高温对金荞麦光合气体交换和叶绿素荧光特性的影响
2015-04-08赵丽丽向清华赵丽芬
赵丽丽,邓 蓉,向清华,夏 林,赵丽芬
(1.贵州大学动物科学学院草业科学系,贵州 贵阳550025;2.贵州省畜牧兽医研究所,贵州 贵阳550005;3.贵阳市农业委员会,贵州 贵阳550003;4.贵州省草原监理站,贵州 贵阳550002)
高温是严重影响植物生长的自然灾害之一。遇到高温胁迫时,植物可以利用外部形态变化、渗透调节物质含量增加、抗氧化酶活性提高、光合机制调控等方面来适应和抵抗高温胁迫。光合作用是植物生长发育的基础,决定着植物的产量和品质[1]。由于其在植物进化过程中的稳定性而成为植物生理生态研究的重要内容[2]。叶片气体交换和叶绿素荧光测定技术具有测定快速且无损伤的特点,被认为是判断植物生长和抗逆性强弱的重要指标[3],已经在玉米(Zea mays)、马尾松(Pinus massoniana)等多种植物逆境生理及抗性鉴定中得到应用[4-5]。郝婷等[6]在高温胁迫下瓜类的光合生理研究中指出高温胁迫导致叶片净光合速率、气孔导度、胞间CO2浓度、蒸腾速率、PSⅡ的实际光化学效率、PSⅡ的光化学量子产量、光化学猝灭系数等总体上均显著降低,利用光合参数鉴定出两个丝瓜(Luffa cylindrica)品种,特别是“五叶香”丝瓜的耐热性较强,可作为耐高温瓜类作物的砧木,该结论为生产实践提供了理论依据。
金荞麦(Fagopyrum dibotrys)属于蓼科多年生草本植物,广泛分布于我国贵州、四川、云南、江西、浙江、江苏、湖南、湖北、陕西等地,具有清热解毒、消肿止痛、抑制癌细胞生长等药效。1999 年被列入国家重点保护野生植物名录,成为国家二级重点保护的野生植物资源之一[7]。同时,金荞麦青绿多汁,猪、马、牛、羊及家禽等喜食,在贵州地区广泛应用于养猪业,并取得了较好的效果[8]。但在生产利用过程中,金荞麦常受到高温等逆境的影响,造成产量下降,限制了其在生产中的推广应用。因此,以8 个野生金荞麦种质为试材,对比研究高温胁迫下各种质光合系统的功能,探讨金荞麦对高温的光合适应机制,评价其耐热能力,以期为开发、利用金荞麦的耐热种质资源提供理论依据。
1 材料与方法
1.1 材料
供试野生金荞麦种质8 份,详见表1。
1.2 试验方法
1.2.1 高温胁迫 2013 年10 月在田间选择大小一致的8 个金荞麦种质的块根播种于由河沙、蛭石、腐殖质土按1∶ 1∶ 1 比例混合的花盆中,各材料种植12 盆,每盆3 株,置于温室中进行日常管理,生长60 d 后选择大小基本一致、长势良好的植株(株高15 ~20 cm),在人工培养箱内分别设置35 和45 ℃进行高温处理,以22 ℃处理为对照,每个处理4 次重复。期间维持温室内空气相对湿度为80%。在高温处理前和连续高温胁迫15 d 后分别测定各种质气体交换参数和叶绿素荧光参数。
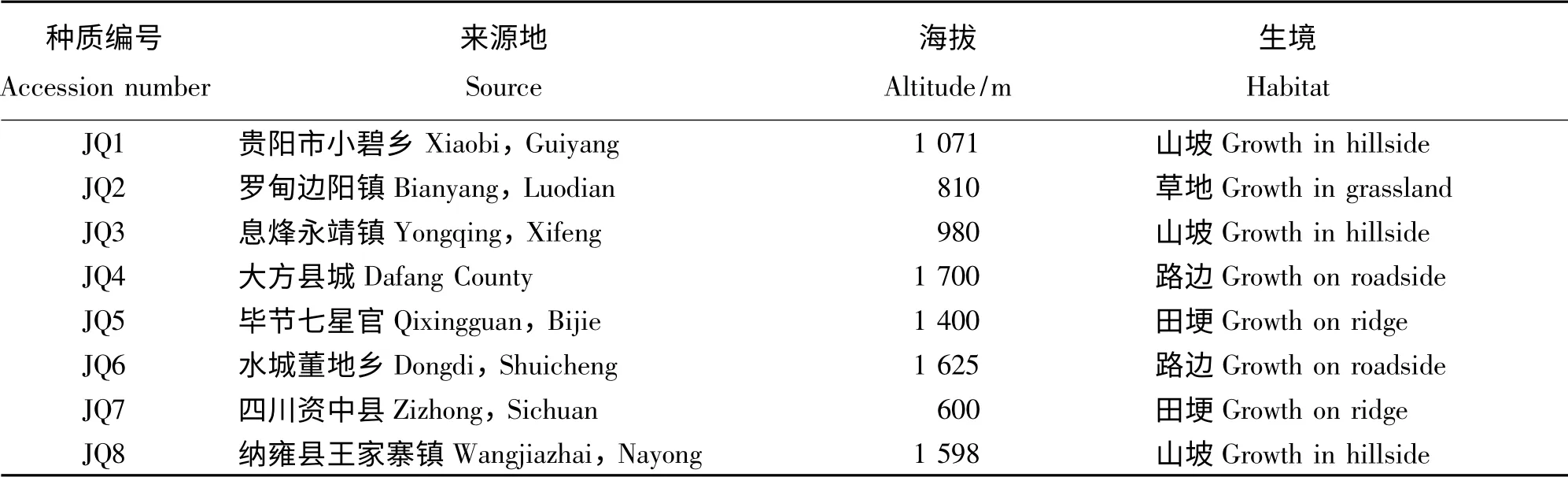
表1 供试种质来源Table 1 Sources of germplasm for the study
1.2.2 气体交换参数测定 上午10:00 选取中上部中等大小、健康的叶片,用LI-COR 公司6400-15透明叶室测定叶片净光合速率(Pn)、气孔导度(Gs)和蒸腾速率(Tr),并计算叶片瞬时水分利用效率(WUE),WUE=Pn/Tr。每个处理选择3 株(每盆1株)进行测定。
1.2.3 叶绿素荧光参数测定 用LI-COR 公司6400-40 叶绿素荧光叶室测定各指标。测定前一天选取中上部中等大小、健康的叶片,用锡纸包裹叶片暗适应一夜,第2 天测定叶片的初始荧光(Fo)和最大荧光(Fm)。之后用活化光处理暗适应的叶片,测定稳态荧光产量(Fs)、最大荧光产量(Fm')等参数。最后将光合仪与电脑连接将PS Ⅱ潜在活性(Fv/Fo)、PSⅡ最大光化学效率(Fv/Fm)、PSⅡ实际光化学量子产量(Yield)、光化学猝灭系数(qP)和非光化学猝灭系数(NPQ)等指标值导出。每个处理选择3 株(每盆1 株)进行测定。
1.3 数据处理与统计分析
1.3.1 主成分分析 计算各指标间的相关系数矩阵,求相关系数矩阵的特征根和特征向量以及各主成分的贡献率和累计贡献率;按累计贡献率大于或等于85%确定主成分个数,并建立主成分方程[9-10]。
1.3.2 隶属函数分析 试验材料各主成分的隶属函数值按公式(1)求得。

式中,U(xi)为各种质第i 个主成分的隶属函数值,xi为各种质某一主成分得分值,ximax和ximin分别为所有参试种质中第i 个主成分的最大值和最小值。
1.3.3 主成分权重 根据主成分贡献率的大小,按公式(2)计算各主成分的权重。
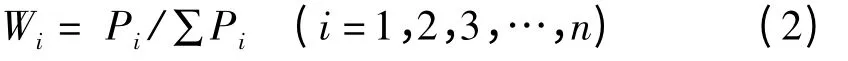
式中,Wi表示第i 主成分在所有主成分中的重要程度;Pi为各种质第i 个主成分的贡献率。
1.3.4 综合评价值 各材料的耐热性综合评分值按公式(3)计算。

式中,D 为高温胁迫下各种质用主成分评价所得的耐热性综合评分值。
原始数据由仪器与计算机联机输出。使用Microsoft Excel 2003 进行数据整理和作图,SPSS 18.0进行差异显著性分析(LSD 法)和主成分分析。
2 结果与分析
2.1 高温胁迫对金荞麦叶片气体交换参数的影响
2.1.1 净光合速率 对高温胁迫下金荞麦叶片净光合速率(Pn)测定分析(图1)可知,22 ℃时,8 个金荞麦种质的Pn介于14.440 ~15.190 μmol·m-2·s-1,但种质间差异不明显。随处理温度的增加,各种质的Pn均呈下降趋势。35 ℃处理时,除种质JQ6、JQ2和JQ3 与对照间无显著差异外,其他种质均显著低于对照(P <0.05)。45 ℃处理时,各种质Pn均显著降低,其中JQ6、JQ2 和JQ3 降幅相对较小,分别为对照的62.95%、54.61%和46.56%。种质JQ5 和JQ7的Pn降幅相对较大,分别为对照的29. 90% 和29.23%。由此可知,金荞麦种质JQ6、JQ2 和JQ3 的Pn受高温胁迫的影响小,下降幅度较小,JQ5 和JQ7的Pn受高温的影响较大,下降幅度较大,JQ8、JQ1和JQ4 下降幅度居中。
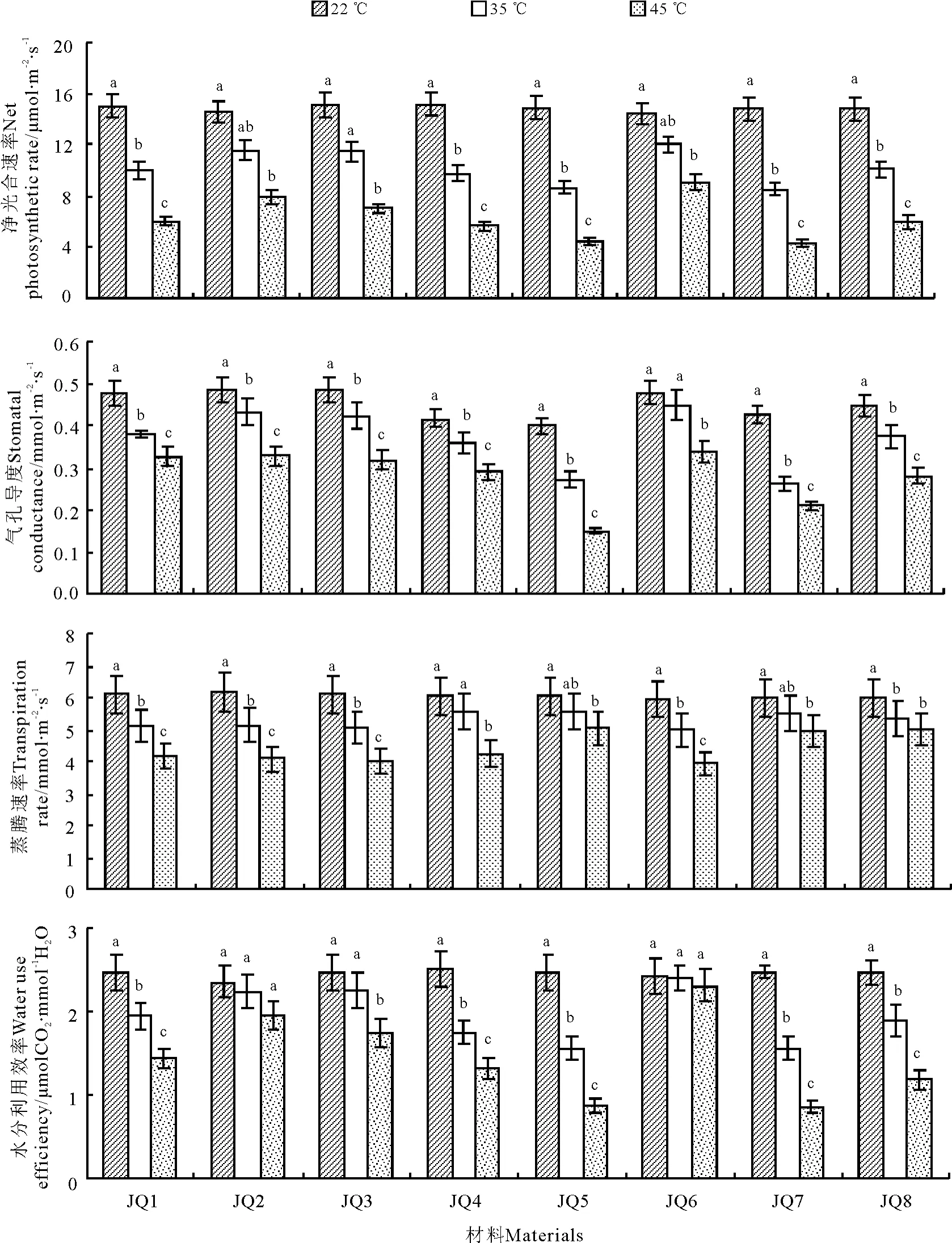
图1 高温胁迫对金荞麦气体交换参数的影响Fig.1 Effects of high temperature stress on gas exchange parameters of Fagopyrum dibotrys
2.1.2 气孔导度 22 ℃时,8 个金荞麦种质的Gs介于0.400 ~0.509 mmol·m-2·s-1(图1),差异不明显。随高温胁迫强度的增加,各种质的Gs均呈降低趋势,除种质JQ6 外,其余7 个种质的Gs均在35 ℃处理时显著降低(P <0.05)。种质JQ6 的Gs在各高温处理梯度下降低幅度均最小,分别为0.453 mmol · m-2· s-1(35 ℃处 理)和0. 339 mmol·m-2·s-1(45 ℃处理)。高温胁迫下,种质JQ7 和JQ5 的Gs降低幅度较大,45 ℃处理时分别降至0.210和0.150 mmol·m-2·s-1;其余种质的Gs降低幅度居中,45 ℃处理时的Gs介于0.292 ~0.330 mmol·m-2·s-1。说明高温对种质JQ6 的Gs影响小且慢,对种质JQ7 和JQ5 的影响大,对其他种质的影响居中。
2.1.3 蒸腾速率 对高温胁迫下金荞麦叶片蒸腾速率(Tr)的测定分析(图1)表明,22 ℃时,8 个金荞麦种质的Tr介于5.970 ~6.200 mmol·m-2·s-1,种质间无明显差异。随高温胁迫强度增加,各种质的Tr逐渐降低。35 ℃处理时,种质JQ4、JQ5 和JQ7的Tr降低幅度较小,与对照间均无显著差异;其余种质均显著低于对照(P <0.05)。45 ℃处理时,种质JQ5、JQ8 和JQ7 的Tr降低幅度较小,分别为5.060、5. 020 和4. 990 mmol·m-2·s-1;种 质JQ6、JQ3 和JQ2 的Tr降低幅度较大,分别为3.940、4.020和4.090 mmol·m-2·s-1;种质JQ1 和JQ4 的Tr降低幅度居中。由此可见,在高温胁迫下,金荞麦各种质均采取降低蒸腾速率的形式适应高温,其中种质JQ6、JQ3 和JQ2 降低蒸腾速率适应高温的表现更突出,种质JQ1 和JQ4 居中,种质JQ5、JQ8 和JQ7 的适应表现最弱。
2.1.4 水分利用效率 对高温胁迫下金荞麦叶片水分利用效率(WUE)的测定分析(图1)表明,22 ℃时,8 个金荞麦种质的差异较小(2. 352 ~2. 502 μmolCO2·mmol-1H2O),各种质间无明显差异。随高温胁迫强度增加,各种质的WUE 降低,种质JQ6和JQ2 的WUE 降低幅度较小,各处理温度间差异均无显著差异。种质JQ7 和JQ5 的WUE 降低幅度最大,35 ℃处理时两个种质的WUE 均为1.562 μmol CO2·mmol-1H2O,45 ℃处理时两个种质的WUE 分 别 为0. 867 和0. 882 μmolCO2·mmol-1H2O。其余种质的WUE 降低幅度居中,35 ℃处理时的WUE 介于1.753 ~1. 948 μmolCO2·mmol-1H2O,45 ℃处 理 时 的WUE 介 于1. 183 ~1. 751 μmolCO2·mmol-1H2O。由此可知,在各高温胁迫梯度下,种质JQ6 和JQ2 能维持较高的WUE,而种质JQ7 和JQ5 的WUE 受影响较大,其他种质居中。
2.2 高温胁迫对金荞麦叶片叶绿素荧光参数的影响
2.2.1 PSⅡ潜在活性 对高温胁迫下金荞麦叶片PSⅡ潜在活性(Fv/Fo)的测定分析(图2)表明,22℃处理时,8 个金荞麦的Fv/Fo值介于4. 116 ~5.072,种质间无明显差异。随高温胁迫强度的增加,各种质的Fv/Fo均呈下降趋势。35 ℃处理时,种质JQ5、JQ7 和JQ4 降幅较大,均显著低于对照(P<0.05),其余种质与对照间无显著差异(P >0.05)。45 ℃处理时,各种质的Fv/Fo均显著低于对照,其中种质JQ5、JQ7 和JQ4 较小,分别为2.234、2.429 和2.831;种质JQ3、JQ6 和JQ2 较大,分别为3.605、3.562 和3.528;其余种质居中。由此可知,金荞麦种质JQ3、JQ6 和JQ2 的Fv/Fo受高温影响较小,种质JQ5、JQ7 和JQ4 的Fv/Fo受高温影响早且大,种质JQ1 和JQ8 受高温的影响居中。
2.2.2 最大光化学效率 正常温度条件下(22℃),8 个金荞麦种质的最大光化学效率(Fv/Fm)介于0.787 ~0.815,种质间无明显差异。随处理温度的增加,8 个金荞麦种质的Fv/Fm逐渐降低。35 ℃处理时,各金荞麦种质的Fv/Fm降低幅度较小,与对照间均无显著差异。45 ℃处理时,种质JQ5、JQ7、JQ4 和JQ3 下降幅度较大,分别为0. 631、0. 647、0.654和0.672,均显著低于对照(P <0.05);而另外4 个种质降幅仍较小,与对照间无显著差异。可见,金荞麦种质JQ5、JQ7、JQ4 和JQ3 受高温胁迫的影响较大,另外4 个种质受高温的影响较小。
2.2.3 实际光化学量子产量 对高温胁迫下金荞麦叶片实际光化学量子产量(Yield)的测定分析(图3)表明,22 ℃处理时,各金荞麦种质Yield 值介于0.587 ~0.652,种质间无明显差异。高温胁迫使金荞麦种质的光化学量子产量减小。其中种质JQ3、JQ2 和JQ6 降低幅度较小,在各处理温度梯度下差异均未达到显著水平,其余种质的光化学量子产量均在35 ℃处理时与对照间存在显著差异(P <0.05)。随处理温度进一步增加至45 ℃时,JQ5、JQ4 和JQ7 的光化学量子产量迅速降低,分别为0.354、0.362 和0.381,低于其余种质;种质JQ1 和JQ8 居中。由此可见,种质JQ3、JQ2 和JQ6 的叶片实际光化学量子产量受到的影响较小,种质JQ5、JQ4 和JQ7 受到的影响较大,种质JQ1 和JQ8 受到的影响居中。

图2 高温胁迫对金荞麦PSⅡ潜在活性、最大光化学效率的影响Fig.2 Effects of high temperature stress on potential activity,maximum photochemical efficiency of Fagopyrum dibotrys

图3 高温胁迫对金荞麦实际化学量子产量的影响Fig.3 Effects of high temperature stress on actual photochemical quantum yield of Fagopyrum dibotrys
2.2.4 光化学猝灭系数 对高温胁迫下金荞麦叶片光化学猝灭系数(qP)的测定分析(图4)表明,22℃处理时,8 个金荞麦种质叶片qP值介于0.723 ~0.829,种质间无明显差异。受到高温胁迫后,各种质的qP均呈降低趋势。35 ℃处理时,种质JQ3、JQ2和JQ6 的qP与对照间无显著差异(P >0.05),其余种质的qP均显著低于对照(P <0.05)。温度进一步增加到45 ℃处理时,各种质的qP均显著低于对照,其中种质JQ3、JQ2 和JQ6 的qP降幅较小,分别为0.742、0.703 和0.689;种质JQ5、JQ4 和JQ7 的qP降低幅度较大,分别为0. 424、0. 447 和0. 440。JQ1 和JQ8 降低幅度居中。由此可知,种质JQ3、JQ2和JQ6 的qP受高温影响晚且小;种质JQ5、JQ4 和JQ7 受高温影响较大,种质JQ1 和JQ8 受高温影响居中。
2.2.5 非光化学猝灭系数 对高温胁迫对金荞麦叶片非光化学淬灭系数(NPQ)的测定分析(图4)表明,22 ℃处理时,种质间NPQ 差异不明显(P >0.05)。35 ℃处理时,各种质的NPQ 均显著增加(P <0.05);到45 ℃时,种质JQ1、JQ2、JQ3、JQ6 和JQ8 的NPQ 继续增加,达到最大值,其中JQ6 和JQ2的NPQ 提高幅度较大,分别为4.393 和4.291,均显著高于35 ℃处理,其余种质均与35 ℃处理时无显著差异。到45 ℃时,种质JQ4、JQ5 和JQ7 的NPQ则降低,种质JQ7 和JQ5 显著低于35 ℃处理,分别为1.896 和2.469,与对照间无显著差异。由此可知,高温胁迫下,各金荞麦种质通过增加热耗散消耗了过剩的激发能,其中种质JQ1、JQ2、JQ3、JQ6 和JQ8 的热耗散能力较强,特别是种质JQ6 和JQ2,而种质JQ4、JQ5 和JQ7 的热耗散能力较弱,当温度增加到一定程度时(45 ℃),其叶片的光合机构受损,表现为NPQ 降低。
2.3 光合参数的主成分分析
利用主成分分析,将9 个相互联系的单项指标转换成9 个相互独立的综合指标(表2)。其中前两个综合指标累计贡献率为93.068%,大于85%,两个综合指标反映了原始单项指标的绝大部分信息,可以选择第1、2 个综合指标代替原来9 个单项指标综合评价8 个金荞麦种质的耐热性。根据各综合指标的特征向量值,可得到两个综合指标与原来9 个单项指标的线性组合方程。

图4 高温胁迫对金荞麦光化学猝灭系数和非光化学淬灭系数的影响Fig.4 Effects of high temperature stress on photochemical fluorescence quenching and non-photochemical fluorescence quenching of Fagopyrum dibotrys
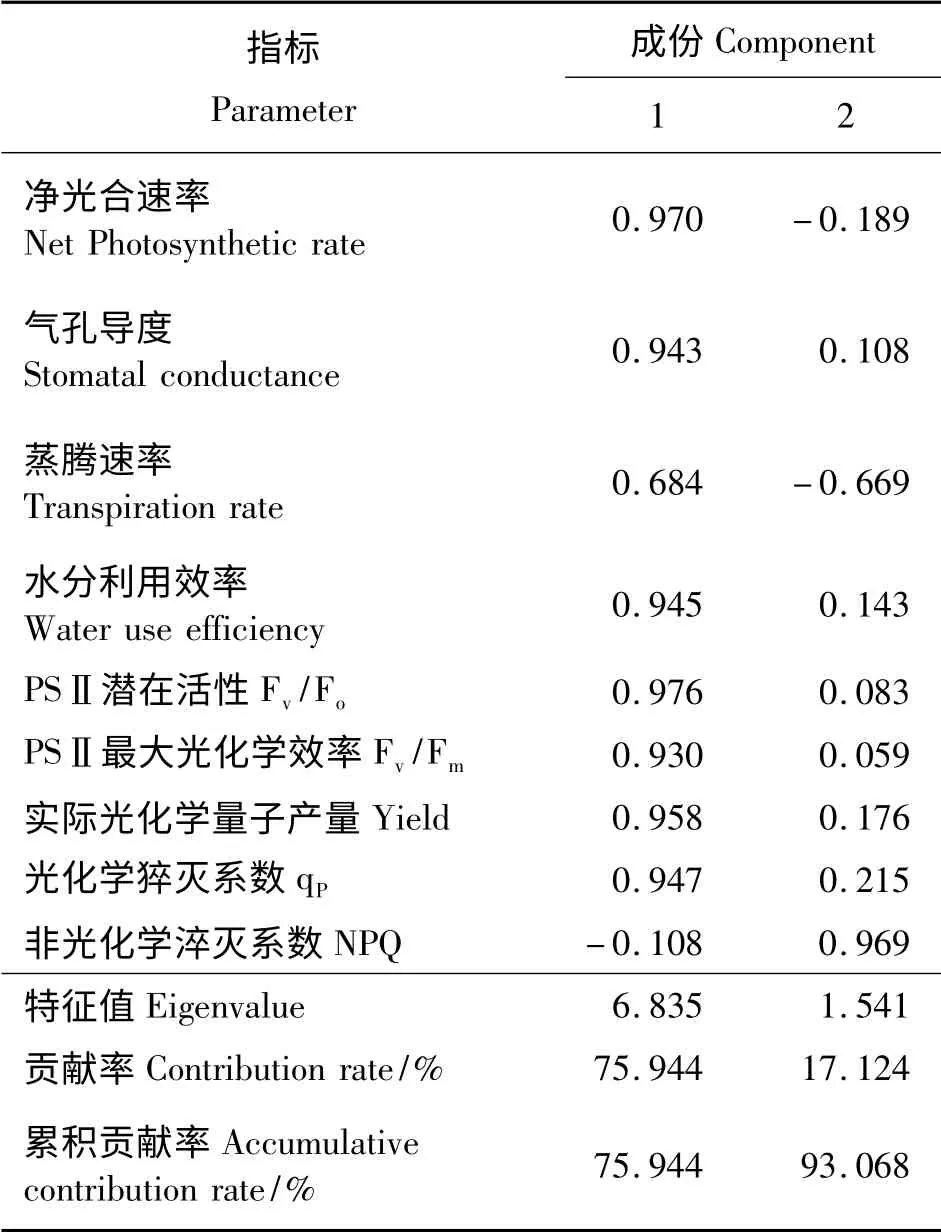
表2 高温胁迫下金荞麦各指标的主成分分析Table 2 Principal component analysis of different indexes of Fagopyrum dibotrys under high temperature stress
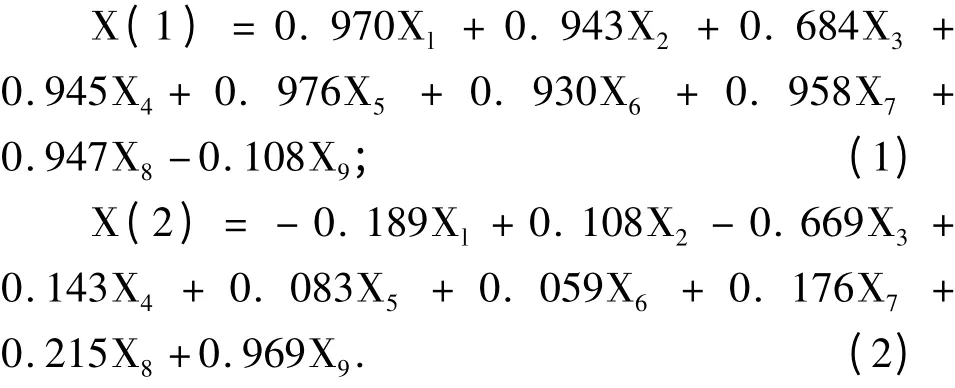
根据各综合指标的贡献率及累计贡献率,利用公式(2)分别求出各综合指标的权重。经计算,两个综合指标的权重分别为0.816 和0.184。
2.4 耐热性综合评价
高温胁迫下,8 个金荞麦种质上述光合参数的反应不同,为更好地评价不同种质幼苗的耐热性差异,在考虑各综合指标的权重的基础上,根据隶属函数公式和耐热性综合评价公式计算出反映供试材料耐热性强弱的综合耐热D 值(表3),结果表明,8 个金荞麦种质资源的耐热性大小表现为JQ6 >JQ2 >JQ3 >JQ1 >JQ8 >JQ4 >JQ5 >JQ7。

表3 高温胁迫下金荞麦综合指标值(xi)、隶属函数值U(xi)、综合评价值(D)和排序Table 3 The values of comprehensive index xi and subordinate function value U(xi),comprehensive evaluation value D and the rank of Fagopyrum dibotrys under high temperature stress
3 讨论与结论
3.1 高温对气体交换参数的影响
当植物遭受高温胁迫且胁迫强度逐渐增加时,首先光合碳同化过程中的Rubisco 活性降低,PSⅡ、细胞色素复合体和类囊体膜功能受到影响,随后类囊体膜结构遭到破坏,电子传递紊乱,甚至导致细胞、叶片乃至植物的死亡[11]。气孔是植物水分向外蒸腾的器官,又是光合作用CO2进入细胞的门户,所以气孔导度的大小既影响蒸腾速率,又影响光合速率和水分利用效率[12]。Berry 等[13]的研究认为,高温对光合作用的抑制主要是降低气孔导度,使叶绿体内CO2的供应受阻。本研究中,金荞麦种质受到高温胁迫后,气孔导度、净光合速率、蒸腾速率和水分利用效率均降低,利用主成分分析和隶属函数综合评价出耐热性强的种质JQ6 和JQ2,气孔导度、净光合速率和水分利用效率降低幅度较小,蒸腾速率降低幅度较大,说明耐热性强的种质通过气孔调节,避免水分过度散失,维持较高水分利用效率,并保证光合机构碳同化的最大运转,合成需要的同化产物[14],即以最小的水分蒸腾量获得最大的干物质积累。
3.2 高温对叶绿素荧光参数的影响
植物对光能的利用主要包括光化学反应转化光能、非光化学热耗散以及以叶绿素荧光形式耗散的过剩光能[15]。PSⅡ的潜在活性(Fv/Fo)表示光反应中心潜在光化学效率[16]。最大光化学效率(Fv/Fm)反映PSⅡ利用光能的能力,非逆境条件下植物的Fv/Fm一般为0.75 ~0.85,在逆境条件下或受到伤害时Fv/Fm会显著降低[17]。实际光化学量子效率(Yield)反映PSⅡ反应中心在部分关闭状态下的实际原初光能捕获效率[18]。光化学猝灭系数(qP)代表PSⅡ反应中心的开放比例,qP越高,PSⅡ反应中心的电荷分离和电子传递能力越强[11]。受到高温胁迫后,金荞麦各种质的上述指标均降低,说明高温胁迫影响了金荞麦PSⅡ潜在活性,降低了光合作用的电子传递,使光合电子由PSⅡ反应中心向电子受体A、B 及质体醌库传递过程受阻[19]。非光化学淬灭(NPQ)是表示热耗散多少的指标,其值越高表明植物将过剩的激发能以热耗散形式释放掉的能力越强,进而降低了过剩光能对PSⅡ反应中心的损伤,缓解了环境对光合作用的影响[20-21]。本研究中,随高温胁迫强度的增加,金荞麦种质的NPQ 在35 ℃时显著增加,45 ℃时综合评价耐热性强的种质JQ6和JQ2 继续显著增加,耐热性弱的种质JQ5 和JQ7显著降低。说明在高温胁迫下,金荞麦种质为保护光合机构,将PSⅡ反应中心接受的多余激发能以热形式耗散;耐热性强的种质的热耗散能力强于耐热性弱的种质。
3.3 耐热性综合评价
史彦江等[11]、赵龙飞等[21]和陈国清等[22]等分别研究了新疆榛(Corylus heterophylla ×C. avellan)、灌浆结实期糯玉米(Zea mays)以及花期前后不同基因型玉米的光合特性,研究结果表明,植物在高温胁迫下,叶片气体交换参数和荧光参数降低,耐热性强的材料下降幅度小于耐热性弱的材料。本研究中,随高温胁迫强度的增强,金荞麦种质的Pn、Gs、Tr、WUE、Fv/Fo、Fv/Fm、Yield 和qP均降低,NPQ 先升高后降低或一直升高,但种质材料间各单项指标的变化存在差异,因此利用单项指标评价种质间的耐热性顺序结果不一致。而且有研究指出植物耐热性属于数量性状,受多个基因的控制,仅凭个别或少数指标的简单比较难以真实地反映植物耐热性的遗传本质[23]。本研究利用主成分分析,在较少损失原有信息的前提下,将9 个彼此相关的光合和叶绿素荧光参数简化为9 个相互独立的综合指标,其中前两个指标的累计贡献率为93.068%,大于85%,确定主成分个数为2。并利用综合指标的贡献率和累计贡献率计算出两个综合指标的权重分别为0.816 和0.184,最后在考虑各综合指标权重的基础上,通过隶属函数综合评价出8 个金荞麦种质的耐热性。该方法既考虑了各单项指标间的相互关系,又兼顾了各指标的重要性,也避免了人为确定权重的主观性[24]。
本研究中,高温胁迫降低了金荞麦种质的Pn、Gs、Tr、WUE、Fv/Fo、Fv/Fm、Yield 和qP。为抵御高温,耐热性强的种质通过调节气孔导度,减小蒸腾速率,提高水分利用率,并将过剩的激发能以热的形式耗散掉,以保护光合机构。利用主成分分析和隶属函数法,综合评价出8 个金荞麦种质的耐热性由强到弱的顺序为:JQ6、JQ2、JQ3、JQ1、JQ8、JQ4、JQ5、JQ7。
[1] 周宇飞,王德权,陆樟镳,王娜,王艺陶,李丰先,许文娟,黄瑞冬.干旱胁迫对持绿性高粱光合特性和内源激素ABA、CTK含量的影响[J].中国农业科学,2014,47(4):655-663.
[2] 张守仁.叶绿素荧光动力学参数的意义及讨论[J].植物学通报,1999,16(4):444-448.
[3] Maxwell K,Johnson G N.ChlorophyⅡfluorescence——A practical guide[J].Journal of Experimental Botany,2000,51(345):659-668.
[4] Angelp M M,Adelaide L,Fabrizio P,Francesco L.The effect of growth at low temperature on photosynthetic characteristics and mechanisms of photoprotection of maize leaves[J].Journal of Experimental Botany,1995,46(1):119-127.
[5] 张向峰,王玉杰,李云霞,王云琦,张会兰,刘敏.模拟酸雨对盆栽马尾松叶片光合特性的影响[J].水土保持学报,2013,27(1):197-202.
[6] 郝婷,朱月林,丁小涛,金海军,张红梅,余纪柱.根际高温胁迫对5 种瓜类作物生长及叶片光合和叶绿素荧光参数的影响[J].植物资源与环境学报,2014,23(2):65-73.
[7] 贾彩凤,李艾莲.药用植物金荞麦的光合特性研究[J].中国中药杂志,2008,33(2):129-132.
[8] 尹迪信,唐华彬,罗红军,郑汝权,郭克祥,郑蓉.野生牧草金荞麦与贵州省推广牧草栽培效益比较试验初报[J].草业科学,2006,23(7):45-48.
[9] 王凯红,凌家慧,张乐华,刘向平,李立.两种常绿杜鹃亚属幼苗耐热性的主成分及隶属函数分析[J].热带亚热带植物学报,2011,19(5):412-418.
[10] 王凯红,刘向平,张乐华,凌家慧,李立.5 种杜鹃幼苗对高温胁迫的生理生化响应及耐热性综合评价[J].植物资源与环境学报,2011,20(3):29-35.
[11] 史彦江,罗青红,宋锋惠,俞涛,寇云玲. 高温胁迫对新疆榛光合参数和叶绿素荧光特性的影响[J]. 应用生态学报,2012,23(9):2477-2482.
[12] 陈托兄,郝文军,陈小兵,卢欣石.10 个紫花苜蓿品种光合特性的比较[J].中国草地学报,2009,31(2):41-45.
[13] Berry J,Bjǒrkman O.Photosynthetic response and adaptation to temperature in higher plants[J].Annual Review of Plant Physiology,1980,31:491-543.
[14] 王刚,孙广玉.不同紫花苜蓿品种光合能力的比较[J].东北林业大学学报,2007,35(4):19-21.
[15] 杜尧东,李键陵,王华,唐湘如,胡飞. 高温胁迫对水稻剑叶光合和叶绿素荧光特征的影响[J]. 生态学杂志,2012,31(10):2541-2548.
[16] 刘志梅,蒋文伟,杨广远,黄建荣.干旱胁迫对3 种金银花叶绿素荧光参数的影响[J].浙江农林大学学报,2012,29(4):533-539.
[17] 张金政,张起源,孙国峰,何卿,李晓东,刘洪章.干旱胁迫及复水对玉簪生长和光合作用的影响[J]. 草业学报,2014,23(1):167-176.
[18] 卢广超,许建新,薛立,刘斌,李诗刚,邵怡若.干旱胁迫下4 种常用植物幼苗的光合和荧光特性综合评价[J].生态学报,2013,33(24):7872-7881.
[19] 陈兵,王克如,李少昆,金秀良,陈江鲁,张东升.病害胁迫对棉叶光谱反射率和叶绿素荧光特性的影响[J].农业工程学报,2011,27(9):86-93.
[20] 赵丽丽,王普昶,陈超,刘洪来,马林.干旱胁迫对百脉根叶绿素荧光特性的影响[J]. 中国草地学报,2014,36(1):18-23.
[21] 赵龙飞,李潮海,刘天学,王秀萍,僧珊珊.花期前后高温对不同基因型玉米光合特性及产量和品质的影响[J].中国农业科学,2012,45(23):4947-4958.
[22] 陈国清,沈鑫,蔡雪梅,陆卫平,陆大雷.灌浆结实期高温胁迫对糯玉米子粒产量和叶片光合特性的影响[J].玉米科学,2014,22(2):104-108.
[23] 张朝阳,许桂芳.利用隶属函数法对4 种地被植物的耐热性综合评价[J].草业科学,2009,26(2):57-60.
[24] 赵丽丽,王普昶,陈超,马林.高温胁迫下百脉根生理生化响应及耐热性评价[J].草业科学,2013,30(12):2018-2023.
