5种木兰科珍贵树种早期生长特性研究
2015-04-20冯祥麟胡光平刘正华韩堂松
冯祥麟,胡 刚 ,廖 炜 ,胡光平,刘正华,韩堂松
(贵州省贵阳市林业科技推广站,贵州 贵 阳550003)
1 引言
木兰科(Magnoliaceae)树种是亚热带常绿阔叶林重要组成,在贵州天然林植被中分布较普遍,多数为贵州亚热带常绿阔叶林主林层或亚层组成成分[1]。据初步统计,贵州省有木兰科树种共7属50种,全省80个市县中57个有自然分布,垂直分布海拔为180~2 800 m[2],是木兰科树种资源最为丰富的省区之一。
木兰科树种材质优良,树形美观,花具芳香,很多是列入国家重点保护的珍贵树种,极具用材兼景观开发利用价值。木兰科树种在贵州的普遍分布说明贵州不少地区适合发展木兰科树种,具备引种栽培和开发利用的有利条件。对其中种群数量少且生态幅度较窄的珍贵树种开展造林试验和研究,对保护树种种质资源和丰富营造林树种具有重要意义。营造林实践证明,造林树种与气候、土壤等诸多生态条件相适应是引种和造林成败的重要因素,依据树种生长特性施予相应的技术措施是提高造林成效的有效途径。自20世纪90年代以来,国内不少省区对木兰科树种的生物生态特性和北移引种栽培开展了一些研究,积累了不少成果[3,4]。相对来说,木兰科树种的营造林和幼树早期生长特性研究较少,某些木兰科树种的生物生态特性和林学特性仍有待进一步认知。本文旨在通过5种木兰科珍贵树种的造林试验,对造林效果和幼树早期生长特性开展观测研究,进一步探索5个树种的生物生态特性和林学特性,减少人工造林的盲目性,为本地区造林树种选择和制定相应的营造林技术措施提供参考。
2 试验地基本概况
试验地位于贵州省贵阳市北部开阳县林场,地理位置约北纬26°47',东经106°44'。试验地地处亚热带季风湿润气候,冬无严寒,夏无酷暑,雨热同季。常年年均气温为15.3℃,年≥10℃积温3 562.7~4 278.2℃,最热月(7月)平均温度24.0℃,极端最高温35.4℃;最冷月(1月)平均温度为4.9℃,极端最低温-9.5℃,全年无霜期约270 d,年均日照时数约1 084.8 h,年相对湿度77%,年均降水量约1 200 mm。
试验地为中山丘陵地貌,地形起伏和相对高差变化不大,造林试验地是马尾松采伐迹地,试验小区之间立地与生态条件基本一致,平均海拔为1 320 m,土壤为二叠系砂页岩发育的硅铝质黄壤,土层较深厚,呈酸性,土壤肥力中等。
3 材料与方法
3.1 试验树种与造林
造林试验树种为红花木莲(Manglietiainsignis)、乐昌含笑(Micheliachapensis)、亮叶含笑(Michelia fulgens)、云南拟单性木兰(Parakmeriayunnanensis)和乐东拟单性木兰(Parakmerialotungensis)。造林时间为2008年3月和2011年3月,2次造林全部采用3年生较大规格苗木,苗木平均株高1.15~1.35 m,平均地径为1.3~2.1 cm。造林前块状整地,种植穴规格为60 cm×60 cm×60 cm,株行距为2 m×2 m,种植密度为2 495株/hm2。由于造林苗木规格较大,种植时在确保主要侧枝和冠幅完好的前提下剪掉部分枝叶,苗木携带25 cm左右泥球,树种间按随机块状混交方式种植,造林后连续3年开展除草培土管理。
3.2 试验设计与观测方法
按完全随机区组设计,苗木不分等级,随机种植于试验小区,种植试验区总面积为20 hm2。苗木种植时随机抽取30株测量高度、地径和冠幅,记录初植苗木规格。造林完成当年12月开展幼树成活率调查并抽取30株幼树测量高度、地径和冠幅,以后在每年同一时间开展幼树生长量测定。用Excel2003对调查数据进行统计分析,用最小显著差数法(LSD)对幼树高和地径进行多重比较。
4 结果与分析
4.1 造林成活率
造林成活率调查显示(表1),5个造林树种成活率从高到低依次为:红花木莲98%,云南拟单性木兰和乐东拟单性木兰均为97%,乐昌含笑91%,亮叶含笑80%。3a生较大规格苗木2个年度的造林成活率并无太大差别,除亮叶含笑外其余4个树种造林成活率均在90%以上。从样地幼树生长势观测来看,用较大规格苗木造林减少了杂灌草对幼树早期生长的不利影响和造林后期管护,对幼树生长十分有利。

表1 造林1~4年幼树生长比较
表1中红花:红花木莲;亮叶:亮叶含笑;乐昌:乐昌含笑;云南:云南拟单性木兰;乐东:乐东拟单性木兰。下同。
4.2 幼树高、地径和冠幅生长比较
幼树株高、地径和冠幅生长是评价幼树生长的重要指标,为便于统计分析,把苗木在苗圃地的培育年限一并计入幼树年龄。从造林1~4年幼树的生长结果来看(表1),5个木兰科幼树株高、地径和冠幅由于受苗木移栽影响分别从造林第2年和第3年有明显生长,到造林第4年幼树高生长最大的是云南拟单性木兰(2.67 m)和乐东拟单性木兰(2.42 m),红花木莲和乐昌含笑次之,高生长分别为2.31 m和2.27 m,亮叶含笑树高仅为1.91 m。地径生长则表现为到造林第3年时有明显增长,到造林第4年时红花木莲地径生长量最大,达到5.1 cm,乐昌含笑和云南拟单性木兰地径均为4.7 cm,乐东拟单性木兰和亮叶含笑地径生长较小,分别只有3.6 cm和3.1 cm。
从造林第4年的冠幅生长来看,红花木莲、乐昌含笑和亮叶含笑冠幅均超过1 m,其中冠幅最大的乐昌含笑达到120 cm,云南拟单性木兰和乐东拟单性木兰冠幅生长偏小,分别只有94.5 cm和75.2 cm。云南拟单性木兰和乐东拟单性木兰冠幅生长较慢很可能与其自身的生物生态特性有关。结合以往的研究[5]和样地幼树观测显示,云南拟单性木兰和乐东拟单性木兰侧生枝分生能力较弱,高生长突出,幼树具有较明显的顶端优势,冠幅偏向于中等或偏窄冠型。相反,红花木莲、乐昌含笑和亮叶含笑冠幅则趋向于向外扩张,具有宽冠幅生长趋势。
4.3 幼树连年生长比较
4.3.1 树高连年生长
统计分析结果表明(表2),5个树种树高连年生长有明显差别。树高生长从造林第2年起普遍增速,连年生长量超过35 cm的有红花木莲和云南拟单性木兰,超过20 cm的有乐昌含笑,亮叶含笑和乐东拟单性木兰可能因生态适应等原因树高连年生长未超过20 cm。造林第3年时,红花木莲、云南拟单性木兰和乐东拟单性木兰树高连年生长放缓,其余2个树种树高连年生长有所增加但都未超过30 cm。造林第4年时,5个树种的树高连年生长量最大,其中以云南拟单性木兰和乐东拟单性木兰高生长最为突出,树高连年生长量分别达到57 cm和75 cm,红花木莲和亮叶含笑分别为47 cm和31 cm,乐昌含笑仅29 cm。从整个树高连年生长过程来看,5个树种树高连年生长增速表现出一定的间歇性,连年增长变化呈正态分布,符合幼树生长的普遍规律。云南拟单性木兰和乐东拟单性木兰造林第4年树高连年生长量最大,高生长积累超过红花木莲、乐昌含笑和亮叶含笑,位居5个树种树高首位。从树高生长势来看,云南拟单性木兰、乐东拟单性木兰和红花木莲树高生长明显优于其他2个树种。
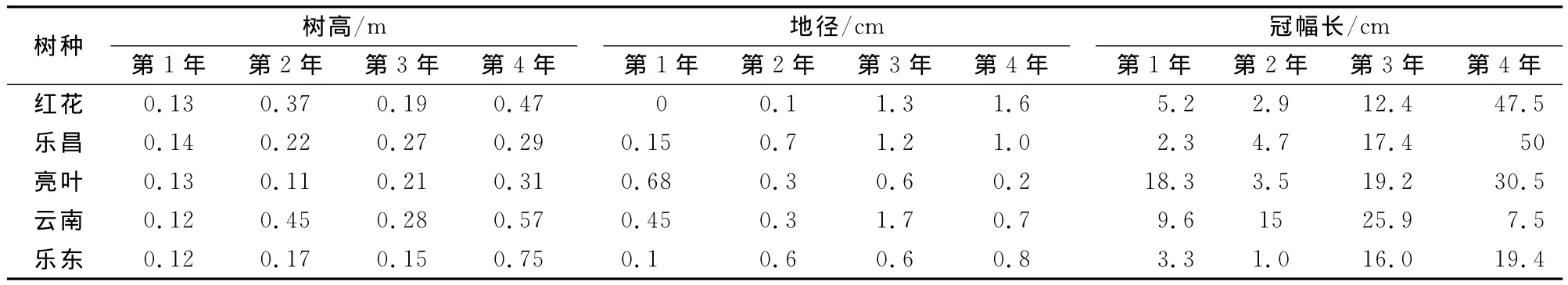
表2 造林1~4年连年生长量比较
4.3.2 地径连年生长
5个树种地径普遍在造林第3年有明显增长,连年生长量1 cm以上的有红花木莲、乐昌含笑和云南拟单性木兰。到造林第4年时,仅红花木莲和乐昌含笑保持1 cm以上的年生长增速,其余3个树种地径连年生长量均未超过1 cm,生长较慢。从表2还可以看出,5个树种地径生长增速普遍滞后于树高。
4.3.3 冠幅连年生长
冠幅的连年生长节奏与地径生长十分接近,造林第3年时开始有明显增长,除红花木莲外其余4个树种冠幅连年生长量均在15 cm以上。造林第4年时连年生长量最大,红花木莲、乐昌含笑和亮叶含笑冠幅连年生长量均超过30 cm,增长量最大的乐昌含笑达50 cm,而云南拟单性木兰和乐东拟单性木兰冠幅连年生长起伏大且增长较慢,增长量远低于其树高,保持窄冠幅的生长特点。
4.4 树高和地径方差分析
通过5个树种造林第4年(7年生幼树)树高和地径的方差分析,得到如下结果(表3)。

表3 树高、地径方差分析
从表3可以看出,树高均方比F=3.676>F(0.05,50)=2.557,且P-value=0.0106<0.05,说明5个树种间树高存在显著差异。地径均方比F=0.458<F(0.05,50)=2.557,结果显示5个树种的地径差异不显著。
通过最小显著差数法(LSD)对5个树种树高进行多重比较(表4),云南拟单性木兰与亮叶含笑、乐东拟单性木兰与亮叶含笑之间树高生长有显著差异,其中云南拟单性木兰与亮叶含笑在LSD(0.01)水平上达到极显著差异。其他树种两两之间树高差异达不到显著程度。

表4 树高生长LSD多重比较
4.5 树高与地径的相关性
树高与地径在生长过程中存在一定的相互关联,通过树高和地径生长的相关分析,以地径(x)为变量树高(y)为函数得到5个树种树高与地径的回归方程(表5)。

表5 树高与地径相关性
由表5得知,红花木莲等5个树种树高与地径存在线性关系,经显著性检验相关性达到极显著水平。
5 结语
幼树生长是随时间推移生长量逐渐增加的过程,生长速度直接影响生长量的积累,导致树种间生长出现差异。红花木莲等5个树种高生长从造林第2年(幼树5年生)开始加速,到造林第4年时树高生长量增速达到最大,无论从年均生长或连年生长增速来看,云南拟单性木兰、乐东拟单性木兰和红花木莲树高生长表现最佳。5个树种地径生长从造林第3年开始增速,增速晚于树高,到造林第4年时红花木莲和乐昌含笑地径生长最快。5个树种的冠幅生长增速与地径同步,到造林第4年时红花木莲和乐昌含笑冠幅均以上一年3倍的连年生长速度增长,是5个树种中冠幅生长最快的。冠幅生长较缓慢的是云南拟单性木兰和乐东拟单性木兰,2个树种冠幅生长增速远低于树高,表现出窄冠幅生长趋势。
树高和地径生长是评价树种的2个重要指标[6],红花木莲等5个树种树高与地径呈显著正相关,但树高生长增速早于地径和冠幅,说明在林分郁闭前期树种间尚未出现明显分化,幼树主要以树高生长方式占据竞争空间,尤其是冠幅生长较慢的偏窄冠树种云南拟单性木兰和乐东拟单性木兰显示出高生长较其他树种更为突出的生长特点。冠幅生长是扩大营养空间和影响树木生产力的因素之一,还影响林分的郁闭,反映树木的长期竞争水平[7]。红花木莲等5个树种在林分郁闭前的幼树早期生长阶段,通过冠幅生长来实现对营养空间的竞争还未突出显现,但通过幼树冠幅生长可预测林分的郁闭进程,同时也可为造林密度和树种配置提供参考,如在造林实践中依据树种冠幅生长趋势设置密度,宽冠幅树种与窄冠幅树种合理搭配等。
幼树早期生长受自身生物生态特性制约,与立地条件和经营管理也有一定关系。有研究指出,气候因子对树木早期生长速度的影响表现为月均温对树高影响最大,月降水对冠幅影响最大,各气候因子对胸径生长影响很小[8],因此,红花木莲等5个木兰科幼树早期生长与气候因子之间的关系还有待观测研究。
[1]李永康.贵州树木手册[M].北京:中国林业出版社,1995:116~124.
[2]徐来福.贵州野生木本花卉[M].贵阳:贵州科技出版社,2006:86~91.
[3]李淑琴,张纪林,肖开生.木兰科树种幼树生长特性的研究[J].植物生态学与地植物学学报,1991,15(4):344~354.
[4]孙起梦,刘兴剑,汤诗杰.南京木兰科观赏植物引种调查及适应性评价[J].安徽农业科学,2008,36(23):9967~9969.
[5]冯祥麟,胡 刚.云南拟单性木兰育苗技术[J].林业适用技术,2007,66(6):25~26.
[6]苏志尧,陈北光,彭华贵,等.北江流域14种乡土阔叶树种苗期生长评价[J].华南农业大学学报,2004,25(4):74~77.
[7]雷柏东,张则路,陈晓光.长白山落叶松等几个树种冠幅预测模型的研究[J].北京林业大学学报,2006,28(6):75~79.
[8]易观路,许方宏,罗建华,等.雷州半岛15个阔叶树种混交林的生长分析[J].生态科学,2004,23(4):346~350.
