调亏灌溉对多年生黑麦草光合特性的影响及阈值的确定
2015-04-08刘译锴郑明珠刘铁军杨有俊刘金荣
刘译锴,郑明珠,马 莉,刘铁军,杨有俊,刘金荣
(1.草地农业生态系统国家重点实验室 兰州大学草地农业科技学院,甘肃 兰州730020;2.河南省永城市试验高级中学,河南 永城476600;3.临夏州种子管理站,甘肃 临夏731100)
草坪是园林绿化中的主要景观之一,具有保护和美化环境、减少噪声污染等重要作用。由于草坪植被的占地面积最大,通常采取一些措施来减缓草坪的生长,以减少草坪的刈割次数,调亏灌溉等可以有效节省养护成本和水资源。叶片水势通过调节气孔开度对光合和植株水分利用有着重要作用[1],调亏灌溉的实质就是通过对土壤水分的管理来控制植株根系的生长从而控制地上部分的营养生长[2]及其叶片水势。也有研究认为,适当的干旱胁迫有助于根系向深层发展,产生更大的地下生物量[3],这对于干旱气候条件下防止草坪退化、延长草坪使用寿命具有重要意义。另外,草坪草在受到干旱胁迫后其光合特性是对土壤水分状况反应最直观、最明显的生理过程之一,水分胁迫会不同程度地影响草坪草的光合速率、蒸腾速率及气孔行为等,有研究表明,光合生理特性对土壤水分反应存在一定的阈值[4],高于或低于阈值的灌溉都不利于草坪草光合及其他生理生化作用的正常进行。因此,研究如何对草坪进行科学的调亏灌溉,控制草坪灌溉用水,刺激根系的补偿生长,调控草坪地上部分的生长过程和光合作用,已成为解决城市绿化草坪节水灌溉问题的关键。
多年生黑麦草(Lolium perenne)是禾本科疏丛型草本植物。由于其种子较大,具有发芽迅速、生长较快、成坪时间短等特点,可与其他草坪草种混播,作为先锋草种,还可用作快速建坪、水土保持及与暖季型草坪的冬季交播[5]。目前,有关多年生黑麦草的研究主要集中在交播性状[6]、坪用特性[7]、饲用价值[8]、生态生物学特性[9-10]及抗旱[11-12]和抗盐碱[13-14]的筛选与评价等方面。有关不同调亏灌溉处理下多年生黑麦草的生理适应性的研究还少有报道。因此,本研究选用两种坪用性较好的多年生黑麦草为材料,研究其在不同调亏灌溉处理下光合特性的变化情况及复水后的恢复程度,以期为干旱半干旱区草坪节水养护提供新的理论依据。
1 材料与方法
1.1 材料
试验材料为美国百绿集团提供的多年生黑麦草品种首相(Premier)和潘多拉(Panterra)。
1.2 试验区地理条件及生长条件
试验始于2013 年7 月10 日,结束于2013 年8月23 日,为期45 d,试验地位于兰州大学榆中校区草坪种植试验区和草地农业科技学院人工智能温室。试验区的地理坐标为35°57' N,104°09' E,典型的黄土高原半干旱气候类型,年平均气温在4.0~5.8 ℃,温差大,降水量少(年降水量300 ~430 mm),分布不均,蒸发量大(年蒸发量为1 450 mm),属于雨养农业区。该温室土柱试验是在前期大田试验的基础上,为了避免气候、土壤等不可控因素对草坪草生理生化反应的影响等问题,从而在温室中模拟当地的气候环境条件进行的补充试验,以求更精确地得出调亏灌溉对草坪草光合生理特性的影响及调亏阈值。
试验期间温室环境条件保持恒定,白天为25 ℃(07:00 -21:00),晚上19 ℃(21:00 -07:00),相对湿度65%,光照强度800 μmol·m-2·s-1。
多年生黑麦草品种首相、潘多拉草皮从试验区的田间小区收集得到,试验材料从当年直接播种成坪培育出的草坪中选取。选择长势健壮均一的草皮移入直径11 cm、高45 cm 的PVC 管中(底部用密质尼龙网封口,此PVC 管又称蒸散仪),每个PVC 管中提前装入3∶ 1 的沙子和原土(土壤类型为黄绵土,质地中壤,土壤pH 为7.44,容重1.29 g·cm-3,有机质含量0.737%,全氮含量0.058 9%,速效磷含量4. 77 mg·kg-1,速效钾含量132. 95 mg·kg-1)混合物,保证每个PVC 管中装入沙土总重一致,且移栽的草皮总重一致,误差控制在±0.2 kg 范围内。移栽后的草皮块于温室中缓苗60 d,待根系生长健壮后开始亏水处理,在此期间根据草皮生长情况追肥(N∶ P∶ K=16∶ 4∶ 8)1 ~2 次,每周浇两次透水,草坪刈剪高度控制在5 cm,保证每周定时修剪一次,且试验各调亏处理开始后不再进行施肥修剪。
1.3 试验设计及土壤水分控制
采用完全随机试验设计,5 个灌溉水平处理,2个品种,4 次重复。试验处理前所有PVC 管中土壤相对含水量均保持在80% ±2%(上限),开始处理后,处理1(CK),使土壤相对含水量始终保持在80% ±2%,处理2、3、4、5 停止浇水,使其干旱,当各处理土壤相对含水量下降到其下限值(处理2,60% ±2%,处理3,50% ±2%,处理4,40% ±2%,处理5,20% ±2%)时再行灌水使土壤相对含水量达到80% ±2%,然后持续进行此“干旱胁迫- 复水-干旱胁迫-复水……”的循环过程。当处理5第3 个干旱阶段土壤相对含水量下降至20% ±2%时,不再复水,所有处理同时停止试验。
用烧杯和量筒给各处理精确加水,各处理土壤的灌水量计算如下:
灌溉量=(0.8 -X)(W0-Wd-W净)。
式中,X 为土壤相对含水量下限值;W0为土壤相对含水量为100%时的蒸散仪总重;Wd为蒸散仪中土壤烘干重;W净为蒸散仪净重。
1.4 测定项目及方法
试验处理期间每天18:00 对所有蒸散仪称重(记为Wt),计算土壤相对含水量(RSWC)。
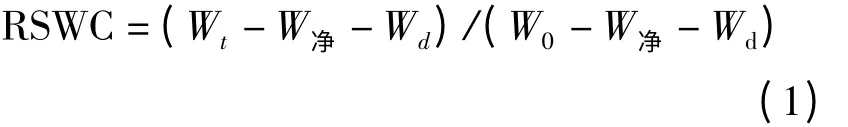
利用Li-6400 型便携式光合仪每3 d 选择晴好天气的09:00 -11:00测定草坪草叶片的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)。叶片叶绿素荧光参数(Fv/Fm)利用便携式调制叶绿素荧光仪PAM-2100 测定(22:00 以后,测定前对植株叶片进行30 min 的暗处理)。草坪质量(TQ)综合得分用数字影像和计算机技术进行综合评定[15]。
1.5 数据处理
测定数据的方差分析和多重比较以及制图分别采用SPSS 18.0 软件和SigmaPlot 10.0 软件。
2 结果与分析
2.1 草坪质量
两品种在处理1(对照)、2、3 下草坪质量(TQ)均保持在80 分左右,且处理2、3 与对照比较差异不显著(P >0.05)。在处理4 下,当RSWC 下降到其下限值时两品种TQ 有显著下降(P <0.05),经过复水后均可恢复到对照水平。在处理5 条件下,每个干旱阶段两品种TQ 都随RSWC 的下降而下降。第1 个干旱阶段,当RSWC 下降到其下限值时首相和潘多拉TQ 分别降至56 分和64 分左右,与对照相比显著下降;复水3 d 后二者均迅速恢复至对照水平。第2、3 干旱阶段结束时,首相TQ 分别降至56分和57 分左右,潘多拉TQ 下降到55 分左右,与对照相比差异显著。二者TQ 在第2 阶段复水3 d 后,均有所上升,但仍低于对照(图1)。
2.2 净光合速率

图1 不同调亏灌溉处理对多年生黑麦草草坪质量的影响Fig.1 Effects of different regulated deficit irrigation treatments on turf quality of perennial ryegrass
对照条件下两草种净光合速率(Pn)都保持较高水平,但由于有时受天气和环境的影响,光照、温度等影响因子导致Pn略呈波动性变化(图2)。在处理2、3 条件下,与对照相比,首相Pn无显著变化(P >0.05)。处理4 条件下,当RSWC 下降到其下限值时,首相Pn下降程度较大。潘多拉Pn从处理2条件下就开始略有下降,在处理3、4、5条件下,Pn下降程度依次增大。处理5 条件下,二者在3 个干旱阶段内,Pn都呈下降趋势,在第2、3 阶段下降显著,在每个干旱阶段结束时首相Pn分别降到对照的25.12%、21. 58%、30.06%,潘 多 拉 分 别 降 到32.80%、15.38%、17.97%。第1、2 个干旱阶段复水3 d 后,首相Pn分别上升为对照的97. 60%和103.80%,潘 多 拉Pn上 升 为 对 照 的73. 24% 和62.47%。说明首相Pn在处理5 条件下复水后可在较短时间内完全恢复,而潘多拉Pn不能完全恢复,由此可见,20%的土壤含水量已经在一定程度上超出了潘多拉的耐受范围。
2.3 蒸腾速率
在对照和处理2、3 下两草种蒸腾速率(Tr)的变化基本相似,趋势不是很明显,处理2、3 与对照间相比差异不显著(P >0.05)(图3)。但在处理4、5 条件下,二者Tr显著下降(P <0.05)。首相Tr在处理5 条件下,第1、2 干旱阶段结束时,分别下降为对照的26.37%和28.82%。第3 干旱阶段结束时,Tr下降为对照的25.15%。第1 阶段复水后3 d,Tr上升为对照的90.16%;第2 阶段复水后3 d,Tr上升为对照的93.08%,说明首相Tr在20%的土壤含水量下基本得以恢复。潘多拉在3 个干旱阶段结束时,Tr分 别 下 降 为 对 照 的36. 78%、20. 22%、21.29%。第1、2 阶段复水3 d 后,Tr分别上升为对照的75.21%和81.31%。两个干旱阶段复水结束后,Tr都不能完全恢复。

图2 不同调亏灌溉处理对多年生黑麦草净光合速率的影响Fig.2 Effects of different regulated deficit irrigation treatments on photosynthetic rate of perennial ryegrass
2.4 气孔导度
对照与处理2、3 条件下两草种气孔导度(Gs)的变化趋势都不明显,但在处理4、5 条件下下降程度依次增大(图4)。处理5 条件下,两草种在3 个干旱阶段内Gs都不断减小,与对照比较差异显著(P <0.05)。3 个干旱阶段结束时,首相Gs分别下降为对照的32.40%、27.87%、28.65%,潘多拉下降为对照的30.73%、23.76%、22.24%。第1、2 干旱阶段复水后3 d,首相Gs分别上升为对照的89.15%和102.56%,可见首相第2 个干旱阶段后复水Gs恢复程度大于第1 阶段,这说明在经历了一次干旱胁迫后,首相Gs对同等干旱胁迫伤害的恢复能力提高。而潘多拉两阶段复水后Gs分别上升为对照的80.35%和65.97%,其Gs的恢复程度与首相恰恰相反,即第2 个干旱阶段复水后Gs恢复程度小于第1 阶段。这说明处理5 的灌溉下限已经大大超出了潘多拉的耐受范围,在经历了一次干旱胁迫后,其Gs对同等干旱胁迫伤害的恢复能力非但没有提高,反而因为经历两次较重的干旱胁迫而下降。
2.5 胞间CO2 浓度
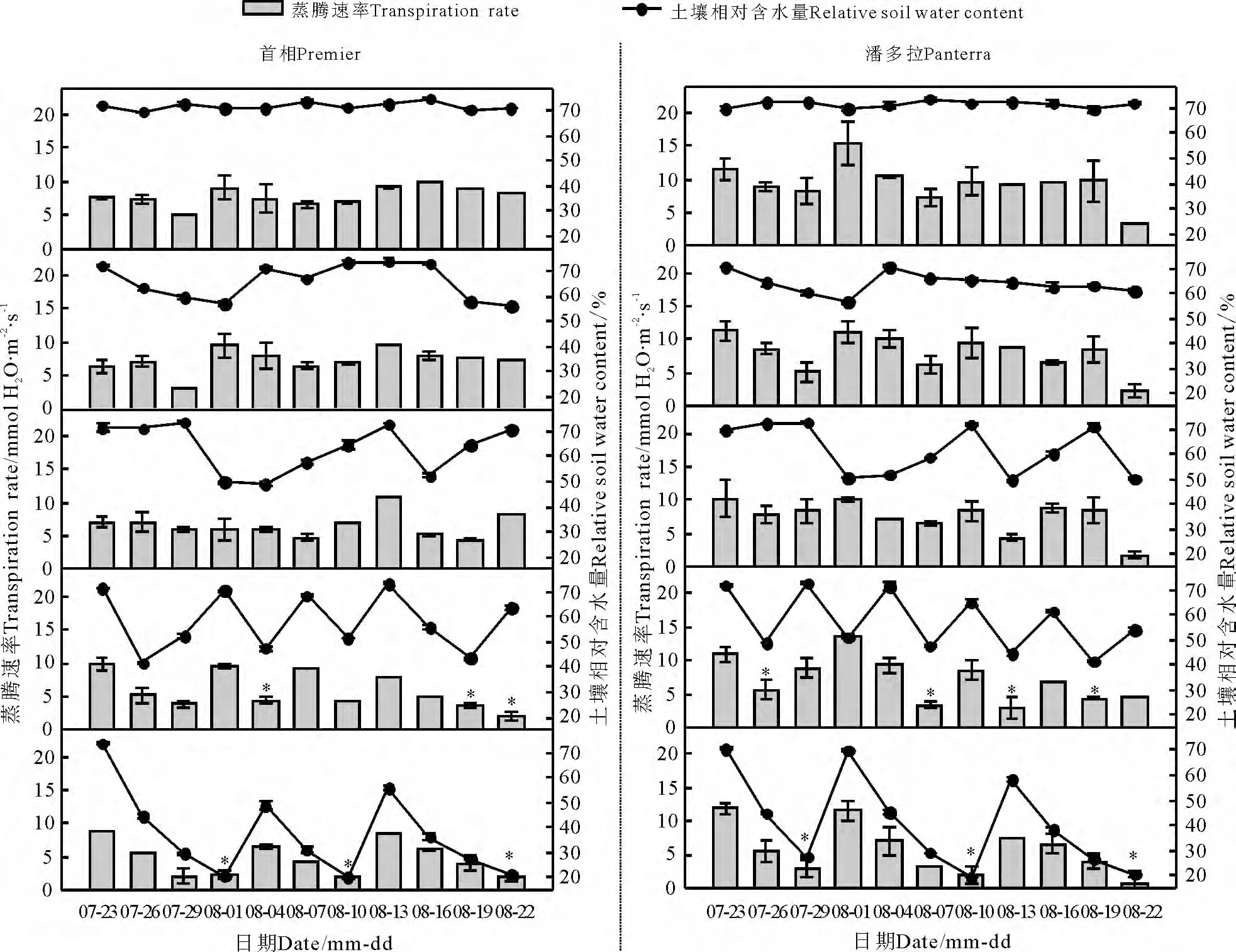
图3 不同调亏灌溉处理对多年生黑麦草蒸腾速率的影响Fig.3 Effects of different regulated deficit irrigation treatments on transpiration rate of perennial ryegrass
两草种胞间CO2浓度(Ci)在对照下都维持较高值,且变化平稳(图5)。处理2、3、4 条件下,两草种Ci随RSWC 的下降而下降,处理2 与对照相比差异不显著(P >0.05)。处理5 条件下两草种在3 个干旱阶段内Ci先持续下降,阶段末又略有上升;在3 个干旱阶段结束时首相Ci分别降至对照的48.0%、53. 4%、43. 2%,潘 多 拉 分 别 降 至50%、46.38%、49.97%。第1、2 阶段复水后3 d,首相Ci分别上升为对照的81.58%和86.58%,说明其Ci在处理5 条件下虽第2 阶段复水后恢复程度略高于第1 阶段,但仍无法完全恢复;而潘多拉Ci分别上升为对照的87%和77.19%,两个干旱阶段复水结束后,Ci都不能完全恢复,且第2 阶段恢复能力下降,这说明处理5 条件下反复的调亏灌溉处理会加重对潘多拉的胁迫伤害。
2.6 荧光参数
对照与处理2、3 条件下首相的荧光参数(Fv/Fm)均可保持0.8 以上的较高水平,三者间差异不显著(P >0.05)(图6)。在处理4 条件下,当RSWC 降至其下限时,Fv/Fm下降至0.73。处理5条件下首相Fv/Fm在3 个干旱阶段都呈现下降趋势,每个阶段结束时,Fv/Fm降至0.64,与对照相比差异显著(P <0.05),各干旱阶段复水3 d 后Fv/Fm可以恢复,但比对照水平略低。潘多拉在处理1、2条件下,Fv/Fm都可保持在0.8 以上的较高水平,差异不显著;在处理3、4 条件下,当RSWC 降至各下限时,Fv/Fm分别下降至0.78、0.66。处理5 条件下潘多拉第1 干旱阶段结束时Fv/Fm下降至0.57,而第2、3 阶段结束时Fv/Fm分别下降至0.54、0.51,与对照相比差异显著;第1、2 干旱阶段复水3 d 后,Fv/Fm虽有较大恢复,但仍低于对照。
3 讨论

图4 不同调亏灌溉处理对多年生黑麦草气孔导度的影响Fig.4 Effects of different regulated deficit irrigation treatments on stomatal conductance of perennial ryegrass
调亏灌溉研究的核心内容之一就是水分胁迫后复水对作物生理指标的影响[16]。而水分亏缺会影响到植物的生长和代谢,其中对光合作用的影响尤为突出[17]。水分在植物光合作用的进程中所占的比例并不多[18],但水分的供应量多少会影响其代谢和物质及气体交换,从而会直接或间接影响光合作用的各项指标,包括净光合速率(Pn)、蒸腾速率(Tr)和气孔导度(Cs)[19]等。
草坪质量(TQ)能从表观上反映草坪草的整体表现,土壤相对含水量(RSWC)能间接反映植物水分亏缺的程度[20]。有研究表明,草坪质量与耗水量并不一定呈正相关,当土壤含水量增加至一定程度时,若继续灌溉并不会对草坪质量有所改善,只会增加蒸散耗水量[21]。本研究结果表明,在处理4、5 条件下,每个干旱阶段两品种TQ 随RSWC 的下降而下降,说明水分胁迫引起两个品种的根系从土壤吸收的水分剧减,影响了供试草种的水分吸收[22],从而导致TQ 下降。
由于植物Pn与Tr、Gs、胞间CO2浓度(Ci)的变化密切相关[23-24],因而水分胁迫下光合作用主要受两方面限制:一是细胞中CO2的浓度,与Gs密切相关(气孔因素限制);二是叶肉细胞对CO2的同化能力,与其自身的光系统受损程度密切相关[25](非气孔因素限制)。Farquhar 和Sharkey[26]认为,通过同时观察气孔导度的大小和胞间CO2浓度的变化才能判断Pn的降低是否是由于气孔因素限制造成的。如果气孔导度的下降同时引起了胞间CO2浓度的降低,那么是由于气孔因素限制所致;反之,是由于非气孔因素限制所致。本研究结果显示,当RSWC的下限值高于40%时,两草种Gs、Ci下降与Pn基本保持一致,复水后Pn、Gs、Ci都能迅速完全恢复。根据Farquhar 和Sharkey[26]的观点可以判断出Pn的降低主要是由于气孔限制。因此,本研究认为灌溉下限在40%以上(包含40%)时,RSWC 的下降引起Gs降低,限制了进入叶肉细胞的CO2含量,导致光合作用缺乏原料,使Pn下降[27]。此时光系统本身并未受损害,复水后Pn、Gs、Ci也可迅速恢复。

图5 不同调亏灌溉处理对多年生黑麦草胞间二氧化碳浓度的影响Fig.5 Effects of different regulated deficit irrigation treatments on intercellular CO2 concentration of perennial ryegrass
而当RSWC 的下限值低于20%时,两草种在干旱阶段末期Pn、Gs下降的同时,Ci反而出现了不同程度的上升或是停止下降。这说明灌溉下限在20%时,非气孔因素逐渐成为限制Pn的主要因素,但不同草种非气孔限制出现的时间不同。首相在处理5 条件下当RSWC 降至20% ~30%时,Ci开始表现出上升趋势,此时Pn与Gs都不断下降。但是,由于所测数据不够全面,无法准确地推断首相开始非气孔限制时对应的RSWC 值和Ci下降的最低值,不过根据现有数据推测,首相可能在RSWC 处于20%~25%时表现出非气孔限制,此时水分亏缺已经显著降低了叶肉细胞的光合能力,Ci最低可降至103.89 μmol·m-2。而潘多拉在RSWC 降至30%时,就表现出非气孔限制。
首相复水3 d 后Pn、Gs都能较快恢复,甚至超过对照,且第2 阶段恢复速度大于第1 阶段。这主要与首相非气孔限制发生较晚、复水时光系统损伤还不算太严重,因此在3 d 内就可以完成生理机能的修复,Pn迅速恢复。而Gs的超补偿恢复可能与Pn迅速增大、对CO2的大量需求有关,导致气孔导度逐渐增大[28];另外复水后叶水势及膨压的恢复也是Gs上升的主要原因[29]。
潘多拉复水后Pn、Gs、Ci均无法完全恢复,且恢复幅度有下降的趋势。这主要是因为潘多拉非气孔限制出现较早,光系统相对受损时间较长,导致其在复水后短时间内生理机能无法完全恢复。而复水后生理机能还未完全恢复时又重新面临新一轮的水分胁迫,上一轮胁迫效应的累加也使其受胁迫更加严重,复水后恢复能力也逐渐降低。
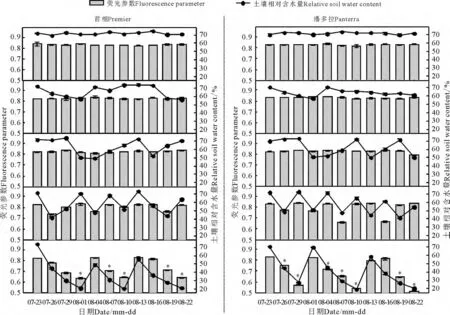
图6 不同调亏灌溉处理对多年生黑麦草荧光参数的影响Fig.6 Effects of different regulated deficit irrigation treatments on fluorescence parameters of perennial ryegrass
蒸腾作用可以反映植物的水分代谢状况和水分利用效率,对于植物的生命活动也具有重要作用。蒸腾作用的实质不仅可以将溶入水中的营养物质运送到植物体内的各个部分,而且水分的散失带走一部分热量,使得植物体表面温度不致于太高。植物体主要通过气孔对蒸腾作用进行调节,而这种调节能力是植物体能很好地适应外界环境的重要保障之一[30]。本研究结果显示两草种Tr与Gs基本同步下降,即在水分胁迫下植物的Tr和Gs互相影响。土壤水分较充足时,植物通过蒸腾作用消耗大量水分,引起水分亏缺,进而使Gs下降。Gs降低后,植物的Tr也随之下降,使水分亏缺不至于更加严重,从而维持一定的气孔开度,保证光合作用的正常进行[31-32]。
叶绿素荧光参数常被用作标明环境胁迫程度的指标和探针[33]。本研究结果表明,灌溉下限低于50%后,首相、潘多拉Fv/Fm降幅随着灌溉下限降低而增大,首相降幅低于潘多拉。这说明同一水分胁迫下首相光抑制出现晚于潘多拉,光抑制程度也小于潘多拉。当RSWC 的下限值为20%时,首相Fv/Fm下降程度较大,但复水后都可恢复至对照水平。说明干旱阶段末期,水分胁迫虽然使两草种的PSⅡ反应中心光合潜能下降,但并未严重破坏光合基质,复水后可较快恢复。而潘多拉Fv/Fm急剧下降,说明其光合基质受损较严重,复水后无法恢复至对照。
水分胁迫下,植物叶片的气孔导度、光合速率、蒸腾速率和胞间CO2浓度之间必然存在一定的联系,而气孔导度是反映叶片气体交换的重要指标,正常情况下,植物叶片气孔的开闭主要受光照强度和CO2浓度控制,但是当叶片受到干旱胁迫时,气孔就会关闭,以避免大量蒸腾失水对植物造成严重伤害。有研究表明在植物缺水时,Pn、Gs、Tr均会显著降低,但是用于光合作用所消耗的水分仅仅是从土壤中吸收的一小部分,其余都为蒸腾失水,可见光合速率的降低主要是受气孔和非气孔因素作用[34]。叶绿素荧光参数Fv/Fm可有效反映叶片的光合机能,水分胁迫下PSⅡ光系统通过调节电子传递速率和光化学效率降低CO2同化能力,以热散失的形式减轻过剩光能对光系统的损害[35]。草坪质量做为草坪草外部生长形态的综合性指标,在遭受环境胁迫时其与各项光合生理指标之间的联系和生理机制还需要更多深入的研究与探讨。
4 结论
随着调亏灌溉亏水程度的加剧,首相和潘多拉两草种的草坪质量及各项光合生理指标均呈现出一定程度的下降趋势,经过复水后,各指标的恢复程度和恢复速度各有所不同,但总体来看,潘多拉的抗旱性能不及首相。在本研究中综合不同处理下对两草种各项生理指标的影响认为首相适宜将RSWC 为40%的灌溉下限作为调亏灌溉阈值,潘多拉适宜将RSWC 为50%作为灌溉下限,在此胁迫程度下,各项指标虽有一定程度的下降,但并未影响到其生理内部机理,且复水后均可在短时间内得以恢复甚至超偿恢复。本研究对草坪草抗旱性能的研究及最佳调亏阈值的确定还需结合更多的生理、形态指标进行更深入和更系统的研究,为西北干旱半干旱地区调亏灌溉技术的运用提供坚实的基础和有力的科学支撑。
[1] Blackman P G,Davies W J.Root to shoot communication in maize plants of the effects of soil drying[J].Experimental Botany,1985,36:39-48.
[2] Chalmers D J,Wilson I B.Productivity of peach trees:Tree growth and water stress in relation to fruit growth and assimilate demand[J].Annals of Botany,1978,42(2):285-294.
[3] Henry A,Veeresh R P,Gowda,Torresa R O,Rolando O,Kenneth L,Mcnally.Variation in root system architecture and drought response in rice (Oryza sativa):Phenotyping of the Oryza SNP panel in rainfed lowland fields[J].Field Crops Research,2011,120(2):205-214.
[4] 刘建利,石元春,罗远培.作物生对土壤水分变动的双重效应[J].生态农业研究,1997,3(5):18-21.
[5] 张自和,柴琦.草坪学通论[M].北京:科学出版社,2009.
[6] 于玉红,王亮,黄晓露,杨志民.6 个多年生黑麦草品种的交播性状比较[J].草业科学,2009,26(8):172-176.
[7] 兰剑,张丽霞.多效唑对多年生黑麦草坪用性状的影响[J].草原与草坪,2002(2):32-38.
[8] 张磊,刘东燕,邵涛.黑麦草的饲用价值及其应用前景[J].草业科学,2008,25(4):64-69.
[9] 于应文,蒋文兰,徐震,冉繁军. 刈割对多年生黑麦草分蘖与叶片生长动态及生产力的影响[J]. 西北植物学报,2002,22(4):900-906.
[10] 占丽平,鲁剑巍,杨娟,李文西,李小坤,刘晓伟.施肥对黑麦草生长和产量的影响[J].草业科学,2011,28(2):260-265.
[11] 石永红,万里强,刘建宁,王运琦,吴欣民,李向林.干旱胁迫对6 个坪用多年生黑麦草品种抗旱性的影响[J].草地学报,2009,17(1):52-57.
[12] 马艳华,宋瑜,张洪荣.烯效唑对黑麦草抗旱生理特性的影响[J].草业科学,2009,26(5):169-173.
[13] 张振霞,刘萍,杨中艺.25 个多年生黑麦草品种萌发期对盐胁迫的抗性研究[J].草业科学,2007,24(2):14-19.
[14] 陈国雄,李定淑.盐胁迫对多年生黑麦草和草地早熟禾种子萌发影响的对比研究[J].草业科学,1996,13(3):41-44.
[15] Michael D,Richardson M D,Karcher D E,Hignight K,Douglas E,Debra R.Drought tolerance and rooting capacity of Kentucky bluegrass cultivars[J].Crop Science,2008,48(6):2429-2436.
[16] 丁端锋,蔡焕杰,王健,张旭东.玉米苗期调亏灌溉的复水补偿效应[J].干旱地区农业研究,2006,24(3):64-67.
[17] Munns R.Physiological processes limiting plant growth in saline soils:Some dogmas and hypotheses[J].Plant,Cell & Environment,1993,16(1):15-24.
[18] 韩烈保,田地,牟新待.草坪建植与管理手册[M].北京:中国林业出版社,1999.
[19] 王英宇,杨建,韩烈保.不同灌溉量对草坪草光合作用的影响[J].北京林业大学学报,2006(12)(S1):27-29.
[20] 余玲,王彦荣,Garnett T,Auricht G,韩德梁. 紫花苜蓿不同品种对干旱胁迫的生理响应[J]. 草业学报,2006,15(3):75-85.
[21] 邪文刚,王艳艳,寇立蜎,邬龙.不同水分处理对高羊茅草耗水及生长质量的影响[J].中国水土保持,2010(2):39-41.
[22] 谢晓蓉,刘金荣,李唯,孙吉雄,杜建雄.干热对草地早熟禾生理反应与坪用质量的影响[J].草地学报,2007,15(6):572-576.
[23] Flexas J,Medrano H.Drought inhibition of photosynthesis in C3plant:Stomatal and non-stomatal limitation revisited[J].Annals of Botany,2002,89(2):183-189.
[24] 张文丽,张彤,吴冬秀,吴冬秀,张岁岐,山仑.土壤逐渐干旱下玉米幼苗光合速率与蒸腾速率变化的研究[J].中国生态农业学报,2006,14(2):72-75.
[25] 廖建雄,王根轩.谷子叶片光合速率日变化及水分利用效率[J].植物生理学报,1999,25(4):362-368.
[26] Farquhar G D,Sharkey T D.Stomatal conductance and photosynthesis[J].Annual Review of Plant Physiology,1982,33:317-345.
[27] Martin A J,Parry P,John A,Alfred J,Stephen J,Carmo S,Elizabete A.Rubisco activity:Effects of drought stress[J].Annals of Botany,2002,89(7):832-839.
[28] Lal A,Ku M S B,Edwards G E.Analysis of inhibition of photosynthesis due to water stress in the C3species Hordeum vulgare and Vicia faba:Electron transport,CO2fixation and carboxylation capacity[J].Photosynthesis Research,1996,49:57-69.
[29] 李国龙,孙亚卿,张少英.水分胁迫对甜菜幼苗光合作用的影响[J].内蒙古农业大学学报,2012,33(1):68-72.
[30] 赵春明.亏水处理对梨树生理生态指标影响的研究[D].杨凌:西北农林科技大学,2009.
[31] 余彦波,刘桐华.植物光效生态学研究Ⅰ.小麦光合作用午休的原因[J].生态学报,1985,5(4):336-342.
[32] 许大全.光合作用“午睡”现象的生态、生理与生化[J].植物生理学通讯,1990,26(6):5-10.
[33] 冀天会,张灿军,杨子光,郭军伟,孟丽梅,马雯.冬小麦叶绿素荧光参数与品种抗旱性的关系[J].麦类作物学报,2005,25(4):64-66.
[34] Irigoyen J J,Fmerich D W,Sdnchez Diaz M.Alfalfa leaf senescence induced by drought stress:Photosynthesis,hydrogen peroxide metabolism,lipid peroxidation and ethylene evolution[J].Physiologia Plantarum,1992,84(1):67-72.
[35] 卜令,张仁和,韩苗苗.苗期玉米叶片光合特性对水分胁迫的响应[J].生态学报,2010,30(5):1184-1191.
