MAPK信号通路调控长波紫外线诱导的皮肤成纤维细胞组织蛋白酶K表达
2014-12-18许庆芳侯巍郑跃刘晨龚子鉴陆春赖维
许庆芳 侯巍 郑跃 刘晨 龚子鉴 陆春 赖维
MAPK信号通路调控长波紫外线诱导的皮肤成纤维细胞组织蛋白酶K表达
许庆芳 侯巍 郑跃 刘晨 龚子鉴 陆春 赖维
目的研究MAPK信号通路调控长波紫外线(UVA)诱导皮肤成纤维细胞组织蛋白酶K(CatK)的表达。方法原代培养的皮肤成纤维细胞取自儿童包皮。Western印迹检测10 J/cm2UVA照射前及照射后0.75、1.5、3和6 h皮肤成纤维细胞中磷酸化JNK(p-JNK)、JNK、磷酸化P38(p-P38)、P38蛋白的表达。800 nmol/L SP600125(SP)、10 μmol/L SB203580(SB)分别孵育皮肤成纤维细胞,实验分为无UVA照射的对照组、SP组、SB组及10 J/cm2UVA照射的对照(UVA)组、UVA-SP组、UVA-SB组。先以Western印迹检测各组UVA照射后1.5 h磷酸化c-Jun(p-c-Jun)和磷酸化MAPKAPK2(p-MAPKAPK2)表达,再以RT-PCR和Western印迹检测各组照射后48 h CatK mRNA、蛋白的表达。结果皮肤成纤维细胞在UVA照射后0.75、1.5 h,p-JNK表达的灰度值分别为4.77±0.19和4.68±0.09,p-P38分别为2.44±0.13、2.30±0.04,均较照射前(p-JNK为3.2±0.27,p-P38为1.61±0.08)显著升高(均P<0.05);而在照射后3、6 h的表达与照射前相比差异无统计学意义(P>0.05)。p-c-Jun在UVA-SP组表达(2.55±0.48)、p-MAPKAPK2在UVA-SB组表达(1.16±0.12)均显著低于UVA组(分别为4.85±0.96和2.46±0.09),均P<0.05。UVA-SP组、UVA-SB组CatK mRNA、蛋白的表达分别降为UVA组的38.9%、28.7%和55.7%、49.6%(P<0.05),UVA-SP组CatK mRNA、蛋白的表达也均显著低于UVA-SB组(P<0.05)。结论JNK和P38信号通路在调控UVA上调的皮肤成纤维细胞CatK表达中起重要作用。
成纤维细胞;组织蛋白酶类;MAP激酶信号系统;紫外线;皮肤衰老
皮肤光老化最具特征性的病理改变为变性弹性纤维在真皮中上层的堆积,即光线性弹性组织变性。其形成机制包括弹性纤维变性机制和堆积机制[1]。组织蛋白酶K(cathepsin K,CatK)属于溶酶体内半胱氨酸蛋白酶家族的成员之一,是皮肤成纤维细胞内降解弹性蛋白的主要蛋白酶,在光线性弹性组织变性中起重要作用[2]。我们的前期研究已揭示,长波紫外线(UVA)促进皮肤成纤维细胞中CatK表达[3]。然而,调控UVA诱导皮肤成纤维细胞中CatK表达的机制目前还不清楚。UVA主要激活皮肤成纤维细胞的丝裂原激活蛋白激酶(mitogen-activated protein kinases,MAPK)通路。本实验检测UVA对皮肤成纤维细胞JNK和P38通路的影响,并以SP600125和SB203580分别抑制JNK和P38通路,再采取RTPCR和Western印迹检测JNK和P38通路的抑制对UVA诱导的CatK表达的影响,以研究MAPK信号通路是否调控UVA诱导的皮肤成纤维细胞中CatK表达。
材料与方法
一、实验材料
1.皮肤成纤维细胞的组织来源:健康人皮肤组织来自于中山大学附属第三医院泌尿外科健康儿童包皮环切术后的包皮组织,年龄5~9岁。本研究经中山大学附属第三医院伦理委员会批准。
2.试剂和仪器:DMEM(Dulbecco's modified Eagle's media)高糖培养基、胰酶、胎牛血清、磷酸盐缓冲液(PBS)、青链霉素为美国Gibco公司产品。SP600125(SP)、SB203580(SB)为德国默克公司产品。一抗兔抗人CatK-IgG抗体为美国Abcam公司产品;一抗兔抗人磷酸化JNK(p-JNK)、兔抗人JNK、兔抗人磷酸化c-Jun(p-c-Jun)、兔抗人磷酸化P38(p-P38)、兔抗人 P38、兔抗人磷酸化MAPKAPK2(p-MAPKAPK2)、内参兔抗人GAPDH多克隆IgG抗体及二抗HRP-羊抗兔IgG均为美国Cell signaling technology公司产品。BCA蛋白定量试剂盒为美国Pierce公司产品。ECL显色试剂盒为美国Millipore公司产品,预染Marker为加拿大Mbi fermentas公司产品。总RNA提取试剂Trizol为美国Invitrogen公司产品。Primescript RT master mix逆转录试剂盒和Sybrpremix ex taqTm试剂盒为日本Takara公司产品。UVA紫外线辐射仪(Sigma ss-03A)和UVA辐照计(Sigma ss-03)均由上海希格玛高科技有限公司生产,灯管为Philips UVA TL10RS,波长320~400 nm;酶联免疫检测仪(美国Biotek公司);CO2细胞培养箱(美国Thermo scientific公司);Abi prism 7500型实时荧光定量PCR仪。
二、方法
1.原代皮肤成纤维细胞培养:取儿童包皮,参照文献[3]分离培养皮肤成纤维细胞,第3代细胞冻存。细胞复苏后10代以内的细胞行后续实验。
2.UVA照射后不同时间皮肤成纤维细胞p-JNK、JNK、p-P38、P38蛋白表达的变化:将3~10代的成纤维细胞按106接种于6 cm细胞培养皿,培养24 h至80%~90%汇合时取出,吸弃培养液,用PBS洗2次,每皿加入等量的薄层PBS。按照文献[3]的方法,把培养皿置于UVA紫外线辐射仪下,距离紫外光源约15 cm,平均照射功率为13 mW/cm2,照射769 s,使照射1次的UVA剂量达10 J/cm2。照射后立即吸弃PBS,加入新鲜培养液,置于95%湿度、5%CO237 ℃培养箱中培养。在照射后 0、0.75、1.5、3、6 h提取蛋白,Western印迹检测各时间点p-JNK、JNK、p-P38、P38蛋白表达的变化。实验重复3次。
3.SP、SB对UVA照射的皮肤成纤维细胞p-MAPKAPK2、p-c-Jun 表达的影响:10 μmol/L SB 工作浓度的选择参照生产厂家推荐及文献[4]。参照文献[4],采用CCK8和Western印迹筛选出800 nmol/L SP可抑制UVA激活的JNK通路。实验设无UVA照射的对照组、800 nmol/L SP 的 SP 组、10 μmol/L SB的SB组及10 J/cm2UVA照射的UVA组、800 nmol/L SP的UVA-SP组、10 μmol/L SB的UVA-SB组。 将细胞从培养箱中取出,向培养皿中分别加入SP、SB(终浓度分别为 800 nmol/L、10 μmol/L)孵育 1 h,吸弃培养液,用PBS洗2次,每皿加入等量的薄层PBS,予 10 J/cm2UVA照射1次,再在含SP、SB的培养基中孵育1.5 h,提取细胞蛋白,最后以Western印迹对比检测p-MAPKAPK2、p-c-Jun在各组的表达变化。实验重复3次。
4.SP、SB对UVA照射的皮肤成纤维细胞CatK表达的影响:按以上方法分组,照射前SP和SB分别孵育细胞及UVA照射后再在含SP、SB的培养基中孵育48 h,提取RNA和蛋白,最后以RT-PCR和Western印迹对比检测各组CatK的表达变化。实验重复3次。
5.RT-PCR检测皮肤成纤维细胞中CatK mRNA的表达变化:按照Trizol试剂盒的方法提取总RNA。按逆转录试剂盒说明书配制逆转录反应体系,总体积20 μl,37℃下反转录15 min,85℃下5 s使反转录酶失活,即获得cDNA,置-20℃冰箱中保存。采用染料法(SYBR Green I)进行相对定量分析。PCR反应体系共20 μl,包括SYBR® Premix Ex TaqⅡ 10 μl,ROXDyeⅡ 0.4 μl,双蒸水 6 μl,上下游引物各 0.8 μl和 cDNA 2 μl。 两步法实时定量PCR:95℃5 s,1个循环;95℃5 s,60℃30 s,40个循环。CatK上游引物序列5'-GTCTGAGAATGATGG CTGTGGA-3',下游5'-CATTTAGCTGCCTTGCCTGT TG-3',扩增片段长度150 bp。GAPDH上游引物序列5'-GCACCGTCAAGGCTGAGAAC-3',下游5'-TGGTGAAGACGCCAGTGGA-3',扩增片段长度138 bp。在实时荧光定量PCR仪上读取扩增曲线、熔解曲线和Ct值。重复3次实验,结果取均值,用2-ΔΔCt法计算目的基因和内参基因的表达相对值。
6.Western印迹检测皮肤成纤维细胞中CatK、p-JNK、JNK、p-P38、P38、p-MAPKAPK2、p-c-Jun 蛋白的表达变化:提取各组细胞总蛋白-80℃保存。BCA法蛋白定量。总蛋白经12%SDS-PAGE分离后,电转移到PVDF膜上。一抗兔抗人CatK-IgG(滴度 1∶500)、兔抗人 p-JNK-IgG(滴度 1∶1 000)、兔抗人JNK-IgG(滴度1∶1 000)、兔抗人p-P38(滴度1∶1 000)、兔抗人 P38(滴度 1∶1 000)、兔抗人 p-MAPKAPK2(滴度1∶1 000)、兔抗人p-c-Jun(滴度1∶1 000),内参兔抗人GAPDH-IgG(滴度1∶4 000),4℃孵育过夜,TBST液洗膜,加入HRP-羊抗兔IgG(滴度1∶1 000)37℃ 孵育1 h,TBST液洗膜。ECL显色。所有实验重复3次。
结 果
一、皮肤成纤维细胞中 p-JNK、JNK、p-P38、P38蛋白的表达随UVA照射后不同时间的变化
见图 1、表 1。UVA 照射后 0、0.75、1.5、3、6 h,各组p-JNK、p-P38灰度值经单因素方差分析,差异均有统计学意义(P<0.05)。LSD-t检验发现,p-JNK、p-P38在照射后0.75、1.5 h的表达均明显高于0 h(P<0.05),在3、6 h的表达与0 h相比差异均无统计学意义(P>0.05)。JNK、P38蛋白表达在各时间点变化无明显差异。
二、SP、SB对UVA激活的皮肤成纤维细胞JNK和P38信号通路的影响
见图 2、表 2。 p-c-Jun、p-MAPKAPK2在各组的灰度值经单因素方差分析,差异均有统计学意义(P<0.01)。LSD-t检验发现,UVA组和UVA-SB组p-c-Jun表达差异无统计学意义(P>0.05),但UVASP组明显低于UVA组(P<0.05);UVA组与UVASP组p-MAPKAPK2的表达差异无统计学意义(P>0.05),但UVA-SB组明显低于UVA组(P<0.01)。

图1 皮肤成纤维细胞中p-JNK、JNK、p-P38、P38表达随UVA照射后不同时间变化的电泳图 1~5:分别为UVA照射后0、0.75、1.5、3、6 h组。p-JNK、p-P38在UVA 照射后0.75、1.5 h均较0 h显著升高,后恢复至0 h水平,而JNK、P38在各时间点表达无显著差异
表1 UVA照射皮肤成纤维细胞后不同时间p-JNK、JNK、p-P38、P38蛋白表达变化(灰度值,±s)
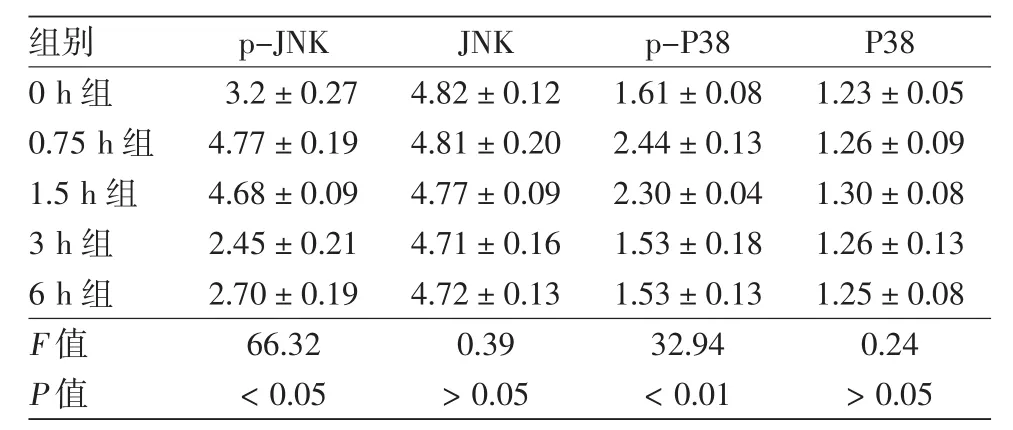
表1 UVA照射皮肤成纤维细胞后不同时间p-JNK、JNK、p-P38、P38蛋白表达变化(灰度值,±s)
注:n=3
p-JNK JNK p-P38 P38 3.2±0.27 4.82±0.12 1.61±0.08 1.23±0.05 4.77±0.19 4.81±0.20 2.44±0.13 1.26±0.09 1.5 h组 4.68±0.09 4.77±0.09 2.30±0.04 1.30±0.08 3 h组 2.45±0.21 4.71±0.16 1.53±0.18 1.26±0.13 6 h组 2.70±0.19 4.72±0.13 1.53±0.13 1.25±0.08 F值 66.32 0.39 32.94 0.24 P值 <0.05 >0.05 <0.01 >0.05
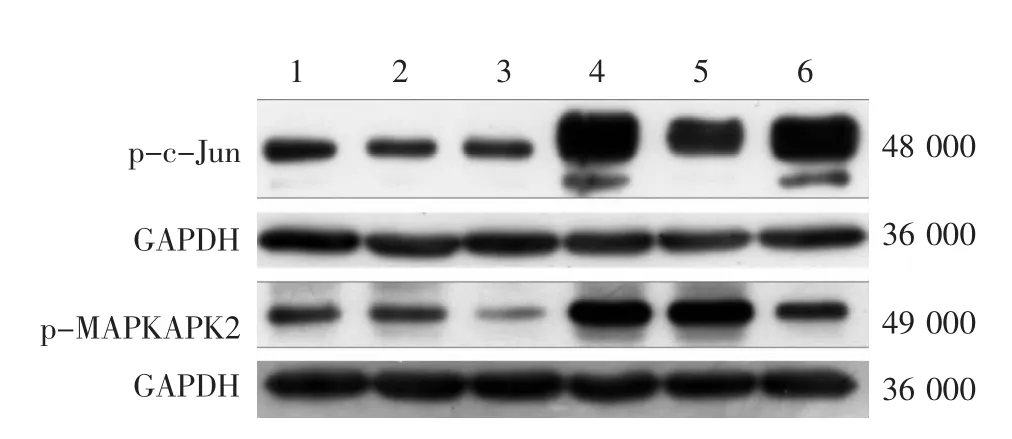
图2 SP600125、SB203580抑制UVA激活的皮肤成纤维细胞JNK和P38信号通路的电泳图 1:对照组;2:SP组;3:SB组;4:UVA组;5:UVA-SP组;6:UVA-SB组。p-c-Jun在UVA-SP组、p-MAPKAPK2在UVA-SB组的表达均显著低于UVA组
表2 SP、SB对UVA激活的皮肤成纤维细胞JNK和P38信号通路及CatK表达的影响(±s)

表2 SP、SB对UVA激活的皮肤成纤维细胞JNK和P38信号通路及CatK表达的影响(±s)
注:n=3
组别 p-c-Jun(灰度值)CatK蛋白(灰度值)对照组 1.63±0.17 1.03±0.13 0.172±0.012 1.58±0.24 SP组 1.46±0.38 1.02±0.18 0.170±0.013 1.57±0.26 SB组 1.48±0.33 0.36±0.08 0.110±0.010 1.02±0.32 UVA组 4.85±0.96 2.46±0.09 0.517±0.030 2.68±0.36 UVA-SP组 2.55±0.48 2.45±0.09 0.201±0.026 0.77±0.10 UVA-SB组 4.61±0.71 1.16±0.12 0.288±0.037 1.33±0.44 F值 23.29 157.43 146.66 14.14 P值 <0.01 <0.01 <0.01 <0.01 p-MAPKAPK2(灰度值)CatK mRNA(2-ΔΔCt)

图3 SP、SB对UVA诱导的皮肤成纤维细胞CatK蛋白表达影响的电泳图 1:对照组;2:SP组;3:SB组;4:UVA组;5:UVASP组;6:UVA-SB组。UVA组CatK蛋白表达显著高于对照组,UVA-SP组、UVA-SB组的蛋白表达均显著低于UVA组,在UVASP组的蛋白表达显著低于UVA-SB组
三、SP、SB对 UVA诱导的皮肤成纤维细胞CatK表达的影响
见表2、图3。Western印迹可检测到CatK酶原和活性CatK两个条带。CatK mRNA表达及CatK蛋白灰度值6组比较,差异均有统计学意义。LSD-t检验发现,UVA组CatK mRNA、蛋白表达均显著高于对照组(P<0.05);UVA-SP组、UVA-SB组CatK mRNA、蛋白表达均显著低于UVA组(P<0.05),且UVA-SP组CatK表达均显著低于UVA-SB组(P<0.05)。
讨 论
组织蛋白酶K属于溶酶体内半胱氨酸蛋白酶家族的成员,是目前最强的降解弹性蛋白和胶原蛋白的酶[5]。最近研究发现,CatK是皮肤成纤维细胞内降解弹性蛋白的主要蛋白酶。光老化皮肤及皮肤成纤维细胞中CatK表达和活性降低,使得胞内弹性蛋白降解减少,从而在光老化病变弹性纤维堆积中起作用[6]。然而,我们的前期研究揭示,急性UVA照射促进皮肤成纤维细胞中CatK表达[3]。目前调控UVA诱导的光老化皮肤成纤维细胞中CatK表达的机制还不清楚。 UVA可激活皮肤成纤维细胞的丝裂原激活蛋白激酶(mitogen-activated protein kinases,MAPK) 通路、NF-κB通路及PI3K-AKT通路等,但主要为MAPK通路[7]。有研究报道,MAPK通路调控CatK在其他细胞的表达[8],所以我们研究该信号通路是否也调控UVA上调的皮肤成纤维细胞中CatK表达。
MAPK通路主要包含JNK、P38和ERK三条通路,而UVA只激活前两者[4]。本研究发现,皮肤成纤维细胞中的p-JNK、p-P38在UVA照射后0.75、1.5 h均较0 h显著升高,后恢复至0 h水平,说明本实验所用10 J/cm2UVA可激活皮肤成纤维细胞JNK和P38通路。JNK和P38通路的活化分别磷酸化下游的 c-Jun 和 MAPKAPK2[4]。 SP600125 和 SB203580分别是JNK和P38通路的高选择性抑制剂,它们对这两条通路的抑制分别表现为下调p-c-Jun、p-MAPKAPK2 表达[9-10],所以我们以 Western 印迹检测 SP、SB对 UVA照射的皮肤成纤维细胞 p-MAPKAPK2、p-c-Jun表达的影响,以研究800 nmol/L SP和10 μmol/L SB能否分别特异性地抑制UVA激活的JNK和P38通路。10 μmol/L SB工作浓度的选择参照文献[4,11]。 因 UVA 照射 SP 孵育的 HaCaT细胞可导致细胞凋亡,所以我们采用CCK8和Western印迹实验筛选出 800 nmol/L SP既可保持成纤维细胞活性在90%以上,也可抑制UVA激活的JNK通路[12]。Western印迹结果显示,p-c-Jun在UVA-SP组表达显著低于UVA组、p-MAPKAPK2在UVA-SB组的表达显著低于UVA组,表明本实验采用800 nmol/L SP和10 μmol/L SB可分别抑制UVA活化的JNK和P38通路。
我们采用RT-PCR、Western印迹观察以SP和SB分别抑制JNK和P38通路活化对UVA上调的皮肤成纤维细胞CatK表达的影响,结果发现,UVA组CatK mRNA、蛋白的表达显著高于对照组,表明UVA促进皮肤成纤维细胞CatK表达。UVA-SP组、UVA-SB组CatK mRNA、蛋白表达均显著低于UVA组,且UVA-SP组CatK mRNA、蛋白表达均显著低于UVA-SB组,提示JNK和P38通路的抑制均可从基因、蛋白水平降低UVA诱导的CatK表达,前者对UVA诱导的CatK表达的抑制程度高于后者。由此我们推测,UVA很可能通过激活JNK和P38通路、且以前者为主刺激皮肤成纤维细胞CatK的表达。这与调控CatK在其他细胞表达的信号通路相似。在巨噬细胞中,其表面沉积的胆固醇通过活化P38MAPK刺激CatK的表达[13];在血管内皮细胞中,肿瘤坏死因子α通过激活JNK通路促进CatK的表达[14];在关节软骨细胞中,Ⅱ型胶原的N-端肽通过激活蛋白激酶C和P38MAPK通路上调CatK的表达[15]。
已有研究揭示,UVA通过激活MAPK通路上调AP-1的转录活性,活化的AP-1与基质金属蛋白酶(matrix metalloproteinases,MMP)启动子结合,使得启动子被反式激活而导致MMP的表达上调[16]。由此可见,UVA诱导皮肤成纤维细胞MMP和CatK表达的调控途径有类似的地方,UVA很可能也通过激活MAPK/AP-1通路上调CatK表达。然而,CatK表达在急性UVA照射的皮肤成纤维细胞中增加,在光老化皮肤及皮肤成纤维细胞中却下降。我们推测这可能由于光老化的皮肤成纤维细胞CatK的启动子结构或功能因紫外线损伤发生改变,使之不能与AP-1很好结合,抑或不能启动CatK基因的转录;也可能因光老化皮肤成纤维细胞线粒体突变、DNA损伤及降低的细胞新陈代谢等致CatK基因转录、翻译下降。至于UVA照射早期皮肤成纤维细胞表达上调的CatK在光老化过程中如何降低及其确切机制,尚需进一步研究。
[1]许庆芳,赖维,郑跃.光线性弹性组织变性形成机制的研究进展[J].国际皮肤性病学杂志,2012,38(1):45-47.
[2]Codriansky KA,Quintanilla-Dieck MJ,Gan S,et al.Intracellular degradation of elastin by cathepsin K in skin fibroblasts--a possible role in photoaging[J].Photochem Photobiol,2009,85(6):1356-1363.
[3]许庆芳,侯巍,刘晨,等.UVA照射对皮肤成纤维细胞组织蛋白酶K表达的影响[J].中华皮肤科杂志,2013,46(9):652.
[4]Silvers AL,Bachelor MA,Bowden GT.The role of JNK and p38 MAPK activities in UVA-induced signaling pathways leading to AP-1 activation and c-Fos expression[J].Neoplasia,2003,5(4):319-329.
[5]Rünger TM,Quintanilla-Dieck MJ,Bhawan J.Role of cathepsin K in the turnover of the dermal extracellar matrix during scar formation[J].J Invest Dermatol,2007,127(2):293-297.
[6]Zheng Y,Lai W,Wan M,et al.Expression of cathepsins in human skin photoaging [J].Skin Pharmacol Physiol,2011,24(1):10-21.
[7]Zheng J,Lai W,Zhu G,et al.10-Hydroxy-2-decenoic acid prevents ultravioletA-induced damage and matrix metalloproteinases expression in human dermal fibroblasts[J].J Eur Acad Dermatol Venereol,2013,27(10):1269-1277.
[8]Matsumoto M,Kogawa M,Wada S,et al.Essential role of p38 mitogen-activated protein kinase in cathepsin K gene expression during osteoclastogenesis through association of NFATc1 and PU.1[J].J Biol Chem,2004,279(44):45969-45979.
[9]Bennett BL,Sasaki DT,Murray BW,et al.SP600125,an anthrapyrazolone inhibitor of jun N-terminal kinase[J].Proc Natl Acad Sci USA,2001,98(24):13681-13686.
[10]Kumar S,Jiang MS,Adams JL,et al.Pyridinylimidazole compound SB 203580 inhibits the activity but not the activation of p38 mitogen-activated protein kinase [J].Biochem Biophys Res Commun,1999,263(3):825-831.
[11]Schieke S,Stege H,Kürten V,et al.Infrared-A radiation-induced matrix metalloproteinase 1 expression ismediated through extracellular signal-regulated kinase 1/2 activation in human dermal fibroblasts[J].J Invest Dermatol,2002,119(6):1323-1329.
[12]许庆芳,侯巍,郑跃,等.SP600125抑制UVA激活皮肤成纤维细胞JNK通路的最佳浓度[J].中山大学学报(医学科学版),2014,35(2):194-199.
[13]Sun Y,Ishibashi M,Seimon T,et al.Free cholesterol accumulation in macrophage membranes activates Toll-like receptors and p38 mitogen-activated protein kinase and induces cathepsin K[J].Circ Res,2009,104(4):455-465.
[14]Keegan PM,Wilder CL,Platt MO.Tumor necrosis factor alpha stimulates cathepsin K and V activity via juxtacrine monocyteendothelial cell signaling and JNK activation [J].Mol Cell Biochem,2012,367(1-2):65-72.
[15]Ruettger A,Schueler S,Mollenhauer JA,et al.Cathepsins B,K and L are regulated by a defined collagen typeⅡpeptide via activation of classical protein kinase C and p38 MAP kinase in articular chondrocytes[J].J Biol Chem,2008,283(2):1043-1051.
[16]Hwang YP,Oh KN,Yun HJ,et al.The flavonoids apigenin and luteolin suppress ultraviolet A-induced matrix metalloproteinase-1 expression via MAPKs and AP-1-dependent signaling in HaCaT cells[J].J Dermatol Sci,2011,61(1):23-31.
MAPK pathway regulates ultraviolet A-induced cathepsin K expression in human dermal fibroblasts
Xu Qingfang,Hou Wei,Zheng Yue,Liu Chen,Gong Zijian,Lu Chun,Lai Wei.Department of Dermatology,Third Affiliated Hospital,Sun Yat-sen University,Guangzhou 510630,China
Lai Wei:Email:drlaiwei@163.com
Objective To investigate whether ultraviolet A(UVA)-induced CatK expression is regulated by the mitogen-activated protein kinases(MAPK)signaling pathway in human dermal fibroblastsin vitro.Methods Human dermal fibroblasts were obtained from circumcised foreskin of children,and subjected to primary culture.After several passages of subculture,some fibroblasts were irradiated with UVA at a dose of 10 J/cm2.Western blot was performed to measure the expressions of total and phosphorylated JNK(t-and p-JNK)and P38(t-and p-P38)at 0.75,1.5,3 and 6 hours after the irradiation.Some fibroblasts were divided into six groups:control group receiving no treatment,SP group treated with SP600125 of 800 nmol/L,SB group treated with SB203580 of 10 μmol/L,UVA group irradiated with UVA at a dose of 10 J/cm2,UVA-SP group treated with SP600125 for 1 hour before and for 1.5 or 48 hours after UVA irradiation at 10 J/cm2,UVA-SB group treated with SB203580 for 1 hour before and for 1.5 or 48 hours after UVA radiation at 10 J/cm2.Subsequently,Western blot was performed to determine the expressions of p-c-Jun and p-MAPKAPK2 in these groups at 1.5 hours after the UVA irradiation,and reverse transcription(RT)-PCR and Western blot to detect the mRNA and protein expressions of CatK at 48 hours after the UVA irradiation,respectively.Statistical analysis was carried out byttest,one way analysis of variance and least significant difference(LSD)-ttest.Results The expression levels(gray values)of p-JNK and p-P38 were significantly increased at 0.75 hour(4.77±0.19 and 2.44±0.13 respectively,bothP<0.05)and 1.5 hours(4.68±0.09 and 2.30±0.04 respectively,bothP<0.05),but showed no significant changes at 3 hours(bothP>0.05)and 6 hours(bothP>0.05)after the UVA irradiation compared with those before the irradiation(3.2±0.27 and 1.61±0.08 respectively).A significant decrease was observed in the expression of p-c-Jun in the UVA-SP group and p-MAPKAPK2 in the UVA-SB group compared with the UVA group(p-c-Jun,2.55±0.48 vs.4.85±0.96;p-MAPKAPK2,1.16 ± 0.12 vs.2.46 ± 0.09,bothP< 0.05).The CatK mRNA and protein expressions were attenuated by 61.1%and 44.3%respectively in the UVA-SP group(bothP< 0.05),and by 71.3%and 50.4%respectively in the UVA-SB group(bothP<0.05)in comparison with the UVA group.The UVA-SP group also showed a significant reduction in CatK mRNA and protein expressions as compared with the UVA-SB group(bothP< 0.05).Conclusion Both JNK and P38 signaling pathways,especially the JNK pathway,may contribute to the upregulation of CatK expression in dermal fibroblasts induced by UVA irradiation.
Fibroblasts;Cathepsins;MAP kinase signaling system;Ultraviolet rays;Skin aging
10.3760/cma.j.issn.0412-4030.2014.08.003
国家自然科学基金(81171523);广东省自然科学基金(10151008901000117);广东省科技计划项目基金(2012B031800057);2010年中国医师协会-宝洁基金;2011年中国医师协会-资生堂DQ基金
510630广州,中山大学附属第三医院皮肤科
赖维,Email:drlaiwei@163.com
2013-09-30)
(本文编辑:颜艳)
