不同树莓品种光合特征1)
2015-03-06程云清耿婉婷张会弟刘剑锋赵一欣杨博文
程云清 耿婉婷 张会弟 刘剑锋 赵一欣 杨博文
(吉林师范大学,四平,136000)
责任编辑:任 俐。
树莓属于蔷薇科(Rosaceae)悬钩子属(Rubus)落叶稀常绿灌木、半灌木或多年生匍匐草本植物,营养丰富,还具特别的保健功能,是重要的小浆果类果树之一[1-2]。树莓浆果酸甜爽口,香味浓郁,可鲜食,也可加工成为果汁、饮料、果酒等饮品[3]。树莓适栽地域较为广泛,对土壤无严格要求,栽植当年即可少量挂果,在定植后的第3、第4年即可进入丰产期[4]。近年来,因树莓浆果新颖奇特,在众多水果中独具一格,市场发展前景很好,适当发展树莓产业对于促进林区经济的发展具有重要的作用。中国树莓产业发展非常迅速,近10年来栽培面积迅猛增加,但目前生产上使用的树莓品种多引自国外,有少数自主品种,这些品种的光合特征多不明确,不利于树莓的栽培与生产。本研究以吉林省白山市四季红树莓基地购买栽培的3个国外品种与1个我国吉林省树莓新品种为试材,深入研究其光合响应参数、CO2响应参数及光合效率的日变化特征,以期为东北地区树莓的科学栽培提供理论依据。
1 材料与方法
供试树莓品种为英国品种‘秋福’(‘Autumn Bliss’,QF),美国品种‘哈瑞太兹’(‘Heritage’,HR)、‘秋红’(‘Red Autumn’,QH),我国品种‘四季红’(‘Sijihong’,SJH),每个品种25株,这些2年生苗于2012年5月7日在吉林师范大学生命科学学院实验田进行了定植。实验田土壤类型为黑土类,pH=7.0左右,属中性土,腐殖质和有机质含量较高。株行距为1 m×1 m,进行常规肥水管理。每个树莓品种选择长势较为整齐的6个植株,选枝梢顶端向下约3个节位的最大无病虫功能叶片进行光合参数的测定。
光合指标的测定:在2012年7月中旬与2013年7月中旬的晴天进行。仪器型号为美国LI-COR公司的LI-6400光合作用测定系统。在08:00—11:00,利用仪器自带光曲线与CO2曲线测定程序进行测定。光响应曲线的测定参数设定为CO2摩尔分数380μmol·mol-1,温度25℃,其他参数为默认值。光合有效辐射(PA,R)梯度设定为2 000、1 800、1 500、1 200、900、600、300、200、100、50、25、0μmol·m-2·s-1。参照文献进行光补偿点与饱和点等参数的计算[5-7]。CO2响应曲线的测定参数设定为PA,R900μmol·m-2·s-1,温度25℃,其他参数为默认值。CO2摩尔分数梯度设定为1 800、1 500、1 200、900、600、300、200、100、50、0μmol·mol-1。参照文献进行CO2饱和点与CO2补偿点等参数的计算[6-7]。光合效率日变化的测定在06:00—19:00进行,每小时测定1次。
数据处理:以上测定重复3~6次,利用SAS 8.2统计软件中的Anova过程进行差异的显著性分析。
2 结果与分析
2.1 4个树莓品种叶片光合响应曲线特征
4个树莓品种的光合曲线结果如图1所示。PA,R为0~900μmol·m-2·s-1时,Pn-PA,R曲线大致呈抛物线型,此后,净光合速率随PA,R的增加变化幅度不大,这表明这4个树莓品种并不存在明显的光抑制现象。净光合速率相比较,不同品种间存在一定的差异,由大到小的顺序为SJH、QH、QF、HR。
气孔导度随光合有效辐射变化曲线变化较为平缓,气孔导度随着PA,R的增加大致呈上升趋势。当PA,R为0~300μmol·m-2·s-1时,气孔导度随PA,R上升的增加较为迅速,此后,上升趋缓。对气孔导度与净光合速率进行了相关性分析,HR、QH、QF和SJH的相关系数分别为0.75、0.88、0.97和0.98,气孔导度与净光合速率的相关性均达到显著水平(P<0.05)。这表明树莓气孔的开关受到PA,R的调节,0~300μmol·m-2·s-1PA,R的增加可诱导气孔开放。相同PA,R条件下气孔导度相比较,4个树莓品种由大到小的顺序大致为HR、QH、SJH、QF,这与净光合速率的大小顺序并不一致。
胞间CO2摩尔分数随PA,R的增加呈下降趋势。在PA,R为0~600μmol·m-2·s-1时,随PA,R的增加,胞间CO2摩尔分数下降迅速,此后,胞间CO2摩尔分数变化不明显。相同PA,R条件下胞间CO2摩尔分数相比较,4个树莓品种由大到小的顺序为HR、QH、QF、SJH,这与净光合速率的大小顺序完全不同,与气孔导度的大小顺序则较为接近。
蒸腾速度随PA,R增加大致呈上升趋势。相同PA,R条件下蒸腾速率相比,4个树莓品种由大到小的顺序为SJH、QF、QH、HR,这与净光合速率的大小顺序大致一致。相关性分析表明,4个树莓品种的蒸腾速率与气孔导度间均存在显著正相关关系(P<0.05),其中,HR、QH、QF、SJH的气孔导度与净光合速率间相关系数分别为0.98、0.95、0.99和0.99,相关均达到极显著水平(P<0.01)。
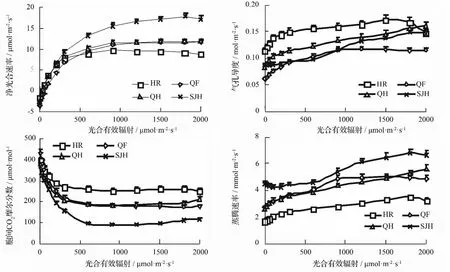
图1 4个树莓品种叶片光合响应曲线特征
4个树莓品种的光合响应参数如表1所示。由 表1可见,4个树莓品种的光补偿点为20.58~72.09 μmol·m-2·s-1,其光饱和点为742.50~887.50μmol·m-2·s-1,光饱和光合速率为9.82~17.73μmol·m-2·s-1,表观量子效率为0.038 1~0.044 5。其中,树莓品种HR光补偿点最低,而光饱和点较高,这表明,HR可利用的光合有效辐射范围较广泛,但光饱和光合速率显著低于其他3个品种(P<0.05)。在4个树莓品种中,SJH的光饱和点、光饱和光合速率与表观量子效率最高,而光补偿点较低,这表明,SJH可利用的PA,R范围较广泛,是4个品种中较为喜光且光效率较高的品种。

表1 4个树莓品种的光合响应参数
2.2 4个树莓品种叶片CO2响应曲线特征
4个树莓品种的CO2响应曲线如图2所示。当CO2摩尔分数为0~300μmol·mol-1时,4个树莓品种的净光合速率随CO2摩尔分数的增加而呈线性增加;当CO2摩尔分数为300~1 500μmol·mol-1时,CO2响应曲线大致呈抛物线型。与表1的光饱和光合速率相比较,4个树莓品种叶片同化CO2速率在高CO2条件下不同程度上升,在CO2摩尔分数增加至1 500μmol·mol-1左右时,净光合速率接近最高水平。从气孔导度随CO2摩尔分数的变化趋势来看,4个树莓品种变化趋势有明显不同,当CO2摩尔分数为0~300μmol·mol-1时,随着CO2摩尔分数的增加,气孔导度均呈下降趋势,此后,SJH的气孔导度变化不明显;而QF、HR与QH 3个品种的气孔导度有明显的增长。4个树莓品种的胞间CO2摩尔分数随CO2摩尔分数变化曲线表明,胞间CO2摩尔分数随叶室中CO2摩尔分数的增加而明显上升。蒸腾速率随CO2摩尔分数变化曲线的变化趋势与气孔导度随CO2摩尔分数变化曲线十分相似。4个树莓品种相比较,其CO2补偿点、CO2饱和点、最大再生速率和羧化效率多存在显著性差异。4个树莓品种的CO2补偿点为89.95~104.43μmol·mol-1,其CO2饱和点为1 136.67~2 860.00μmol·mol-1,最大再生速率为25.75~45.41μmol·m-2·s-1,羧化效率为0.025 8~0.039 3(表2)。

图2 4个树莓品种的CO2响应曲线

表2 4个树莓品种的CO2响应参数
2.3 4个树莓品种的光合日变化
环境因子日变化如图3所示。结合表1的结果可知,08:00—16:00,环境中的PA,R在4个树莓品种光饱和点之上。气温在14:00时达到最高。CO2摩尔分数在06:00—08:00与17:00—19:00保持较高水平,09:00—17:00在小范围内波动。相对湿度在06:00与17:00—19:00保持较高水平。

图3 环境因子日变化
由图4可看出,06:00—19:00,树莓的净光合速率变化曲线大致呈双峰型,午休现象明显。4个树莓品种净光合速率的第一个峰值出现在08:00,第二个峰值大约出现在16:00。相同时刻净光合速率相比较,4个树莓品种由大到小的顺序为SJH、QF、QH、HR,这与图1的结果大致吻合。对空气湿度与净光合速率进行了相关性分析,结果表明,4个树莓品种的净光合速率与空气湿度呈显著正相关关系(P<0.05)。

图4 4个树莓品种光合作用日变化
气孔导度大致为双峰曲线(图4),第一个峰值出现在06:00—08:00,第二个峰值出现在17:00。胞间CO2摩尔分数变化趋势与大气环境中的CO2摩尔分数变化趋势相似(图3),均呈先降后升的趋势。不同品种蒸腾速率的峰值出现的时间有一定差异,第一个峰值大约出现在07:00或10:00,第二个峰值出现在14:00或16:00。使用公式(Ls=1-Ci/Ca)对气孔限制值进行计算(Ci与Ca分别为胞间CO2摩尔分数与大气CO2摩尔分数),结果表明,在08:00—16:00,4个品种的气孔限制值均维持在全天的高位水平上,这表明树莓光合的午休现象与气孔限制值的增加密切相关。
3 结论与讨论
我国树莓野生资源丰富,分布广泛,从黑龙江、新疆到长江以北,以及长江以南高海拔地区均有栽培,但资源开发利用少,目前的栽培品种多由国外引进。由于我国地域辽阔,不同地理生态环境差异较大,引入品种存在不同的生态适宜性问题。蔷薇科杏的光饱和点与补偿点分别为970~1450、40~50 μmol·m-2·s-1[7];梨的光饱和点和补偿点分别为1 100、50μmol·m-2·s-1[8];矮樱桃的光饱和点和补偿点分别为600、40μmol·m-2·s-1左右[9]。从本研究的结果来看,4个树莓品种的光饱和点为742.50~887.50μmol·m-2·s-1,其光补偿点为20.58~72.09μmol·m-2·s-1。因此,在蔷薇科植物中其光饱和点低于杏与梨而高于矮樱桃,而光补偿点与矮樱桃相接近。孙骏威等[10]研究了高粱泡(R.lambertianus)、寒莓(R.buergeri)、山莓(R.corchorifolius)、茅莓(R.parvifolius)和蓬蘽(R.hirsutus)5种悬钩子属植物叶的光合特征,其中山莓光饱和点和光补偿点光强最低,分别为495、9.3μmol·m-2·s-1;寒莓的光饱和点和光补偿点最高,分别为1 285、15.2 μmol·m-2·s-1。可见,在悬钩子属植物中,本研究中的4个品种的光饱和点介于山莓和寒莓之间,其光补偿点则明显高于以上5种悬钩子属植物。山西栽培的美国红树莓(R.strigosus Mick)具阴生植物特征[8],该品种在光照强度超过饱和点时净光合速率迅速下降;与之相比较,本研究中的4个树莓品种在光合有效辐射超过其饱和点时光抑制现象并不明显,这表明其对不同的光照条件均有较好的适应性。有学者调查了加拿大温带落叶阔叶林中红树莓(R.idaeus L.)的自然分布情况,结果表明,野生红树莓多分布于光照强度较弱的林下区域[11]。本研究中的4个树莓品种在7月中旬07:00接近光饱和,此时,光合有效辐射仅相当于最大光合有效辐射的40%左右,因此,也表现出明显的阴生植物特征,能否进行密植或林下栽培还有待生产性试验的进一步证实。
树莓光合速率、生长与温度间的关系已经有一些报道。‘秋福’最适温度为20~25℃,超过该温度范围其生长速率下降,光照强度过高则可诱导其早花,进行设施栽培更有利于其环境因子的调节[12];‘海瑞特兹’在根际土壤温度为25℃时叶片的净光合速率最高,不同月份间净合光合速率变化不明显[13]。在露地栽培条件下,7月中旬中午最高温度可达40℃左右,远偏离最佳生长温度,叶片蒸腾速度上升迅速,诱导气孔关闭,引起明显的光合午休现象。CO2是植物光合的原料,在密封温室中,施用CO2气肥有利于植物叶片对CO2的同化固定,与对照相比,温室中1 500μmol·mol-1CO2处理可使黄瓜的产量增加19.7%,增产增收效果十分显著[14]。CO2摩尔分数与树莓净光合速率间关系的研究尚不多见。从4个树莓品种叶片CO2响应曲线特征来看,‘海瑞特兹’与‘秋红’这两个品种的最大再生速率分别达到43.63、45.41μmol·m-2·s-1,约为其光饱和光合速率的4倍,这表明这两个品种应当非常适宜于密闭温室中的高CO2栽培,而‘四季红’在该环境中栽培的长势可能会远不及前两者。当叶室中CO2摩尔分数超过大气(约390μmol·mol-1)中的摩尔分数以后,‘海瑞特兹’、‘秋福’与‘秋红’的气孔导度随叶室中CO2摩尔分数增加而迅速上升,但‘四季红’的气孔对CO2摩尔分数的变化没有明显的响应,因此,CO2摩尔分数对气孔的调节作用存在明显的品种特异性;4个品种光合速率与空气湿度呈明显正相关。综上所述,露地栽培时四季红净光合速率最高,在设施栽培与高CO2摩尔分数条件下,‘秋红’和‘海瑞特兹’碳同化能力最优,适宜的温度(25℃左右)和较大的空气湿度有利于树莓光合效率的提高。
[1]Dossett M,Kempler C,Daubeny H.BC 90-19-34 and BC 93-16-43 Red Raspberries[J].Hortscience,2013,48(5):664-667.
[2]Comeau C,Prive JP,Moreau G.Effects of reflective groundcovers on ground beetles(Coleoptera:Carabidae)in red raspberry(Rubus idaeus)cropping systems[J].Journal of Applied Entomology,2013,137(4):264-274.
[3]Carvalho E,Fraser P D,Martens S.Carotenoids and tocopherols in yellow and red raspberries[J].Food Chemistry,2013,139(1/4):744-752.
[4]Neocleous D,Vasilakakis M.Effects of NaCl stress on red raspberry(Rubusidaeus L.‘Autumn Bliss’)[J].Scientia Horticulturae,2007,112(3):282-289.
[5]刘剑锋,程云清,刘强,等.平榛与平欧杂交榛叶片光合特征[J].浙江大学学报:农业与生命科学版,2012,38(4):413-420.
[6]程云清,赵桂兰,刘剑锋,等.乙烯抑制剂AVG和促进剂ACC处理对大豆幼苗叶片光合特征的影响[J].浙江大学学报:农业与生命科学版,2010,36(4):430-438.
[7]王金政,张安宁,单守明.3个设施或露地栽培常用杏品种光合特性的研究[J].园艺学报,2005,32(6):980-984.
[8]李六林,杨佩芳,田彩芳,等.树莓光合特性的研究[J].园艺学报,2003,30(3):314-316.
[9]黄卫东,吴兰坤,战吉成.中国矮樱桃叶片生长和光合作用对弱光环境的适应性调节[J].中国农业科学,2004,37(12):1981-1985.
[10]孙骏威,李素芳,金松恒.5种悬钩子属植物的光合荧光特性[J].浙江林学院学报,2010,27(6):950-955.
[11]Ricard J P,Messier C.Abundance,growth and allometry of red raspberry(Rubus idaeus L.)along a natural light gradient in a northern hardwood forest[J].Forest Ecology and Management,1996,81(1/3):153-160.
[12]Carew J G,Mahmood K,Darby J,et al.The effect of temperature,photosynthetic photon flux density,and photoperiod on the vegetative growth and flowering of‘Autumn Bliss’raspberry[J].Journal of the American Society for Horticultural Science,2003,128(3):291-296.
[13]Percival D C,Proctor JT A,Sullivan JA.Supplementary irrigation and mulch benefit the establishment of‘Heritage’primocane-fruiting raspberry[J].Journal of the American Society for Horticultural Science,1998,123(4):518-523.
[14]康康,呼世斌.净化后废气作为CO2气肥的施用效果研究[J].西北农林科技大学学报:自然科学版,2012,40(1):101-114.
