从浙江台州地区橘园内的柑橘木虱上分离纯化获得.以上各菌株的分离基物和地理来源详见表1.
1.1.2 柑橘木虱 健康柑橘木虱采集于浙江省台州市黄岩区橘园内,并在温室条件下人工饲养于九里香植株上.
1.1.3 主要试剂 DNA提取试剂如十六烷基三甲基溴化铵(CTAB)、酚/三氯甲烷/异戊醇、三氯甲烷/异戊醇、异丙醇、聚乙烯基吡咯烷酮(PVP)、β-巯基乙醇等为Amresco公司产品,PCR试剂如Taq酶、d NTP等购自大连宝生物公司(Ta KaRa),琼脂糖为Biowest公司产品,凝胶染料为Biotium公司生产的Gel Red,DNA凝胶回收试剂盒为Axygen公司产品,六胺银(GMS)染料为Merck公司产品,其余试剂为国产分析纯.
1.1.4 引物 通过预实验反复筛选,从60条随机引物中获得扩增多态性高、扩增条带清晰、重复效果好的7条随机引物对蜡蚧菌进行随机扩增多态性DNA(randomly amplified polymorphic DNA,RAPD)分析,所用引物名称和序列分别为:A04(5′-ATCA GCGCACCA-3′)、OPA02(5′-TGCCGAGCTG-3′)、OPH13(5′-GACGCCACAC-3′)、OPB06(5′-TGCTC TGCCC-3′)、OPD18(5′-GAGAGCCAAC-3′)、OPB12(5′-CCTTGACGCA-3′)、M13(5′-GAGGGTGGCGG TTCT-3′).用于扩增蜡蚧菌ITS1-5.8S r DNA-ITS2基因所用引物为真菌通用引物ITS5(5′-GGAAGT AAAAGTCGTAACAAGG-3′)和ITS4(5′-TCCTCCG CTTATTGATATGC-3′),扩增β-tubulin基因所用引物为真菌通用引物Bt2a(5′-GGTAACCAAAT CGGTGCTGCTTTC-3′)和Bt2b(5′-ACCCTCAGT GTAGTGACCCTTGGC-3′).以上引物均由上海Invitrogen公司合成.
1.2 方法
1.2.1 样品总DNA提取 将各菌株分别接种至PDA培养基平板上,置于25℃恒温培养10 d左右,以无菌药匙刮取PDA平板上生长的真菌菌丝于无菌研钵中,加入液氮研磨至粉末状,采用改良的CTAB法[7]提取菌株总DNA.最后将所提取的DNA沉淀溶于10~20μL TE[含核糖核酸酶(Ribonuclease,RNase)50 mg/L]中即得样品DNA溶液.
1.2.2 RAPD分析 25μL的RAPD反应体系:5 ng真菌基因组DNA,1×PCR缓冲液,1.5 mmol/L MgCl2,0.2 mmol/L d NTP,0.6μmol/L引物,1 U Taq酶.反应条件:94℃预变性5 min,然后94℃1 min,37℃1.5 min,72℃2 min,共45个循环,最后72℃延伸10 min.扩增产物上样于GelRed染料预染的2%琼脂糖凝胶电泳,并于凝胶成像系统中观察拍照.

表1 蜡蚧菌各菌株的分离基物、地理来源和生物学特性Table 1 Isolation substrates,geographical origins and biological characteristics of L.lecanii strains
对每一引物RAPD扩增带进行样品间比较,将每条RAPD条带均看作一个遗传位点,有带记为1,无带记为0,构建二元数据矩阵,应用UPGMA进行聚类分析,以NTSYS-pc 2.1运算建立聚类图.
1.2.3 基因序列分析 分别以真菌通用引物ITS5/ITS4和Bt2a/Bt2b扩增蜡蚧菌ITS1-5.8S rDNA-ITS2和β-tubulin基因,PCR反应体系和反应条件参考鹿连明等[8]的报道.PCR反应结束后上样于2%琼脂糖凝胶电泳,并用Axygen公司的DNA凝胶回收试剂盒回收PCR产物.将纯化后的PCR产物送交Invitrogen公司,利用上述引物直接测序.对所测序列整理后,利用Clustal X 2.0软件进行多重序列比对,通过BioEdit 7.2.5软件进行编辑,最后利用软件MEGA 5.05的邻接法联合基因ITS1-5.8S r DNAITS2和β-tubulin,以L.longisporum作为外群,构建系统发育树.
1.2.4 生物学特性测定 在PDA培养基上培养7 d左右的各菌株菌落周围,以无菌打孔器打取5 mm大小的菌饼,然后将菌饼分别置于数个新鲜制备的PDA平板中央,在25℃微生物培养箱中倒置暗培养.连续培养14 d,每24 h观察菌落生长情况并采用十字交叉法测量菌落直径.每个处理设3个重复.
将直径为9 mm的菌饼接种至150 m L PDB培养液中,置于25℃恒温摇床中以转速200 r/min振荡培养3 d.取各菌株的培养液适当稀释后滴加至血球计数板中,在光学显微镜下观察并记录孢子数量,然后计算各培养液的孢子浓度.同时,利用显微镜成像软件测量并记录孢子大小.每个处理设3个重复.
分别取30μL上述培养的各菌株的分生孢子悬浮液滴加至无菌载玻片的中央,然后将玻片放于垫有滤纸的培养皿中的U型管上,以无菌水湿润滤纸,然后置于25℃培养箱中暗培养.培养8 h后在光学显微镜下计数400~600个孢子,以芽管长度超过孢子长度1/2为标准视为孢子萌发,统计孢子萌发个数并计算孢子萌发率.每个处理设3个重复.
1.2.2 致病性测定 以无菌水调整各菌株的分生孢子悬浮液使其为1.0×108个孢子/m L,并向其中加入适量的吐温-80至终体积分数为0.1%.参考鹿连明等[8]的方法用各分生孢子悬浮液处理柑橘木虱,同时设立以含0.1%吐温-80的无菌水处理柑橘木虱为对照.之后放于25℃人工气候箱中每天14 h光照/10 h黑暗培养,以上每个处理设3个重复,每个重复处理30头柑橘木虱.每天观察病菌对柑橘木虱的侵染和致死情况,记录柑橘木虱病死数目,计算病死率和校正病死率.
在培养后不同时间对菌株ZJVL-A处理的柑橘木虱取样,在体视显微镜下直接观察柑橘木虱体表的菌丝生长情况;将柑橘木虱样品用2.5%戊二醛溶液固定,然后经清洗、干燥、喷金等处理[9]后,在扫描电子显微镜下观察菌株ZJVL-A在柑橘木虱体表的侵染情况;将柑橘木虱样品先于4%的多聚甲醛溶液中固定,再对其进行石蜡包埋和切片,然后将超薄切片放入苏木精-伊红(HE)染色液或六胺银(GMS)染色液中染色处理[10]后,置于光学显微镜下观察菌株ZJVL-A在柑橘木虱体内的侵染情况.
1.2.6 数据统计分析 将蜡蚧菌各菌株的孢子大小、菌落大小、产孢量、孢子萌发率和对柑橘木虱的校正病死率等数据在Microsoft Excel 2003中进行整理,利用SAS 9.1软件的邓肯新复极差法(DMRT)进行差异显著性分析.
2 结果与分析
2.1 蜡蚧菌的分子特征
利用7条随机引物共从19个蜡蚧菌菌株中扩增获得104个条带,扩增条带大小分布在0.2~3 kb范围内,其中多态性条带为96个,多态性比率为92.31%,表明供试菌株群体具有较丰富的遗传多样性.引物A-04和OPA-02扩增产物的电泳结果如图1所示.
从供试菌株全基因组扩增DNA片段的系统树状聚类图(图2)可以看出,供试的19个菌株表现为遗传多样性,遗传相似系数在0.38~1.0之间.以遗传相似系数0.38为阈值,可将所有供试蜡蚧菌菌株划分为2大遗传聚类组Ⅰ和Ⅱ.其中Ⅱ组中仅含有CGVL-11菌株,而Ⅰ组则包括CGVL-11之外的所有18个菌株.在遗传相似系数0.73水平上,Ⅰ组的18个菌株又被划分到2个亚组中.其中所有来自中国的8个蜡蚧菌菌株都聚类到亚组ⅰ中,另外该亚组还包括2个来自古巴的菌株和2个未知地理来源的菌株.而亚组ⅱ则包含有来自欧洲的3个菌株和来自美洲的3个菌株.
以引物ITS5/ITS4扩增蜡蚧菌各菌株的基因大小为523 bp左右,其序列已在GenBank中登录,登录号按表1从上到下菌株的顺序依次为KJ598810~KJ598828.以引物Bt2a/Bt2b扩增蜡蚧菌各菌株的基因片段大小为357bp左右,在GenBank中的登录号依次为KJ598829~KJ598847.多重序列比对结果显示,蜡蚧菌各菌株的ITS1-5.8S r DNA-ITS2序列相似性为98%~100%,β-tubulin基因片段的序列相似性为96%~100%,不同菌株间基因序列或完全一致,或存在一定的碱基差异,其中尤以菌株CGVL-11与其他各菌株的序列差异最大.联合蜡蚧菌ITS1-5.8S r DNA-ITS2和β-tubulin 2个基因构建系统发育树(图3),从中可以看出,基于各菌株基因序列的聚类结果与基于RAPD的聚类结果基本一致.

图1 引物A-04和OPA-02扩增蜡蚧菌基因组DNA的RAPD图谱Fig.1 RAPD fingerprints of L.lecanii genomic DNA amplified by primers A-04 and OPA-02
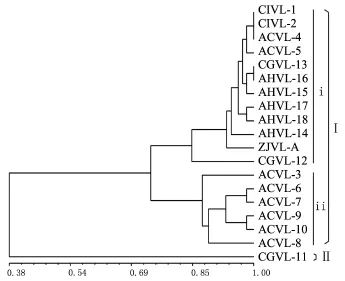
图2 基于RAPD的扩增结果通过UPGMA算法构建的蜡蚧菌系统树状聚类图Fig.2 Dendrogram of L.lecanii derived from UPGMA cluster analysis based on RAPD-PCR
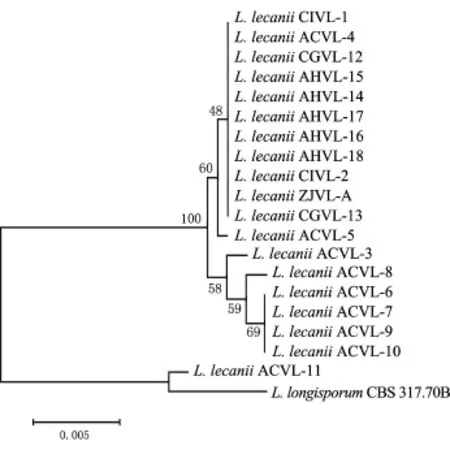
图3 基于ITS1-2.8S rDNA-ITS2和β-tubulin基因构建的蜡蚧菌的系统发育树Fig.3 Phylogenetic tree of L.lecanii constructed based on ITS1-5.8S r DNA-ITS2 andβ-tubulin genes
2.2 蜡蚧菌的生物学特性
蜡蚧菌各菌株的菌落生长速度存在明显差异.在25℃下暗培养10 d后各菌株的菌落直径在4.25~2.85 cm不等(表1),其中菌株AHVL-14生长速度最快,菌落直径可达4.25 cm;而菌株ACVL-4生长速度最慢,其菌落直径仅为2.85 cm.
于25℃恒温摇床中以转速200 r/min振荡培养3 d后,蜡蚧菌各菌株的产孢量详见表1.从中可以看出,蜡蚧菌不同菌株在相同条件下产孢量大小亦明显不同.其中以菌株ACVL-3的产孢量最大,为9.75×108个/m L;而菌株CGVL-11的产孢量最小,仅为0.10×108个/m L.
在光学显微镜下观察菌株孢子大小,结果(表1)表明,蜡蚧菌各菌株的孢子大小不等,但多数菌株的孢子长度在4.0~5.0μm范围内,宽度在1.4~1.7μm范围内.菌株CGVL-11孢子最大,长度约为7.52μm,宽度约为2.35μm;而菌株ACVL-5的孢子长度最小,约为3.49μm;菌株ACVL-9和AHVL-15的孢子宽度最小,约为1.38μm.
在25℃培养箱中暗培养8 h后,蜡蚧菌各菌株的分生孢子萌发率详见表1.从中可以看出,不同菌株的孢子萌发率存在较大差异.其中菌株CIVL-2的萌发率最高,约为78.44%;而菌株ACVL-6的孢子萌发率最低,仅为3.55%.
2.3 蜡蚧菌的致病性
在实验室条件下,蜡蚧菌不同菌株对柑橘木虱的致病性表现出明显差异.在1.0×108个/m L分生孢子悬浮液接种后第6天,各菌株对柑橘木虱的校正病死率在70%~90%之间(表1).其中以菌株ZJVL-A、AHVL-18和AHVL-16对柑橘木虱的致死率为最高,其校正病死率分别为89.84%、88.91%和88.73%;而菌株CGVL-11对柑橘木虱的校正病死率仅为70.13%.
以菌株ZJVL-A的分生孢子悬浮液接种柑橘木虱后30 h,在体视显微镜下可观察到柑橘木虱腹部末端的肛门、生殖器等部位有白色菌丝长出,此时柑橘木虱仍保持较正常的活动能力;接种后42 h,虫体的头部、胸部背面、翅下和腹部背面等部位均可观察到菌丝(图4A),此时柑橘木虱表现为反应迟钝、行动迟缓;之后,菌丝在虫体上持续生长,至54 h柑橘木虱出现死亡;接种后108 h,可见大量的白色菌丝将柑橘木虱虫体完全覆盖(图4B).
在扫描电子显微镜下可观察到柑橘木虱头部的触角窝、触角基节、复眼与触角间隙处等部位,胸部的翅腋区、胸足基节窝,腹部的肛门、产卵器或交配器等褶皱凹陷部位均分布有较多孢子.在接种后16 h,可观察到柑橘木虱体表的分生孢子萌发形成芽管,有些芽管伸长后向体壁定向生长直接穿透体壁,有的则在芽管末端形成附着胞吸附在体壁上再侵入,有的芽管生长形成菌丝沿体表平行生长,寻找到合适部位后再侵入;在接种后30 h,可观察到菌丝在柑橘木虱体表纵向或横向分支生长(图4C);之后,菌丝在虫体表面继续迅速生长,菌丝相互缠绕交织,形成菌丝体,最后又新形成大量的分生孢子(图4D).
取菌株ZJVL-A接种后培养不同时间的柑橘木虱样品,经石蜡包埋切片后于HE或GMS溶液中染色,然后置于光学显微镜下观察.可以发现在接种后16 h,在寄主体表的分生孢子萌发产生的菌丝穿透表皮层或在表皮层横向生长;之后,菌丝侵染寄主的皮细胞层,并在其中不断生长和繁殖;接种后42 h,菌丝侵入寄主血腔,并在其中不断增殖至菌丝充满整个血腔(图4E);接种后60 h,可观察到血腔内的菌丝侵染寄主的脂肪体、肌肉组织、消化道、卵巢等组织器官(图4F);之后,寄主各组织器官逐步被破坏降解,菌丝相互粘连缠绕.
3 讨论与结论
已有研究结果表明,蜡蚧菌种内存在着丰富的遗传多样性,但对于遗传多样性的影响因素目前尚没有确定的结论.Sugimoto等[11]通过PCR-RFLP和PCR-SSCP对46个不同寄主和地理来源的蜡蚧菌分离物进行了分析,结果发现不同菌株间存在遗传多样性,但其与寄主和地理来源无相关性;Mor等[12]和Mitina等[13]分别对不同蜡蚧菌分离物进行研究也均未发现菌株的聚类与寄主或地理来源有相关性;而邱君志等[14]分析认为蜡蚧菌菌株间的遗传多样性与地理来源有关.本研究结果表明蜡蚧菌种内存在遗传多样性,但其与菌株的寄主种类无相关性,尽管所有来自中国的8个分离物聚为一组,但该组同时也包括了来自古巴的2个菌株,因此也不足以表明与菌株的地理来源具有相关性.
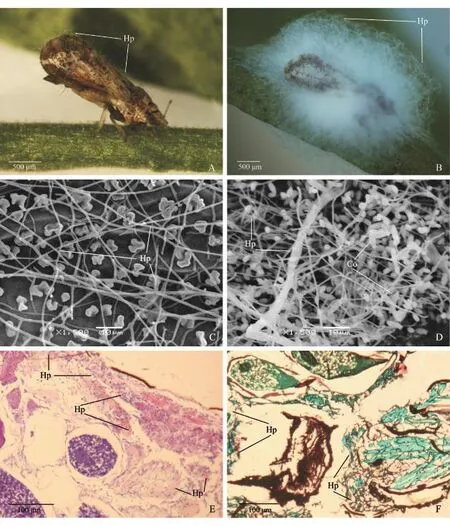
图4 在显微镜下观察菌株ZJVL-A对柑橘木虱的侵染过程Fig.4 Invasion process of strain ZJVL-A against D.citri observed under microscopes
蜡蚧菌不同菌株对相同的寄主表现出明显的致病力差异,但关于菌株的致病力强弱与菌株的寄主种类、地理来源和生物学特性等之间的关系,不同的研究结果不尽相同.Sugimoto等[11]和Mitina等[13]分别研究发现蜡蚧菌的致病性强弱与菌株的地理来源和生物学特性无明显的相关性,但从与靶标昆虫种类相同的寄主上分离的菌株往往具有更强的致病性;Mor等[12]和Fatiha等[15]的研究结果表明蜡蚧菌的致病性强弱与菌株的RAPD多态性、寄主种类和地理来源均未表现出相关性,但生长速率快、产孢量大的菌株对烟粉虱的致病力更高;Hall[16]发现蜡蚧菌对粉虱和蚜虫的致病力强弱与菌株的产孢能力、孢子的大小和萌发率具有相关性,且分生孢子大的菌株具有更高的病害流行潜能;Jackson等[17]发现蜡蚧菌的致病力与菌株的产孢量和孢子萌发率有关,但与孢子大小无关;Mitina等[13]也发现蜡蚧菌对白粉虱的致病力与孢子大小之间的相关性很低.本研究发现对柑橘木虱致病力最强的3个菌株均来自于中国,但不能据此认为蜡蚧菌的致病力与地理来源相关,因为来自于古巴的2个菌株较中国的其他菌株具有更强的致病力.除菌株ACVL-3外,在聚类树状图和系统发育树中与ZJVL-A聚为一组的菌株普遍较另外一组的菌株具有更强的致病力.因此,相比地理来源,蜡蚧菌对柑橘木虱的致病力或与菌株的亲缘关系具有更大的相关性,但这需要通过增加菌株的样本量进一步论证分析.另外,本研究也未发现蜡蚧菌的生长速度、产孢量、孢子萌发率、孢子大小和对柑橘木虱的致病性彼此之间具有相关性.因此,想获得一株同时具有产孢量大、孢子萌发率高、菌丝生长速率快和对寄主致病性强等优良性状的生防菌株往往较为困难.在进行优良菌株筛选时,可以致病性和产孢量为首要参考指标,其次再考虑孢子萌发率和菌丝生长速率.或者可通过紫外诱变、基因工程等手段对菌株改良,使其尽量具备以上各优良性状及更强的抗逆性等.
多数昆虫病原真菌侵染寄主通常包括分生孢子附着寄主表皮、分生孢子在昆虫表皮上萌发、穿透表皮、菌丝在血腔内生长、毒素产生、寄主死亡、菌丝侵入寄主的所有器官、菌丝穿出表皮、产生侵染单位和侵染单位扩散等几个阶段[18].本研究发现菌株ZJVL-A的分生孢子多分布在柑橘木虱的褶皱凹陷部位,而胸部孢子数量较少,分析其原因为木虱呈45℃爬行时,胸部与寄主植物发生摩擦导致其上的分生孢子脱落.在接种后16 h即可观察到寄主体表的孢子萌发入侵,60 h后血腔中的菌丝侵染并破坏昆虫的内部组织器官.值得注意的是,在菌丝完全侵染破坏寄主内部组织器官之前,接种后的柑橘木虱即已开始死亡,由此推测菌株ZJVL-A或许不是通过菌丝侵染破坏昆虫组织器官导致昆虫死亡,而是在菌丝侵入血腔后可能产生了某些毒素而加速了寄主的死亡,这也在一定程度上解释了以上所述的亲缘关系近的菌株可能具有更相近的致病力,但这需要进一步研究确定.
通过对不同寄主和地理来源的蜡蚧菌的遗传多样性、菌丝生长速度、产孢量、孢子大小和萌发率等生物学特性及对柑橘木虱的致病性的研究,我们更深入地了解了蜡蚧菌的遗传变异和致病性分化及其与菌株的寄主种类、地理来源和生物学特性之间的关系.而对蜡蚧菌对柑橘木虱的侵染过程的观察,则有助于我们进一步了解蜡蚧菌对柑橘木虱的致病机制.本研究对于我们今后筛选或改良对柑橘木虱具强致病力的病原微生物,开发用于柑橘木虱田间防治的生物制剂提供了依据.
(References):
[1] 杜丹超,鹿连明,张利平,等.柑橘木虱的防治技术研究进展.中国农学通报,2011,27(25):178-181.
Du D C,Lu L M,Zhang L P,et al.Research progress in control technology of Diaphorina citri.Chinese Agricultural Science Bulletin,2011,27(25):178-181.(in Chinese with English abstract)
[2] 陈祝安,曹光照,许益伟,等.柑桔害虫病原真菌资源的考察和生测.微生物学通报,1985,12(5):194-198.
Chen Z A,Cao G Z,Xu Y W,et al.Investigation and bioassay of pathogenic fungi resources of citrus pests. Microbiology China,1985,12(5):194-198.(in Chinese)
[3] Meyer J M,Hoy M A,Boucias D G.Isolation and characterization of an Isaria fumosorosea isolate infecting the Asian citrus psyllid in Florida.Journal of Invertebrate Pathology,2008,99(1):96-102.
[4] Meyer J M,Hoy M A,Boucias D G.Morphological and molecular characterization of a Hirsutella species infecting the Asian citrus psyllid,Diaphorina citri Kuwayama(Hemiptera:Psyllidae)in Florida.Journal of Invertebrate Pathology,2007,95(2):101-109.
[5] Aubert B.Trioza erytreae Del Guercio and Diaphorina citri Kuwayama(Homoptera:Psylloidea),the two vectors of citrus greening disease:biological aspects and possible control strategies.Fruits,1987,42(3):149-162.
[6] 谢佩华,苏朝安,林自国.柑桔木虱寄生菌——蜡蚧头孢菌初步研究.生物防治通报,1988,4(2):92.
Xie P H,Su C A,Lin Z G.A preliminary study on an entomogenous fungus of Diaphorina citri Kuwayama(Hom. Psyllidae).Chinese Journal of Biological Control,1988,4(2):92.(in Chinese with English abstract)
[7] 鹿连明,范国成,姚锦爱,等.柑橘黄龙病菌核糖体蛋白基因的多态性及系统发育分析.浙江大学学报:农业与生命科学版,2011,37(2):125-132.
Lu L M,Fan G C,Yao J A,et al.Polymorphism and phylogenetic analysis of ribosomal protein genes of Candidatus Liberibacter asiaticus.Journal of Zhejiang University:Agriculture&Life Sciences,2011,37(2):125-132.(in Chinese with English abstract)
[8] 鹿连明,杜丹超,胡秀荣,等.一株柑橘木虱虫生真菌的分离与鉴定.浙江农业科学,2013(10):1319-1322.Lu L M,Du D C,Hu X R,et al.Isolation and identification of an entomopathogenic fungi against Diaphorina citri. Journal of Zhejiang Agricultural Sciences,2013(10):1319-1322.(in Chinese)
[9] Leemon D M,Jonsson N N.Comparative studies on the invasion of cattle ticks(Rhipicephalus(Boophilus)microplus)and sheep blowflies(Lucilia cuprina)by Metarhizium anisopliae(Sorokin).Journal of Invertebrate Pathology,2012,109(2):248-259.
[10] Liu W M,Xie Y P,Xue J L,et al.Histopathological changes of Ceroplastes japonicus infected by Lecanicillium lecanii.Journal of Invertebrate Pathology,2009,101(2):96-105.
[11] Sugimoto M,Koike M,Hiyama N,et al.Genetic,morphological,and virulence characterization of the entomopathogenic fungus Verticillium lecanii.Journal of Invertebrate Pathology,2003,82(3):176-187.
[12] Mor H,Gindin G,Ben-Ze’Ev,et al.Diversity among isolates of Verticillium lecanii as expressed by DNA polymorphism and virulence towards Bemisia tabaci. Phytoparasitica,1996,24(2):111-118.
[13] Mitina G V,Mikhailova L A,Yli-Mattila T.RAPD-PCR,UP-PCR and r DNA sequence analyses of the entomopathogenic fungus Verticillium lecanii and its pathogenicity towards insects and phytopathogenic fungi. Archives of Phytopathology and Plant Protection,2008,41(2):113-128.
[14] 邱君志,黄天培,潘黎,等.蜡蚧轮枝菌种内遗传差异的研究.中国菌物学会第三届会员代表大会暨全国第六届菌物学学术讨论会论文集,2003:352-356. Qiu J Z,Huang T P,Pan L,et al.Intraspecific genetic diversity in Verticillium lecanii.Memoir of the 3rd Member Representative Assembly of Mycological Society of China and the 6th National Mycology Symposium,2003:352-356.(in Chinese)
[15] Fatiha L,Ali S,Ren S,et al.Biological characteristics and pathogenicity of Verticillium lecanii against Bemisia tabaci(Homoptera:Aleyrodidae)on eggplant.Pakistan Entomologist,2007,29(2):63-72.
[16] Hall R A.Epizootic potential for aphids of different isolates of the fungus Verticillium lecanii.Entomophaga,1984,29(3):311-321.
[17] Jackson C W,Heale J B,Hall R A.Traits associated with virulence to the aphid Macrosiphoniella sanborni in eighteen isolates of Verticillium lecanii.Annals of Applied Biology,1985,106(1):39-48.
[18] 蒲蛰龙,李增智.昆虫真菌学.合肥:安徽科学技术出版社,1996:94-95. Pu Z L,Li Z Z.Insect Mycology.Hefei:Anhui Science and Technology Press,1996:94-95.(in Chinese)
Genetic diversity of Lecanicillium lecanii and its pathogenicity against Diaphorina citri.Journal of
Zhej iang Universi ty(Agr ic.&Li fe Sci.),2015,41(1):34-43
Lu Lianming1,Cheng Baoping2,Du Danchao1,Hu Xiurong1,Pu Zhanxu1,Chen Guoqing1*(1.Citrus Research Institute of Zhejiang Province,Taizhou 318026,Zhejiang,China;2.Research Institute of Plant Protection, Guangdong Academy of Agricultural Sciences,Guangzhou 510640,China)
Lecanicillium lecanii;genetic diversity;Diaphorina citri;pathogenicity
Q 939.93;S 476.12
A
10.3785/j.issn.1008-9209.2014.06.304
农业部公益性行业科研专项(201003067-08).
陈国庆,Tel:+86 576 84117695;E-mail:cgq5373@163.com
联系方式:鹿连明,E-mail:minglu79@126.com
2014 06 30;接受日期(Accepted):2014 07 18;
日期(Published online):2015 01 19 URL:http://www.cnki.net/kcms/detail/33.1247.S.20150119.1651.003.html
