真鲷、黑鲷及其杂交子代的染色体构成与AFLP分析
2014-10-27林勉苗亮李明云俞寅寅徐万土
林勉,苗亮,李明云*,俞寅寅,徐万土
(1.宁波大学 应用海洋生物技术教育部重点实验室,浙江 宁波315211;2.象山港湾水产苗种有限公司,浙江 象山315702)
1 引言
真鲷Pagrosomusmaj or和黑鲷Spr ausmacrocephalus均是我国重要的海水养殖经济鱼类。真鲷属鲈形目、鲷科、真鲷属,具有个体大、生长快、肉质好、色泽美、抗病能力强等特点,但对盐度、温度敏感;黑鲷属鲈形目、鲷科、黑鲷属,对温度、盐度适应范围较广,但养成周期长、抗病能力弱[1]。二者各有优缺点,通过杂交育种,有可能获得具有双亲优良性状的杂交子代。目前已有鲷科鱼类杂交育种的研究报道,如江世贵等[2]进行了平鲷(♀)与真鲷(♂)的杂交;单保党等[3]对真鲷、黑鲷进行了正、反交,均获得了杂交子代;蒋宏雷等[4]、姜景腾等[5]的研究显示由真鲷(♀)×黑鲷(♂)所得的杂交子代具有生长速度快、适温盐广、营养价值高、抗病能力强等特点。虽然现有研究显示真鲷(♀)×黑鲷(♂)的杂交子代呈现出明显的杂交优势,但尚不清楚杂交子代的遗传物质构成情况。笔者采用核型和AFLP分析,研究了真鲷、黑鲷及杂交子代的遗传构成。
核型和AFLP分析作为遗传学研究的有效手段,被广泛的应用于海洋生物学的研究中,如杂交子代的染色体组成与双亲的差异分析[6],群体遗传结构的检测[7—8]、种质鉴定[9]、遗传连锁图谱的构建[10—13]、基因定位[14]等。本研究不但能了解杂交育种后代的染色体组成、与双亲染色体组型的差异和杂种染色体的归宿,而且能掌握亲本和子代3个群体的遗传结构,互为补充更能揭示杂交子代杂种优势的遗传情况,其结果可以为杂种优势研究和鲷科鱼类杂交育种实践提供理论依据和参考资料。
2 材料与方法
2.1 材料
实验用真鲷、黑鲷、杂交子代各10尾,均取自浙江象山港湾水产苗种有限公司,真鲷平均体质量29.5 g、体长12.9 cm,黑鲷平均体质量24.9 g、体长11.8 m,杂交子代平均体质量30.5 g,体长11.8 cm。
AFLP试剂盒购自北京鼎国昌盛生物技术有限责任公司;PHA、秋水仙素购自Sigma公司;甲醇、乙酸、硝酸银等均购自生工生物工程(上海)有限公司;TaqDNA聚合酶、T4 DNA连接酶等购自宝生物工程 (大连)有限公司。
2.2 染色体标本制备与核型分析
以体细胞体内培养法制备真鲷、黑鲷及杂交子代的染色体标本,具体为:按10μg/g鱼体质量剂量体腔注射PHA,16~36 h后再按3μg/g鱼体质量剂量体腔注射秋水仙素溶液,4 h后取头肾,以常规空气干燥法制备染色体玻片标本,经0.037 5 mol/L的氯化钾溶液低渗处理和固定液(甲醇:乙酸=3∶1)固定,Giemsa液染色后镜检。
对真鲷、黑鲷及杂交子代的染色体标本分别取100个染色体分散良好的中期分裂相进行染色体计数,并选出10个清晰的中期分裂相进行显微照相、放大和测量。按Levan等[15]的标准对染色体进行分类;按Corman[16]的方法统计臂数,即中部和亚中部着丝粒染色体的臂数计为2,亚端部和端部着丝粒计为1。
2.3 AFLP扩增与分析
以常规酚/氯仿/异戊醇法提取真鲷、黑鲷、杂交子代的基因组DNA。参照王志勇等[17]的方法进行AFLP扩增。通过预实验筛选出能在3种鱼种扩增到清晰条带的2对AFLP选扩引物组合(E-AAC/M-CCT和E-ACA/M-CGA)进行扩增分析,引物序列见表1。扩增产物经8%变性聚丙烯酰胺凝胶电泳分离条带,硝酸银染色后凝聚成像。参照严骏骢等[18]和王晓清等[19]的方法进行条带统计和数据分析。有关计算公式参照如下:
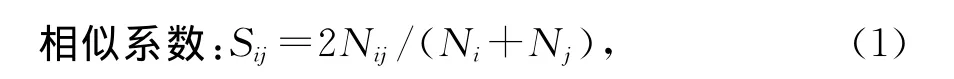
式中,Nij为个体i与j共有的片段数量,Ni、Nj分别为个体i与j各自具有的片段数量;

式中,S为相似系数。
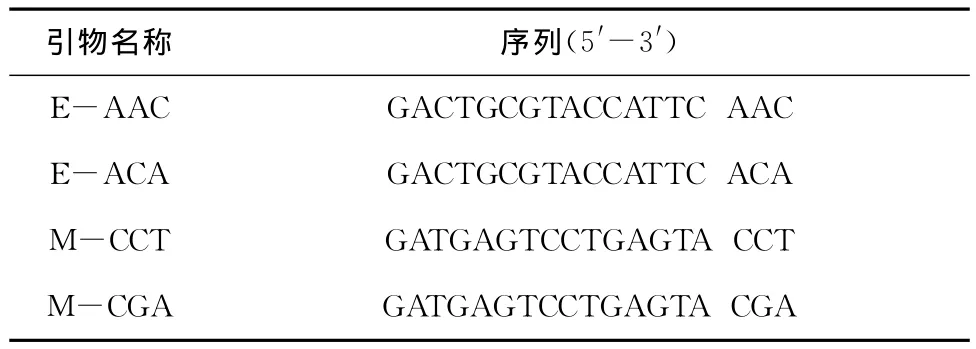
表1 AFLP选扩引物序列Tab.1 Sequence of AFLP selective amplification pri mer
3 结果
3.1 真鲷、黑鲷及其杂交子代的染色体核型
中期染色体观察计数结果显示真鲷、黑鲷和杂交子代中染色体数为48的分裂相分别占总数的68%、77%和60%(表2),因此确定三者的染色体众数均为48。
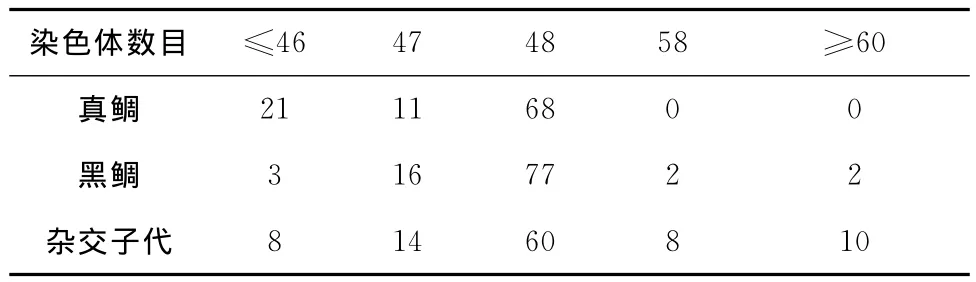
表2 不同染色体数目的分裂相个数占全部分裂相个数的百分比Tab.2 The percentage of different number of chromosomes split phase number
真鲷、黑鲷和杂交子代的染色体中期分裂相见图1。真鲷染色体核型为2n=2st+46t,NF=48(见图1a);黑鲷染色体核型为2n=4 m+4sm+2st+38t,NF=56(见图1b);杂交子代染色体核型为2n=30 m+8sm+2st+8t=48,NF=86(见图1c)。真鲷的染色体只有st和t两种类型;黑鲷的染色体有st、t、sm和m 4种类型。杂交子代虽然染色体类型与黑鲷相同,但各类型染色体数目与黑鲷不同:黑鲷染色体以t型居多,而杂交子代则是m型居多。
3.2 真鲷、黑鲷及其杂交子代的AFLP分析
两对AFLP选择性扩增引物组合(E-AAC/M-CCT和E-ACA/M-CGA)对真鲷、黑鲷及杂交子代的扩增结果显示,共检出扩增条带278个;真鲷特有条带108条,有67条出现在杂交子代中;黑鲷特有条带93条,有21条出现在杂交子代中;杂交子代中出现了15条非亲条带(表3)。

图1 中期分裂相染色体及核型Fig.1 The metaphase chromosome and karyotype
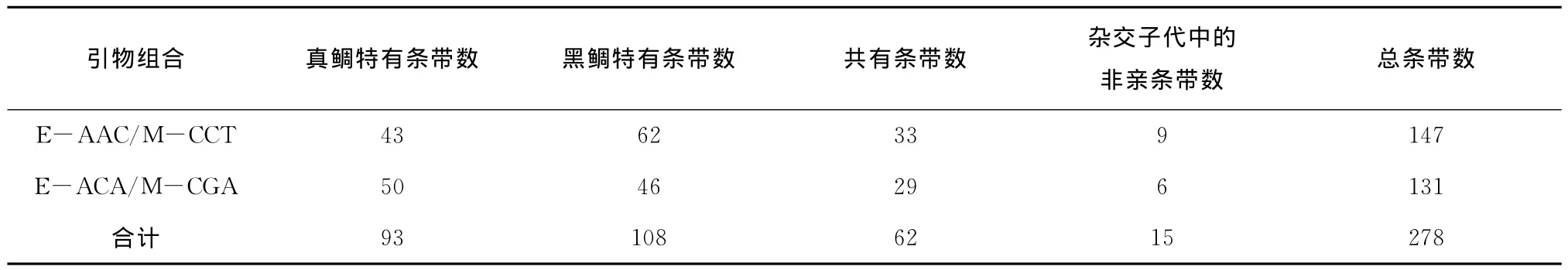
表3 AFLP扩增条带统计Tab.3 Numbers of amplified bands with 2 AFLP selective primer pairs
AFLP条带计算出杂交子代与真鲷、黑鲷的相似系数分别为0.350和0.113,杂交子代个体间的相似系数为0.709;杂交子代与真鲷、黑鲷的遗传距离分别为1.050和2.180,杂交子代个体间的遗传距离为0.344。结果显示杂交子代与母本种(真鲷)的遗传相似度更高、遗传距离更近。
4 讨论
在杂交育种中,杂交子代由于具有双亲的遗传物质而同时具有父本种和母本种的一些性状,并且杂交子代还可能因双亲的遗传物质的重组而出现一些异于双亲的性状。如黄帆[20]对高加索三叶草与白三叶杂交后代的遗传特性进行了研究,结果显示杂种F1代是双亲真实的杂交后代,其抗旱能力和生育期均介于两者中间。周丽清等[21—22]对栉孔扇贝和虾夷扇贝的正反交子代进行了核型分析,结果显示正反交子代均为偏向母本类型的异源二倍体,杂交子代显著提高了栉孔扇贝的生产性能尤其是抗逆能力。核型分析显示真鲷、黑鲷、及其杂交子代的染色体数目均为48条,但具体到染色体类型及各类型染色体数目,杂交子代与其父本种和母本种均存在较大差异:杂交子代有15对中部着丝粒染色体,4对亚中部着丝粒染色体,4对端部着丝粒染色体和1对亚端部着丝粒染色体,其父本种(黑鲷)虽然也是有这4种类型的染色体,但数目分别为2对、2对、1对和19对,而母本种(真鲷)则仅有端部(23对)和亚端部(1对)着丝粒染色体。表明真鲷和黑鲷的遗传物质在杂交过程中发生了较大的重组。
AFLP结果显示108条真鲷特有条带中有67条出现在杂交子代中,93条黑鲷特有条带中有21条出现在杂交子代中,说明子代是同时含有父、母本遗传物质的真正的杂交种。遗传相似系数和遗传距离分析表明,杂交子代与母本种(真鲷)的遗传相似度更高、遗传距离更近,说明杂交子代与母本之间具有遗传同质性,而与父本的遗传差异则较大。隋班良等[23]对黄姑鱼(♀)与大黄鱼(♂)杂交子代的AFLP分析结果也显示杂交子代在遗传上偏向母本。另外,在杂交子代中还出现了15条非亲条带,这与核型分析结果相互印证,表明父、母本遗传物质在杂交子代中发生了重组。万俊芬等[24]对栉孔扇贝和华贵栉孔扇贝杂交的ISSR检测显示子代中非亲位点的出现比例为16.7%~18.7%;何斌等[25]发现栉孔扇贝和虾夷扇贝正反杂交子代中分别有12.9% 和14.3%ISSR位点为非亲位点。非亲位点可能来源于不同长度的扩增片段之间形成的异源双链[26],或者由于配子形成过程中染色体的不等价交换产生的新序列[27]。遗传物质的重组可能会产生新的性状,蒋宏雷等[4]、姜景腾等[5,28]的研究结果均显示,与父母本相比,其杂交子代表现出明显的杂交优势,并具有部分异于双亲的新性状,如耐低氧能力,这可能与双亲遗传物质在子代中发生了重组有关。另外,研究结果显示杂交子代个体间的遗传相似系数为0.709、遗传距离为0.344,这可能与父、母本遗传物质在不同个体中的重组率不同有关。
通过染色体核型分析和AFLP扩增分析,证明了真鲷(♀)×黑鲷(♂)的杂交子代是同时具有父本和母本遗传物质的异源二倍体,并且父、母本遗传物质在杂交中发生了一定程度的重组,使杂交子代表现出一定的杂交优势,但这些优良性状能否稳定遗传还需要做进一步的追踪研究。本研究为今后更加深入地研究杂交子代的生长、抗逆、抗病等特性提供了理论基础,也为探讨杂交优势的遗传机制提供了参考资料。
[1]徐恭绍,郑澄伟.海产鱼类养殖与增殖[M].济南:山东科学技术出版社,1987:322-345.
[2]江世贵,李加儿,区又君,等.平鲷♀与真鲷♂杂交研究[J].海洋科学,1997(5):33-38.
[3]单保当,洪万树,何大仁.真鲷与黑鲷杂交与多倍体育种系列研究-Ⅰ[J].海洋渔业,1999,21(1):14-15.
[4]蒋宏雷,吴雄飞,沈庞幼.真鲷与黑鲷杂交繁育技术研究[J].渔业现代化,2008,35(6):33-35,60.
[5]姜景腾,吴雄飞,蒋宏雷.低氧胁迫对真鲷(♀)与黑鲷(♂)杂交子一代体内酶活力的影响[J].宁波大学学报(理工版),2010,23(4):10-14.
[6]吴仲庆.水产生物遗传育种学[M].3版.厦门:厦门大学出版社,2000:7-25.
[7]周贺,秦艳杰,周伯文,等.4种扇贝 AFLP分析体系的建立[J].大连水产学院学报,2009,24(S1):56-58.
[8]安丽,刘萍,李健,等.“黄海1号”中国对虾不同世代间的 AFLP分析[J].中国海洋大学学报(自然科学版),2008,38(6):921-926.
[9]Liu Z,Nichols A,Li P,et al.Inheritance and usefulness of AFLP mar kers in channel catfish(Ictaluruspunctatus),blue catfish(I.f urcatus),and their F1,F2,and backcross hybrids[J].Molecular and General Genetics,1998,258(3):260-268.
[10]Agresti J J,Seki S,Cnaani A,et al.Breeding new strains of tilapia:develop ment of an artificial center oforigin and linkage map based on AFLP and microsatellite loci[J].Aquaculture,2000,185(1/2):43-56.
[11]王伟继,孔杰,董世瑞,等.中国明对虾 AFLP分子标记遗传连锁图谱的构建[J].动物学报,2006,52(3):575-584.
[12]任军,黄路生,艾华水,等.24个中外猪种(群)的 AFLP多态性及其群体遗传关系[J].遗传学报,2002,29(9):774-781.
[13]Lallias D,Lapègue S,Hecquet C,et al.AFLP-based genetic linkage maps of the blue mussel(Mytilusedulis)[J].Ani mal Genetics,2007,38(4):340-349.
[14]Guo J,Jiang R H Y,Kamphuis L G,et al.A c DNA-AFLP based strategy to identify transcripts associated with avirulence inPhytophthorainf estans[J].Fungal Genetics and Biology,2006,43(2):111-123.
[15]Levan A,Fredga K,Sandberg A A.Nomenclature for centromeric position on chromosomes[J].Hereditas,1964,52(2):201-220.
[16]Cor man G C.The chromosomes of the Reptilia,a cytotaxonomic interpretation[C]//Chiarelli A B,Capanna E.Cytotaxonomy and Vertebrate Evolution.New York:Academic Press Inc,1973:5-30.
[17]王志勇,王艺磊,林利民,等.利用AFLP指纹技术研究中国沿海真鲷群体的遗传变异和趋异[J].水产学报,2001,25(4):289-293.
[18]严骏骢,赵金良,李思发,等.鳙中国土著群体与移居群体遗传变异的AFLP分析[J].中国水产科学,2011,18(2):283-289.
[19]王晓清,王志勇,谢中国,等.大黄鱼(♀)与鮸(♂)杂交的遗传分析[J].水产学报,2008,32(1):51-57.
[20]黄帆.高加索三叶草与白三叶杂交后代遗传特性及育性恢复的研究[D].呼和浩特:内蒙古农业大学,2012.
[21]周丽清,杨爱国,刘志鸿,等.栉孔扇贝×虾夷扇贝杂交子一代与双亲染色体核型的分析[J].水生生物学报,2005,29(1):105-109.
[22]杨爱国,王清印,刘志鸿,等.栉孔扇贝与虾夷扇贝杂交及子一代的遗传性状[J].海洋水产研究,2004,25(5):1-5.
[23]隋班良,蔡明夷,刘颖,等.黄姑鱼♀与大黄鱼♂杂交试验与AFLP分析[J].集美大学学报(自然科学版),2012,17(4):241-246.
[24]万俊芬,包振民,刘广绪,等.扇贝种间单对杂交一代幼虫ISSR标记的分离方式[J].高技术通讯,2004,14(5):82-87.
[25]何斌,杨爱国,王清印,等.栉孔扇贝×虾夷扇贝单对杂交子一代幼虫ISSR标记的分离方式[J].海洋水产研究,2007,28(6):32-38.
[26]Ayliffe M A,Lawrence G J,Ellis J G,et al.Heteroduplex molecules for med bet ween allelic sequences cause nonparental RAPD bands[J].Nucleic Acids Research,1994,22(9):1632-1636.
[27]房经贵,章镇,马正强,等.AFLP标记在两个芒果品种间杂交F1代的多态性及分离方式[J].中国农业科学,2000,33(3):19-24.
[28]姜景腾.真鲷♀×黑鲷♂杂交子一代抗逆性初步研究[D].宁波:宁波大学,2011.
