柔嫩艾美耳球虫5-磷酸脱氧木酮糖还原异构酶基因的克隆、原核表达及酶活性分析
2015-03-22廖申权吴彩艳戚南山吕敏娜张健騑谢明权孙铭飞
廖申权,吴彩艳,戚南山,李 娟,吕敏娜,张健騑,谢明权,孙铭飞
(广东省农业科学院动物卫生研究所,广东省畜禽疫病防治研究重点实验室,广东省兽医公共卫生公共实验室,广州 510640)
柔嫩艾美耳球虫5-磷酸脱氧木酮糖还原异构酶基因的克隆、原核表达及酶活性分析
廖申权,吴彩艳,戚南山,李 娟,吕敏娜,张健騑,谢明权,孙铭飞*
(广东省农业科学院动物卫生研究所,广东省畜禽疫病防治研究重点实验室,广东省兽医公共卫生公共实验室,广州 510640)
旨在克隆柔嫩艾美耳球虫5-磷酸脱氧木酮糖还原异构酶(EtDXR)基因,初步分析EtDXR的酶活性。根据EuPathDB数据库信息注释、拼接Etdxr基因序列,设计特异性引物扩增Etdxr基因,构建原核表达质粒pCold I-EtDXR,转化E.coliRosetta(DE3),IPTG诱导表达,进行SDS-PAGE及Western blot分析,纯化rEtDXR,对rEtDXR进行酶活性分析,测定pH和Mg2+离子浓度对酶活性的影响。结果显示,Etdxr基因完整编码序列为1 746 bp,氨基酸序列具有与其他物种高度保守的酶活性中心;成功构建原核表达质粒pCold I-EtDXR,SDS-PAGE以及Western blot分析显示,得到约50 ku的蛋白质;利用直接检测底物NADPH 消耗速率的方法分析EtDXR酶活性,结果显示,不同pH和离子浓度对EtDXR的酶活性有显著影响,其最佳的酶反应条件为pH 7.0,Mg2+离子浓度4.5 mmol·L-1。本研究为进一步以EtDXR为药靶筛选抗球虫先导化合物奠定了基础。
柔嫩艾美耳球虫;5-磷酸脱氧木酮糖还原异构酶;克隆;原核表达;酶活性
柔嫩艾美耳球虫(E.tenella)是一种细胞内寄生的顶复门原虫,它引起的鸡球虫病对养禽业危害极大。鸡球虫病传统的控制策略依赖于抗球虫药物的应用,但随着球虫耐药性的产生与持续增加,使得药物防治鸡球虫病举步维艰,新的抗球虫药物的研制已成为生产实际的迫切需求。
顶质体(apicoplast)是顶复门原虫特有的细胞器,类似植物叶绿体样质体。寄生原虫的顶质体内存在一些特殊且必需的代谢途径,参与寄生原虫脂肪酸、血红素及类异戊二烯的生物合成,对寄生原虫的生存、发育起着十分重要的作用[1]。这类特殊代谢途径及其关键酶为抗寄生虫药物的研发提供了潜在药靶。5-磷酸脱氧木酮糖还原异构酶(1-deoxy-D-xylulose-5-phosphate reductoisomerase,DXR)是类异戊二烯合成途径关键酶,已成为抗寄生原虫药物研发的关键靶标[2]。以DXR为靶标筛选的抑制剂膦胺霉素成功用于抗恶性疟原虫的临床试验[3]。本研究对柔嫩艾美耳球虫5-磷酸脱氧木酮糖还原异构酶进行基因克隆、原核表达、分离纯化,并初步对rEtDXR的酶活性进行分析,为深入研究EtDXR的生物学特性奠定基础。
1 材料与方法
1.1 虫株、菌株、质粒和主要试剂
E.tenella(GD株)、E.coliDH5α、E.coliRosetta(DE3)由广东省农业科学院动物卫生研究所寄生生物室保存;pMD18-T 载体购自宝生物(大连)有限公司;pCold I表达载体由南京农业大学茅翔教授馈赠。LATaqDNA 聚合酶、DNA Marker、BamHⅠ、HindⅢ、RT-PCR试剂盒购自TaKaRa公司;TRIzol 购自Invitrogen公司;E.Z.N.A质粒小量制备试剂盒和DNA胶回收试剂盒购自Omega公司;Ni-NTA Resin 购自Novagen公司;1-脱氧-D-木酮糖-5-磷酸(1-deoxy-D-xylulose-5-phosphate,DXP),辅酶NADPH购自Sigma公司。
1.2 基因注释
利用EuPathDB数据库(http://eupathdb.org)信息,查找寄生原虫,恶性疟原虫(Plasmodiumfalciparum)、刚地弓形虫(Toxoplasmagondii)、牛巴贝斯虫(Babesiabovis)、环形泰勒虫(Theileriaannulata)等类异戊二烯合成途径关键酶DXR的基因序列。利用在线多重比对工具ClustaW2(http://www.ebi.ac.uk/Tools/msa/clustalw2/)分析顶复门原虫dxr的保守序列;利用保守的氨基酸序列作为种子序列,搜索柔嫩艾美耳球虫基因组数据库(http://toxodb.org/toxo/),注释、拼接Etdxr的基因序列。分析发现ETH_00017440、ETH_0003428含有dxr的保守序列,注释、拼接Etdxr基因序列。
1.3 裂殖子纯化及总RNA的提取
裂殖子的纯化参照谢氏方法[4]进行。以1×105个孢子化卵囊·只-1经口感染14日龄无球虫感染健康鸡,感染后112 h剖杀试验鸡,取盲肠纯化第二代裂殖子。应用Trizol法提取E.tenella第二代裂殖子总RNA,以该RNA为模板,用一步法RT-PCR试剂盒进行扩增。
1.4Etdxr基因的扩增
根据注释序列设计特异性扩增引物:上游引物,EtdxrF:5′-CGCGGATCCATGAGGCGGCGGTGGTTTGTC-3′ (含BamHⅠ酶切位点);下游引物EtdxrR:5′-CGCAAGCTTTCAGGGGCCCCCTTC-CCCC-3′ (含Hind Ⅲ酶切位点)。RT-PCR扩增目的基因Etdxr,回收目的片段PCR产物,连接T载体,经转化、鉴定,送阳性克隆测序。
1.5Etdxr基因序列分析
利用在线分析工具ClustalW(http://www.ebi.ac.uk/Tools/msa/clustalw2/),将EtDXR与其他物种DXR氨基酸序列进行多重比对分析,分析预测转导序列及酶活性位点。
1.6 密码子优化
利用http://gcua.schoedl.de/index分析,选用E.coli偏好性密码子,对EtDXR 141-224位(即成熟蛋白质1—84位)氨基酸密码子进行优化,由上海英潍捷基贸易有限公司全基因合成,并运用重叠延伸PCR方法重叠合成改造后的基因,与T载体连接,转化E.coliDH5α,筛选阳性克隆进行测序验证。
1.7 表达载体构建
用限制性内切酶BamHⅠ、HindⅢ双酶切质粒pMD18T-Etdxr和载体pCold I,切胶回收后经T4 DNA Ligase连接,构建pCold I-Etdxr表达载体,转化E.coliDH5α,筛选阳性克隆进行测序验证。
1.8 重组质粒在E.Coli中的表达及纯化
将测序正确的pCold I-Etdxr转入E.ColiRosetta (DE3),筛选阳性单克隆在10 mL LB(含Amp)培养基中培养10 h,按1∶100扩大培养于LB(含Amp)培养基中,37 ℃培养至OD600 nm为0.4~0.6时,0.5 mmol·L-1IPTG,22 ℃过夜诱导表达,SDS-PAGE分析表达产物,以Ni-NTA Resin纯化目的蛋白质,具体参照M.Sun等的方法[5]。
1.9 重组蛋白质的Western blot检测
参照《分子克隆实验指南》的方法进行。将纯化的重组蛋白质进行SDS-PAGE分析,再将SDS-PAGE中的蛋白质以电转仪转至NC膜上,以封闭液(PBST + 5% BSA)封闭2 h,与鼠抗His-tag抗体室温反应2 h,以PBST 洗膜3次,加入辣根过氧化物酶标记的羊抗鼠IgG二抗(1∶5 000),室温反应2 h,洗膜3次,以HPR/DAB显色试剂盒避光显色。
1.10 重组EtDXR酶活性的测定
EtDXR催化DXP转化为2C-甲基-D-赤藓糖醇-4-磷酸(2C-methyl-D-erythritol-4-phosphate,MEP)。DXR 的催化活性需要NADPH作为辅酶及二价金属离子(Mg2+、Mn2+或 Co2+)作为辅离子。本研究采用直接检测底物消耗速率的方法,以Varioskan Flash 全波长扫描式多功能读数仪在激发波长340 nm和发射波长460 nm测定的相对荧光值(RFU)检测NADPH降解速率,测定该酶的酶活性[6]。反应体系:100 mmol·L-1MOPS、4.5 mmol·L-1MgCl2、0.3 mmol·L-1DXP、0.15 mmol·L-1NADPH、10 μg·mL-1EtDXR,反应溶液于30 ℃孵育5 min,加入EtDXR开始反应。
为分析pH和Mg2+离子浓度对EtDXR酶活性的影响,本研究采用响应曲面法(response surface methodology,RSM)确定酶最适反应条件,以SAS/Statistic分析酶的最佳反应条件。试验设pH和离子浓度2个因素,具体反应设计见表1。
表1 影响因素的试验设计
Table 1 Range of factors used in the experimental design

影响因素Factor试验因素的水平Leveloffactor-1.41-1011.41pH4.175799.83I/(mmol·L-1)014.589.45
2 结 果
2.1Etdxr基因的注释与PCR扩增
利用EuPathDB数据库信息,进行生物信息学分析,注释、拼接Etdxr完整编码序列1 746 bp。以TRIzol方法提取第二代裂殖子总RNA,根据预测基因片段设计的特异性引物,以第二代裂殖体总RNA为模板,进行RT-PCR扩增,1%琼脂糖凝胶电泳分析显示,在1 800 bp处有目的条带,与预期片段大小一致(图1);将PCR产物与pMD18-T载体连接,筛选阳性克隆送测序,测序结果显示所扩增目的片段大小为1 746 bp。

M.DL2000 DNA相对分子质量标准;1、2.RT-PCR扩增产物M.DL2000 DNA marker;1,2.RT-PCR products of Etdxr图1 Etdxr基因的RT-PCR扩增产物Fig.1 RT-PCR amplification of Etdxr gene
2.2Etdxr基因序列分析
利用在线分析工具ClustalW将EtDXR与其他物种DXR氨基酸序列进行多重比对分析,结果显示,顶复门原虫均具有保守的酶活性位点,如图2。经比对分析,推测E.tenelladxr存在转导肽序列(1—140位氨基酸),与顶复门的T.gondii、P.falciparumdxr基因类似,存在转导肽序列,其蛋白质在细胞质中合成后经转运靶向虫体的顶质体。
2.3 密码子优化基因的原核表达与鉴定
利用重叠延伸PCR方法重叠全基因合成的EtDXR(141—224)及EtDXR(225-581),构建pCold I-EtDXR表达质粒,经测序分析正确后,将pCold I-EtDXR转化E.ColiRosetta (DE3),鉴定为阳性的E.coliRosetta (DE3) (pCold I-EtDXR)以0.5 mmol·L-1IPTG,22 ℃低温诱导表达,收集菌体,重组表达产物进行SDS-PAGE分析,见图3A。以PBS重悬菌体,冰浴超声破碎,离心取上清过亲和层析柱,梯度洗脱,250 mmol·L-1咪唑洗涤,获得较纯的His-EtDXR融合蛋白质,约50 ku(图3B)。将纯化后的融合蛋白质进行SDS-PAGE分析,切胶转膜至NC膜上,封闭液封闭2 h,鼠抗His标签一抗孵育2 h,HRP标记羊抗鼠二抗孵育2 h,显色,Western blot结果显示纯化获得目的蛋白质,见图3C。

下划线标记为保守的酶活性位点Conserved domains are highlighted in underline图2 E.tenella DXR与其他物种DXR的多重比对结果Fig.2 Alignment of the amino acid sequences of E.tenella DXR with other DXR sequences
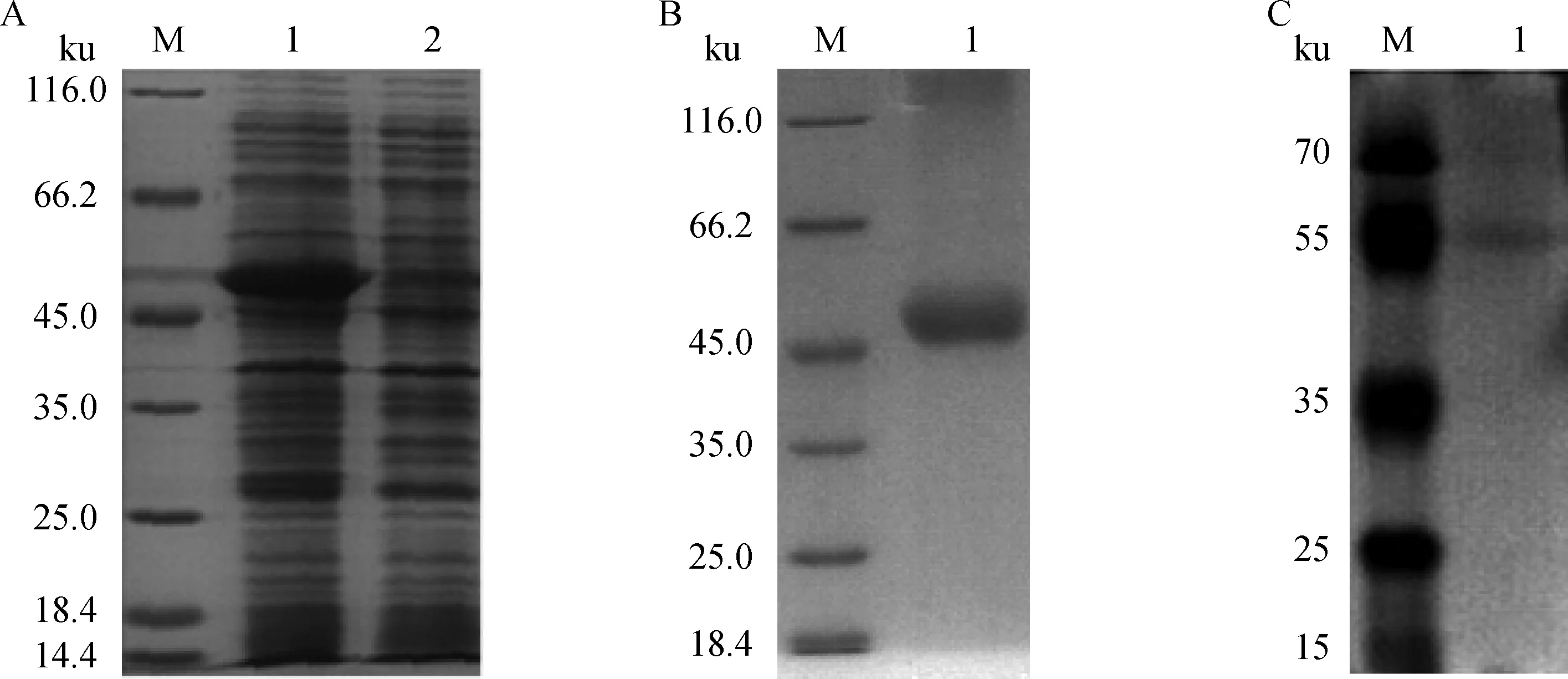
M.蛋白质相对分子质量标准;A1.诱导的pCold I-EtDXR/ Rosetta (DE3);A2.未诱导的pCold I-EtDXR/ Rosetta (DE3);B1.纯化的His-EtDXR;C1.纯化的His-EtDXRM.Protein marker;A1.Induced pCold I-EtDXR/ Rosetta (DE3);A2.Non-induced pCold I-EtDXR/ Rosetta (DE3);B1.Purified His-EtDXR;C1.Purified His-EtDXR图3 重组蛋白质的表达及分析Fig.3 SDS-PAGE and Western blot analysis of recombinant EtDXR
2.4 重组EtDXR的酶活性分析
利用生物化学的方法对重组EtDXR进行了酶活性测定,分析了pH和Mg2+离子浓度对其酶活性的影响。结果显示,重组EtDXR可以利用底物1-脱氧-D-木酮糖-5-磷酸与NADPH,具有5-磷酸脱氧木酮糖还原异构酶的催化活性。响应曲面法分析结果显示,不同pH和离子浓度对大肠杆菌表达EtDXR的酶活性有显著影响,其酶动力学反应的最佳条件为pH 7.0,Mg2+离子浓度4.5 mmol·L-1,见图4。

图4 离子浓度及pH对EtDXR酶活性的影响Fig.4 Effects of ion concentration and reaction pH on enzymatic activity
3 讨 论
类异戊二烯及其衍生物构成自然界极为多样性的化合物,这些化合物是泛醌、多萜醇的前体。类异戊二烯的衍生物泛醌参与电子传递,长醇参与蛋白的糖基化,蛋白的法尼基化和牻牛儿基化对原虫的生存是必需的[7]。早期研究认为类异戊二烯通过甲羟戊酸途径(the mevalonic acid pathway,MVA)合成。直至1993年,报道真细菌利用甲基赤藻糖醇途径(the methylerythritol pathway,MEP)合成类异戊二烯。研究显示,顶复门原虫(如Plasmodiumspp.、Trypanosomaspp.、T.gondii、Eimeriaspp.及Babesiaspp.)等[8-9]仅利用MEP途径合成类异戊二烯;而宿主则利用MVA途径合成类异戊二烯,MEP途径的关键酶已成为备受关注的药物靶标。5-磷酸脱氧木酮糖还原异构酶(DXR)是MEP途径的关键酶,催化1-脱氧-D-木酮糖-5-磷酸转化为2C-甲基-D-赤藓糖醇-4-磷酸(MEP)。由于1-脱氧-D-木酮糖-5-磷酸可以合成硫胺素焦磷酸和磷酸吡哆醇,因此,MEP是类异戊二烯生物合成途径中真正意义上的关键中间体,DXR催化的反应是MEP生物合成途径“碳流”的分支点,也是进行调控的有效靶点,成为研究抗感染性药物的重要靶标[10-11]。
目前仍未见柔嫩艾美耳球虫dxr基因序列及功能研究的报道,本研究利用生物信息学预测并注释Etdxr基因,克隆获得Etdxr完整编码序列。结果显示,扩增的序列与预测序列完全一致;不同物种间多重比对发现,EtDXR与其他物种DXR存在相同的酶活性位点;T.gondii、P.falciparum及E.tenellaDXR都存在转导肽序列,蛋白质在细胞质中合成后经转运靶向顶质体,在顶质体内发挥生物学功能。
在前期试验过程中,尝试将未优化的Etdxr更换不同的原核表达载体及优化原核表达条件,均未发现Etdxr在大肠杆菌中的表达。Etdxr序列的GC含量非常高,达到67.87%,可能导致Etdxr在大肠杆菌中难以表达。研究表明,影响外源基因表达效率的主要因素为密码子选用、mRNA稳定性、翻译起始效率和载体选择等。其中密码子偏爱性极大地影响外源蛋白质的表达量,研究表明按照宿主细胞的偏爱性将目的基因编码区全部进行优化,将使表达量提高10~50倍,即使仅将5′端编码区进行改造或者改变其中的稀有密码子,也可大幅度提高蛋白质的表达量[12]。本研究采用优化5′端编码区的方法,优化成熟蛋白质的前84个氨基酸,优化后Etdxr序列GC含量为58.81%。试验证实优化后的序列在大肠杆菌中能有效表达,经纯化、鉴定,获得可溶性目的蛋白质。
对EtDXR的生化分析显示,原核表达的EtDXR具有酶活性。利用响应曲面法分析发现,EtDXR的酶活性受pH和Mg2+离子浓度的影响,其酶动力学反应的最佳条件为pH 7.0,Mg2+离子浓度4.5 mmol·L-1,与J.Cheleski等[13]的报道相似。基于对大肠杆菌、结核分枝杆菌、鼠疫耶尔森菌等重要致病菌DXR结构信息学的解析,已经筛选出具有抗菌活性的先导化合物[14-15],为顶复门原虫DXR抑制剂的设计及筛选提供了试验依据。本研究初步分析了柔嫩艾美耳球虫5-磷酸脱氧木酮糖还原异构酶的酶活性,并筛选到最佳酶反应条件,为进一步以EtDXR为药靶筛选抗球虫先导化合物奠定基础。
[1] SATO S.The apicomplexan plastid and its evolution [J].CellMolLifeSci,2011,68(8):1285-1296.
[2] GOBLE J L,ADENDORFF M R,DE BEER T A,et al.The malarial drug targetPlasmodiumfalciparum1-deoxy-D-xylulose-5-phosphate reductoisomerase (PfDXR):development of a 3-D model for identification of novel,structural and functional features and for inhibitor screening [J].ProteinPeptLett,2010,17(1):109-120.
[3] BORRMANN S,LUNDGREN I,OYAKHIROME S,et al.Fosmidomycin plus clindamycin for treatment of pediatric patients aged 1 to 14 years withPlasmodiumfalciparummalaria [J].AntimicrobAgentsChemother,2006,50(8):2713-2718.
[4] XIE M Q,GILBERT J M,FULLER A L,et al.A new method for purification ofEimeriatenellamerozoites [J].ParasitolRes,1990,76(7):566-569.
[5] SUN M,ZHU G,QIN Z,et al.Functional characterizations of malonyl-CoA:acyl carrier protein transacylase (MCAT) inEimeriatenella[J].MolBiochemParasitol,2012,184(1):20-28.
[6] DHIMAN R K,SCHAEFFER M L,BAILEY A M,et al.1-Deoxy-D-xylulose 5-phosphate reductoisomerase (IspC) fromMycobacteriumtuberculosis:towards understanding mycobacterial resistance to fosmidomycin [J].JBacteriol,2005,187(24):8395-8402.
[7] SEEBER F,SOLDATI-FAVRE D.Metabolic pathways in the apicoplast of apicomplexa [J].IntRevCellMolBiol,2010,281:161-228.
[8] JOMAA H,WIESNER J,SANDERBRAND S,et al.Inhibitors of the nonmevalonate pathway of isoprenoid biosynthesis as antimalarial drugs [J].Science,1999,285(5433):1573-1576.
[9] ROOS D S.The apicoplast as a potential therapeutic target inToxoplasmaand other apicomplexan parasites:some additional thoughts [J].ParasitolToday,1999,15(1):41.
[10] DE RUYCK J,WOUTERS J,POULTER C D.Inhibition studies on enzymes involved in isoprenoid biosynthesis:Focus on two potential drug targets:DXR and IDI-2 enzymes [J].CurrEnzymInhib,2011,7(2):79-95.
[11] BEHRENDT C T,KUNFERMANN A,ILLARIONOVA V,et al.Reverse fosmidomycin derivatives against the antimalarial drug target IspC (Dxr) [J].JMedChem,2011,54(19):6796-6802.
[12] YADAVA A,OCKENHOUSE C F.Effect of codon optimization on expression levels of a functionally folded malaria vaccine candidate in prokaryotic and eukaryotic expression systems [J].InfectImmun,2003,71(9):4961-4969.
[13] CHELESKI J,FREITAS R F,WIGGERS H J,et al.Expression,purification and kinetic characterization of His-tagged glyceraldehyde-3-phosphate dehydrogenase fromTrypanosomacruzi[J].ProteinExpresPurif,2011,76:190-196.
[14] UMEDA T,TANAKA N,KUSAKABE Y,et al.Molecular basis of fosmidomycin′s action on the human malaria parasitePlasmodiumfalciparum[J].SciRep,2011,1:9.
[15] JACKSON E R,DOWD C S.Inhibition of 1-deoxy-D-xylulose-5-phosphate reductoisomerase (Dxr):a review of the synthesis and biological evaluation of recent inhibitors [J].CurrTopMedChem,2012,12(7):706-728.
(编辑 白永平)
Cloning,Prokaryotic Expression and Enzymatic Activity of 1-deoxy-D-xylulose-5-phosphate Reductoisomerase ofE.tenella
LIAO Shen-quan,WU Cai-yan,QI Nan-shan,LI Juan,LÜ Min-na,ZHANG Jian-fei,XIE Ming-quan,SUN Ming-fei*
(GuangdongLaboratoryforAnimalDiseases,GuangdongOpenLaboratoryofVeterinaryPublicHealth,InstituteofAnimalHealth,GuangdongAcademyofAgriculturalSciences,Guangzhou510640,China)
The aim of this study was to clone 1-deoxy-D-xylulose-5-phosphate reductoisomerase (DXR) gene inEimeriatenella,and analyze its enzymatic activity.The predicted sequences ofEtdxrwere obtained from EuPathDB by bioinformatics analysis.The full-length cDNA was amplified by PCR and then cloned into pCold I vector.The recombinant plasmid was transformed intoE.coliRosetta (DE3) and induced by IPTG.The expression products were analyzed by SDS-PAGE and Western blot,and then were purified.We further evaluated the enzymatic activity of EtDXR by monitoring the consumption of NADPH.The results showed that the open reading frame ofE.tenellaDXR was 1 746 bp.Further analysis of the amino acid sequence revealed that EtDXR contains the conserved domains with other DXRs.The results showed that the recombinant vector pCold I-EtDXR was constructed successfully.The SDS-PAGE and Western blot results showed that the purified protein was 50 kDa.The characterizations of reaction conditions used in the experiments,such as pH and ion concentration have significant effects on the enzymatic activity of rEtDXR.According to the response surface plots,the maximum enzymatic activity was pH 7.0 and 4.5 mmol·L-1Mg2+.This study provides a foundation for the further study to select inhibitors ofE.tenellaDXR.
Eimeriatenella;1-deoxy-D-xylulose-5-phosphate reductoisomerase;cloning;prokaryotic expression;enzymatic activity
10.11843/j.issn.0366-6964.2015.04.017
2014-07-28
国家自然科学基金项目 (31201902;31302087;31402186);广州市珠江科技新星项目(2012J2200059);广东省自然科学基金项目(S2013040015220);广东省科技计划项目(2011B050700007);广州市科技计划项目(2014J4100230);广东省农业科学院院长基金(201413)
廖申权(1981-),女,重庆荣昌人,博士,主要从事鸡球虫生化代谢研究,E-mail:lsq6969@163.com
*通信作者:孙铭飞,副研究员,E-mail:smf7810@126.com
S852.723
A
0366-6964(2015)04-0631-06
