长江口及邻近海域浮游植物现存量的上下行控制分析
2014-10-27宋洪军张学雷王保栋孙霞王晓辛明
宋洪军,张学雷*,王保栋,孙霞,王晓,辛明
(1.国家海洋局 第一海洋研究所,山东 青岛266061;2.海洋生态环境科学与工程国家海洋局重点实验室,山东 青岛266061)
1 引言
根据营养级联关系(Cascading Trophic Interactions)[1],浮游植物现存量主要受营养盐水平与光照条件的上行控制(Bottom-up Control)和藻类摄食者的下行控制(Top-down Control)两效应影响[2-3]。上行效应一般是指浮游植物生长受环境条件影响,下行效应主要是指浮游动物通过摄食作用影响浮游植物生物量的累积。上下行效应对浮游植物现存量控制的相对作用在不同水域和季节是不同的,例如,Sommer[4]现场培养实验研究指出在寡营养的红海北部浮游植物生物量主要受上行效应控制,下行控制作用相对较弱;Goericke[5]研究指出阿拉伯海雨季的高营养盐低叶绿素现象(“high-nutrient,low-chlorophyll”,HNLC)主要受浮游动物的下行控制作用;Thompson等[6]在澳大利亚Huon河口浮游植物水华过程研究发现,冬季该区域硅藻的优势主要受光照条件的上行效应影响,夏季甲藻水华则与潜在的氮限制有关。
近年来,国内学者关于长江口水域浮游植物的营养盐限制研究已开展了较多的工作,并形成了一定的科学认识。研究结果显示,长江口及邻近水域溶解无机氮(DIN)和硅酸盐(Si O3-Si)的浓度均较高(年平均值分别约为20μmol/L和15μmol/L),而磷酸盐(PO4-P)浓度较低(年平均值约0.5μmol/L)[7];硅氮比值(Si/DIN)大于1或小于1的情况均有,但四季的Si O3-Si浓度远高于2μmol/L的阈值,因此长江口及邻近海域不构成硅限制[8];氮磷比值(DIN/P)变化范围较大,一般高于Redfield比值(16∶1),但PO4-P浓度基本也都高于0.2μmol/L,在浮游植物生长的限制性阈值(0.1μmol/L)之上[9];但长期变化来看,DIN浓度的持续升高以及PO4-P浓度的持续低值致使DIN/P的升高趋势显著[10],导致长江口及邻近海域浮游植物生长存在潜在的磷限制。光照条件对长江口浮游植物生长的影响也已引起大家重视,例如,方涛[11]通过现场培养实验研究了不同光照条件下长江口水域浮游植物生长和营养盐吸收作用机制以及不同浮游生物类群对碳贡献率的变化;孙百晔[12]通过对长江口及邻近海域浮游植物生物量平面分布、垂直分布和年代变化光照效应的综合分析,证实光照是影响该海域赤潮发生的重要因素。而浮游动物对浮游植物的下行控制研究目前主要集中在研究区域水华过程中的摄食实验方面[13-14],对浮游植物现存量方面的影响分析尚未见报道。因此,针对控制长江口及邻近海域浮游植物现存量的上下行效应仍需开展综合研究。
广义相加模型(Generalized Additive Model,GAM)是一种基于数据分析的数学模型,数据决定着响应变量与预测因子间的关系,而不是根据影响机制所假设的某种参数关系[15]。该模型能有效揭示响应变量与预测因子间非线性和非单调潜在联系,在生态学和海洋学研究中已经广泛应用。国际上已有利用GAM 模型在湖泊[16]、海湾[17]与海峡[18]等区域研究叶绿素与不同环境因子间关系的报道,国内研究则在异养细菌[19]、浮游动物[20]、渔业资源[21-22]等与环境因子间关系的研究中引入了GAM模型分析方法。
本文利用2010-2011年度的调查资料研究了长江口及邻近海域营养盐、光照(悬浮物)和浮游动物对浮游植物现存量(以叶绿素a浓度表征)空间分布和季节变化的影响,以GAM模型为工具量化分析了影响叶绿素时空变化的主要环境因子,探讨了上下行效应对该区域浮游植物现存量的控制,以期丰富对长江口及邻近海域浮游植物生态学的认识,为生态系统动力学研究提供数据支撑。
2 材料与方法
2.1 数据来源
长江口及邻近海域现场调查站位分布如图1所示。共执行3个航次,调查时间分别为2010年8月(夏季)、2010年11月(秋季)和2011年5月(春季)。样品采集、处理和测定参照《海洋调查规范》[23-24]和《海洋监测规范》[25]执行,其中叶绿素a(表征浮游植物现存量)根据萃取荧光法以Turner-Designs 700型荧光计测定,温盐由RBR(XR-620)仪器测定,营养盐使用SKALAR自动分析仪测定(其中DIN为NO3-N、NO2-N和NH4-N总和),悬浮物采用重量法测定,浮游动物生物量指浅水Ⅰ型浮游生物网采样品的湿重生物量(含胶质浮游动物)。
图1 2010—2011年长江口及邻近海域调查采样站位分布(n=24)Fig.1 Sampling stations off the Changjiang Estuary in 2010—2011(n=24)
2.2 GAM 模型
利用R程序包mgcv中的gam函数[26]模拟研究海域叶绿素对各环境因子的响应。环境因子包括温度(表层,SST)、盐度(表层,SSS)、悬浮物(TSS)、DIN、PO4-P、Si O3-Si和浮游动物生物量(ZB)。由于叶绿素值属于高度右偏态分布,分析时取log10(Chl)转换以达到近似正态分布[27],同理对于浮游动物生物量和TSS也采取对数转换。分析时采取步退式模型选择方法(根据统计学p值和R2值以及模型广义交叉验证分值即GCV值,逐步减少变量),得到最适模型及其响应因子(即影响叶绿素变化的主要环境因子)。
3 结果
3.1 现场观测结果
2010—2011年度三航次叶绿素观测值在0.28~32.01μg/L之间,平均值2.75μg/L。季节比较以夏季叶绿素值最高,春季次之,秋季最低,平均值依次为:5.54μg/L、1.92μg/L、0.78μg/L。春季,叶绿素高值区主要分布在122.5°E以东海域(>3.0μg/L,图2a);夏季,在长江口门外南北两侧形成两个高值区(>5.0μg/L,图2b);秋季,叶绿素浓度普遍较低,仅在3个站位稍高于1.0μg/L(图2c)。叶绿素高值区的季节分布与孙军等[28]报道相似。
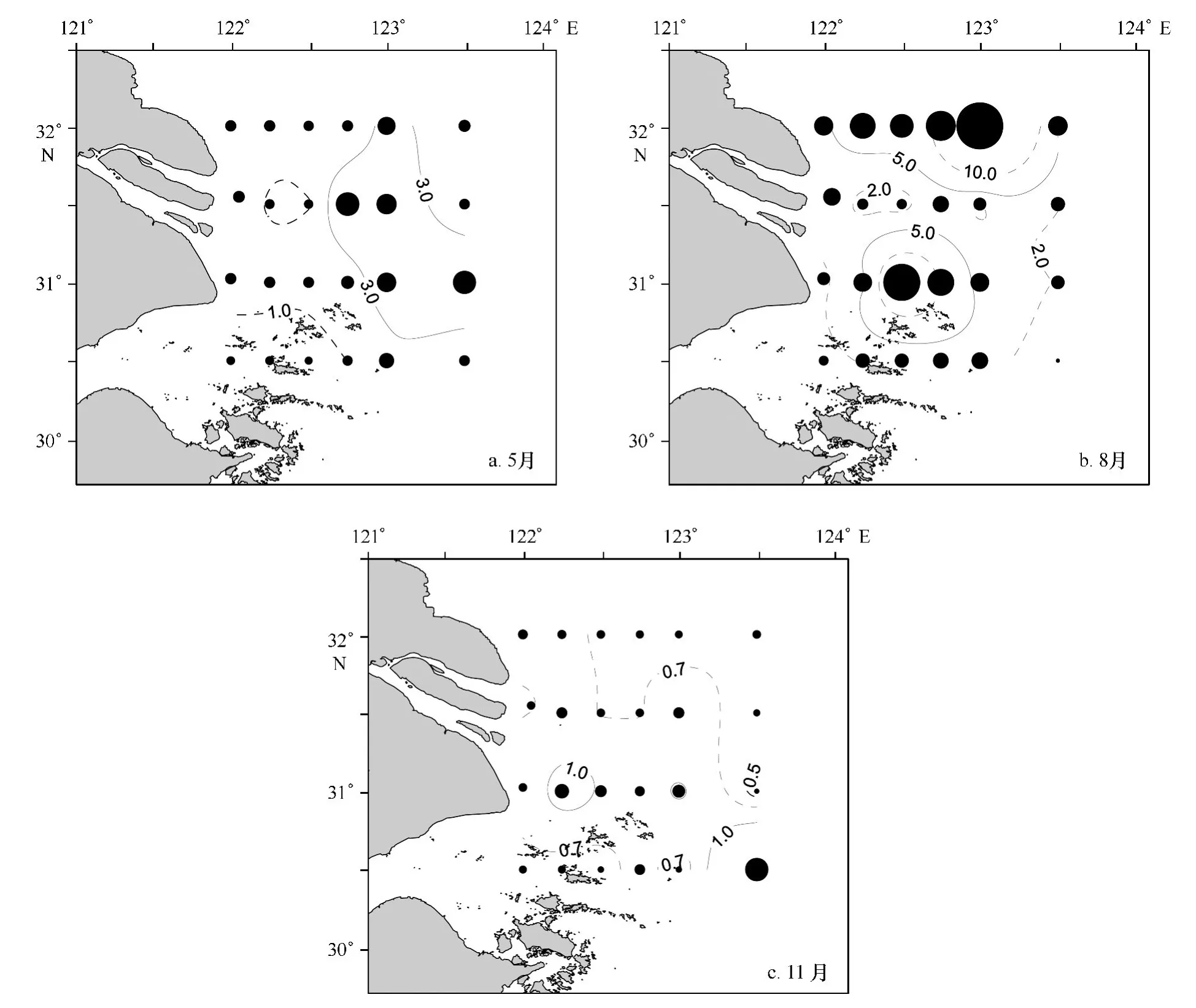
图2 2010—2011年度长江口及邻近海域叶绿素平面分布Fig.2 Spatial distribution of chlorophyll concentration off the Changjiang Estuary in 2010—2011
同航次环境因子的观测值(表1)显示:春季,与整个调查区相比,叶绿素高值区的温度平均值相同,盐度稍高,TSS显著低值,而营养盐(DIN、PO4-P和Si O3-Si)也相对较低,且PO4-P和Si O3-Si浓度在3个季节中最低;夏季,叶绿素高值区的温度相对整个调查区几乎无变化,盐度稍低(不同于春季),TSS显著低值,而3项营养盐也低于全调查区;秋季叶绿素高值区温度、盐度和TSS的平均值较之全调查区稍高,而3项营养盐在叶绿素高值区的平均值相对较低(这与春夏两季类似)。DIN/P在全调查区春夏秋三季节依次为37.4、34.7、19.7,而在叶绿素高值区依次为43.3、32.9、19.2;Si/DIN在全调查区春夏秋三季节依次为0.44、0.55、0.76,而在叶绿素高值区依次为0.43、0.61、0.76。这说明叶绿素高值区的DIN/P在春季相对全调查区较高,而在夏季和秋季稍低,Si/DIN仅在夏季相对较高,春季和秋季则与全调查区基本相同。
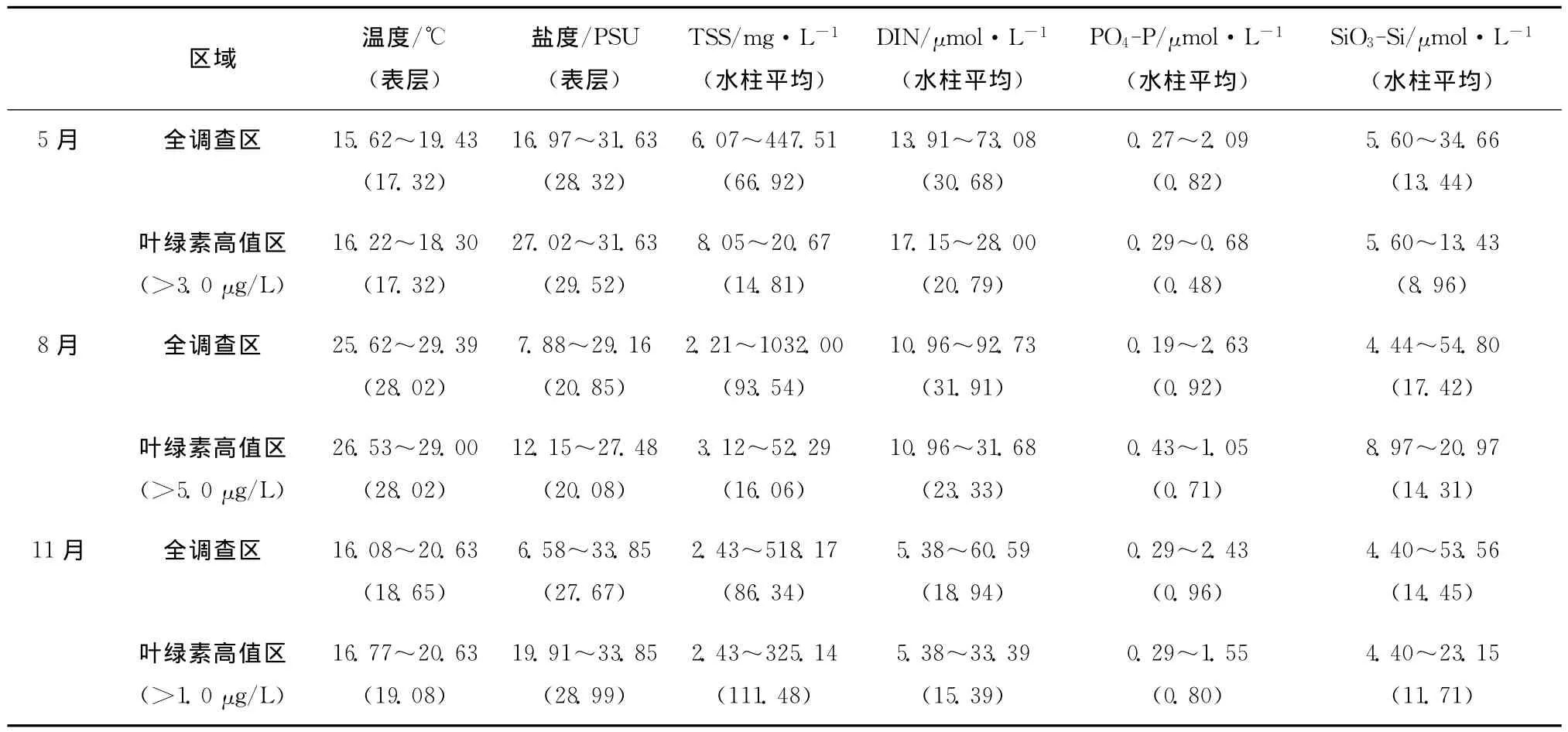
表1 2010—2011年长江口及邻近海域环境因子观测值Tab.1 Variations of environmental factors off the Changjiang Estuary in 2010—2011
浮游动物湿重生物量三航次观测值在1.25~2 563.79 mg/m3之间,平均值为362.62 mg/m3。季节比较来看,春季浮游动物生物量最高,夏季次之,而秋季最低,平均值依次为:571.54 mg/m3、371.31 mg/m3、145.02 mg/m3。春季(如图3a所示),浮游动物高值区(>1 000 mg/m3)分布在调查区中央的B4和C4站位及西北部的A2站位,其中B4站位亦为叶绿素高值区,C4站位靠近叶绿素高值区;夏季(如图3b所示),浮游动物高值区(>1 000 mg/m3)分布在调查区中央的B3和C3站位,其中C3站位处于叶绿素高值区内,而B3站位处于两个叶绿素高值区之间;秋季(图3c),浮游动物相对高值区(>300 mg/m3)处于近岸的B1和C1站位以及中央的C4站位,亦与叶绿素高值区的C2和C5站位相邻,且浮游动物生物量在3个季节中的相对低值与叶绿素趋势一致。
3.2 GAM模型结果
GAM模型结果(见表2)显示:温度(SST)、盐度(SSS)、悬浮物(TSS)、营养盐(DIN、PO4和Si O3)和浮游动物生物量(ZB)变化对叶绿素时空变化的贡献合计可达74.6%(模型1),依次去除统计学意义上不显著(p>0.05)的TSS(模型2)、浮游动物生物量(模型3)和温度(模型4)后拟合结果仅有微小变化,其中GCV值仅下降0.001,而R2最大降低不足0.03;去除统计学意义上显著(p<0.05)的盐度后R2显著降低为0.535,GCV值也升至0.099(模型5),继续去除DIN后,R2显著降低为0.428,GCV值也升至0.118(模型6)。这表明上述因素中影响长江口及邻近海域叶绿素变化的主要因子为盐度和营养盐(DIN、PO4-P和Si O3-Si三者共同作用),而叶绿素变化与TSS、浮游动物生物量和温度三因子的直接相关性不显著。
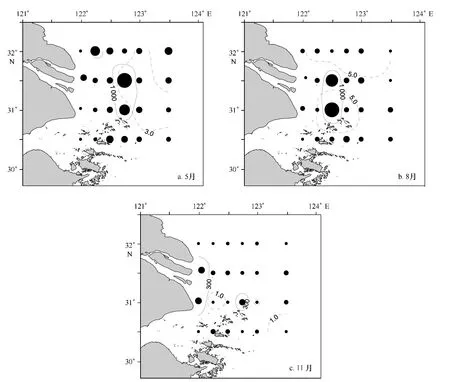
图3 2010—2011年度长江口及邻近海域浮游动物生物量平面分布Fig.3 Spatial distribution of zooplankton biomass off the Changjiang Estuary in 2010—2011

表2 GAM模型统计结果(样本数n=72)Tab.2 Statistical results of generalized additive models(samples number:n=72)
图4示出了主要影响因子(盐度、DIN、PO4-P和Si O3-Si)与叶绿素变化间的关系:叶绿素(对数值)变化随着盐度和PO4-P升高呈逐渐下降的趋势;DIN浓度较低时对应的叶绿素变化无明显规律,在DIN浓度约32μmol/L时叶绿素达到一个峰值,然后降低且在DIN约42μmol/L时至叶绿素最低,随着DIN的继续升高,叶绿素亦有升高趋势,但由于数据点较少以及置信区间较为离散导致结果可信度较低;Si O3-Si与叶绿素变化间的明显规律为在Si O3-Si浓度约22μmol/L时叶绿素表现为一高值,Si O3-Si较低时叶绿素值较低,而Si O3-Si较高时叶绿素值变化较大。此外,受长江冲淡水的影响,研究区域的盐度与DIN、PO4-P、Si O3-Si等营养盐及TSS之间存在显著的相关关系(p<0.001,见图5)。这属于GAM模型中的变量间共相关问题,从直接联系上分析应该去除盐度因子,但去除盐度后模型结果变化较大(如表2模型5所示)。这说明盐度对叶绿素变化的影响可能体现了营养盐和光照条件等因子的作用。
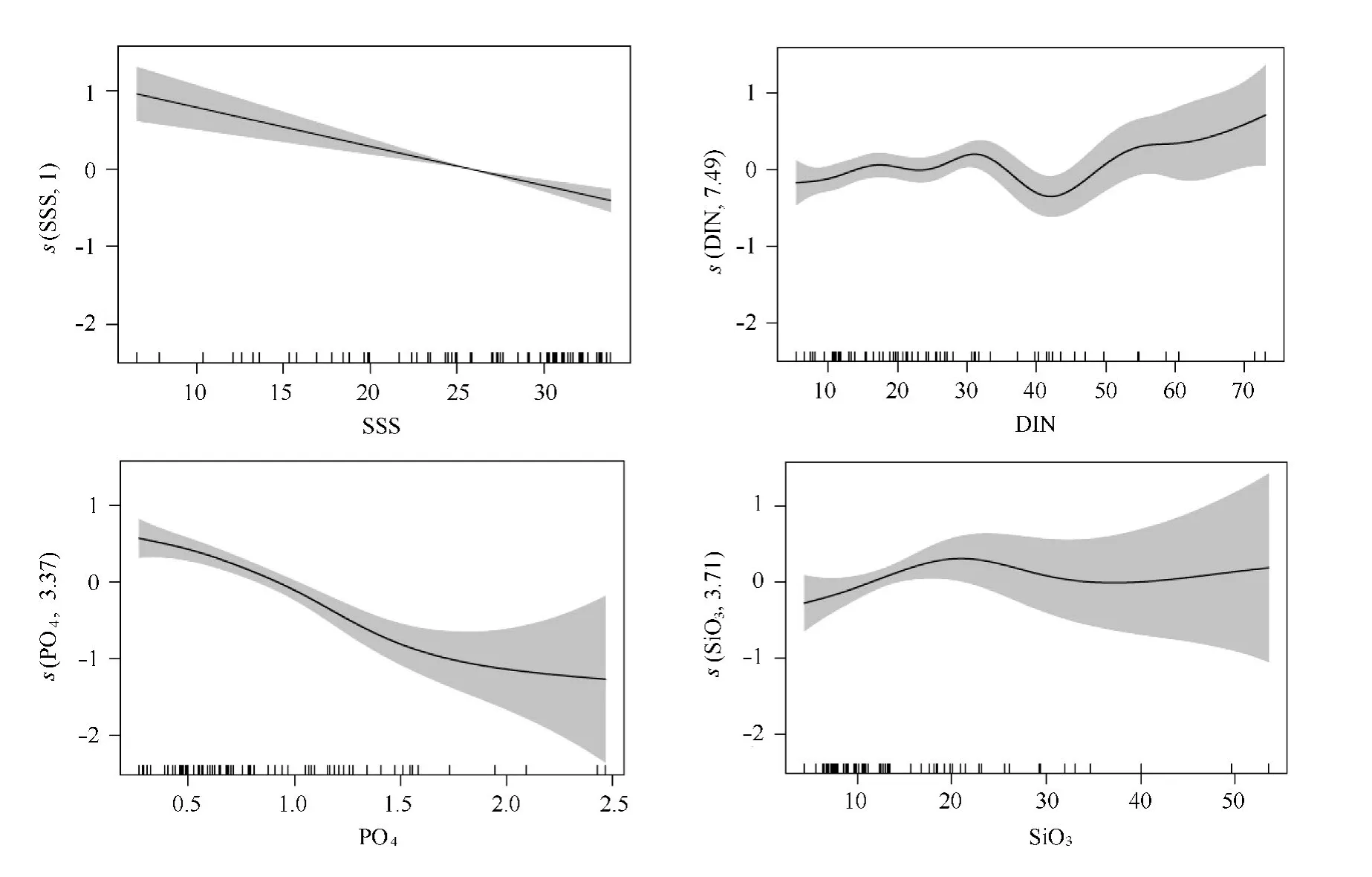
图4 长江口及邻近海域叶绿素变化主要影响因子分析Fig.4 Partial effects of the main environmental factors on the chlorophyll variability off the Changjiang Estuary
4 讨论
水体叶绿素浓度主要取决于浮游植物生长率、垂向扩散与混合、水平移动、沉降损失与被摄食消耗等因素[29]。浮游植物的生长率主要受营养盐和光照的上行效应影响,而被摄食消耗则以浮游动物的下行效应为主。本文观测结果发现,长江口及邻近海域3个季节叶绿素高值区营养盐相对全区平均值较低,夏春两季叶绿素高值区(也是全年叶绿素高值)的水体TSS相对较低,夏季叶绿素高值区盐度较低,而温度在叶绿素高值区差别不大;浮游动物生物量高值区与叶绿素高值区未体现出较好的对应关系,但两者基本都有交叉或位置邻近。GAM模型结果进一步显示,营养盐是影响长江口及邻近海域叶绿素变化的主要环境因子,盐度在解释叶绿素变化趋势上亦起了较大作用(由于盐度与营养盐和TSS显著相关,导致盐度体现了营养盐和光照条件等因子的共同作用),而浮游动物生物量和温度等因子直接影响不显著。

图5 长江口及邻近海域盐度与营养盐和悬浮物之间的相关性(n=72)Fig.5 The correlations bet ween surface salinity and nutrients &total suspended solids off the Changjiang Estuary(n=72)
长江冲淡水低盐、高浑浊度和高营养盐的特性,导致了长江口及邻近海域形成了盐度、TSS、营养盐等一系列梯度变化,而光照条件和营养盐含量达到最佳平衡时往往形成浮游植物高生物量区[30-31]。韩秀荣[7]利用2002—2007年的调查资料统计得知叶绿素高值发生时(最大值可达70μg/L)的DIN、PO4-P以及浊度均维持在较低的水平,而Si O3-Si浓度约在20μmol/L、盐度约30 PSU以及温度约为20℃时叶绿素浓度最高。本文GAM模型中的PO4-P和Si O3-Si结果(PO4-P浓度较低,Si O3-Si浓度约22μmol/L时叶绿素表现为一高值)与其相一致,但与DIN和盐度模拟结果存在差异(图4),这可能与两者研究区域和时间上的不同(韩秀荣[7]研究区域往南延伸,研究时间以春季为主)有关。本文引入的GAM模型分析方法在量化上具有较大的优势,在甄别影响浮游植物现存量的多环境因子不同作用方面具有很好的效果。
长江口及邻近海域近年来已成为赤潮频发区,春季赤潮藻类群以甲藻为主(米氏凯伦藻、链状亚历山大藻、具齿原甲藻等)[32],这与水体 PO4-P含量较低以及较高的DIN/P有关。李瑞香等[33]的船基围隔实验结果表明低磷条件下具齿原甲藻生长比中肋骨条藻生长迅速且持续时间长;孙军等[34]研究表明米氏凯伦藻在氮磷比为80∶1的条件下比生长率最快;吕颂辉和李英[35]研究结果显示具齿原甲藻和链状亚历山大藻较之中肋骨条藻和尖刺拟菱形藻具有较高的细胞氮磷营养储存能力,属于k-策略者,有助于甲藻赤潮的长时间持续。本文研究结果亦表明春季叶绿素高值区在空间和季节比较上均具有较低的PO4-P含量和较高的DIN/P比值,为该海域春季甲藻赤潮频发提供了有利条件。另一方面,夏季叶绿素高值区的Si/DIN在空间比较上比值较高(见3.1部分),这与夏季藻华种类主要是硅藻(优势种为中肋骨条藻和细长翼鼻状藻[36])相一致。长期变化来看(近50年),DIN的升高和Si O3-Si的降低导致DIN/P和DIN/Si比值均呈升高的趋势,也是近年来赤潮优势种由硅藻演变为甲藻的主要原因[10]。
长江口及邻近海域水体的高浑浊度使光照条件成为影响浮游植物生长的重要环境因子。本文观测结果显示,春、夏两季叶绿素高值区的TSS浓度在时空比较上均显著低值(见表1),说明较好的光照条件是浮游植物形成高生物量的一个重要条件。李雁宾[37]通过环境因子变异系数研究指出光照对硅藻赤潮的季节性生消影响最大,光照条件的增大有助于硅藻在与甲藻的竞争中占优;孙百晔[12]利用生长光照效应分析及模拟证实了海水中的光照条件是限制赤潮进一步向近岸方向扩展的关键因素。但GAM模型分析结果显示,TSS与叶绿素变化之间的直接相关性不显著,说明光照条件对长江口及邻近海域浮游植物现存量的控制作用不起决定性作用。
本文观测结果和模型分析显示,长江口及邻近海域浮游动物生物量和叶绿素浓度之间相关性不显著,这表明浮游动物生物量在影响浮游植物现存量时空变化方面不是决定性因子。现场实验结果显示,微型浮游动物在靠岸站位对浮游植物现存量的摄食压力相对较高[13],而在水华爆发前后以及水华区和非水华区微型浮游动物比摄食率各有不同[14]。另一方面,营养传递导致的浮游动物生物量升高或下降相对浮游植物现存量变化存在一定的滞后性,往往导致两者之间直接相关性不显著。高的浮游植物现存量可为浮游动物提供足够食物,而浮游动物生物量升高会反过来加剧对浮游植物的摄食,使浮游植物现存量下降;另一方面,浮游植物低生物量时,可能是环境条件不适宜浮游生物的生长(此时浮游动物生物量也低),也可能是较高的浮游动物摄食作用所导致。此外,浮游动物相对浮游植物一般具有较强的水平或垂直移动能力,也可能影响两者之间的直接联系。因此,就单一或某几个航次结果来看,很难找出浮游动物和浮游植物现存量之间较好的相关关系。
因此,在长江口及邻近海域,营养盐的上行效应是浮游植物现存量的主要控制作用,而光照条件和浮游动物生物量与浮游植物现存量虽然在时空分布上存在一定的联系,但非决定性控制因素。
[1]Carpenter S R,Kitchell J F,Hodgson J R.Cascading trophic interactions and lake productivity[J].BioScience,1985,35(10):634-639.
[2]Mc Queen D J,Post J R,Mills E L.Trophic relationships in freshwater pelagic ecosystems[J].Canadian Journal of Fisheries and Aquatic Sciences,1986,43(8):1571-1581.
[3]Leh man J T.Interacting growth and loss rates:the balance of top-down and bottom-up controls in plankton communities[J].Limnology and Oceanography,1991,36(8):1546-1554.
[4]Sommer U.Scarcity of medium-sized phytoplankton in the northern Red Sea explained by strong bottom-up and weak top-down control[J].Marine Ecology Progress Series,2000,197:19-25.
[5]Goericke R.Top-down control of phytoplankton biomass and community structure in the monsoonal Arabian Sea[J].Li mnology and Oceanography,2002,47(5):1307-1323.
[6]Thompson P A,Bonham P I,Swadling km.Phytoplankton blooms in the Huon Estuary,Tasmania:top-down or bottom-up control?[J].Journal of Plankton Research,2008,30(7):735-753.
[7]韩秀荣.长江口及邻近海域浮游植物生长的多环境效应因子影响解析研究[D].青岛:中国海洋大学,2009.
[8]王保栋.长江口及邻近海域富营养化状况及其生态效应[D].青岛:中国海洋大学,2006.
[9]杨东方,王凡,高振会,等.长江口理化因子影响初级生产力的探索:Ⅱ.磷不是长江口浮游植物生长的限制因子[J].海洋科学进展,2006,24(1):97-107.
[10]王江涛,曹婧.长江口海域近50a来营养盐的变化及其对浮游植物群落演替的影响[J].海洋环境科学,2012,31(3):310-315.
[11]方涛.光照和N、P营养盐的共同作用对长江口浮游植物生长的影响[D].上海:华东师范大学,2008.
[12]孙百晔.长江口及邻近海域浮游植物生长的光照效应研究[D].青岛:中国海洋大学,2008.
[13]孙军,刘东艳,王宗灵,等.春季赤潮频发期东海微型浮游动物摄食研究[J].应用生态学报,2003,14(7):1073-1080.
[14]孙军,宋书群.东海春季水华期浮游植物生长与微型浮游动物摄食[J].生态学报,2009,29(12):6429-6438.
[15]Hastie T,Tibshirani R.Generalized additive models[J].Statistical Science,1986,1(3):297-310.
[16]Lamon III E C,Reckhow K H,Havens K E.Using generalized additive models for prediction of chlorophyllain Lake Okeechobee,Florida[J].Lakes &Reservoirs:Research and Management,1996,2(1/2):37-46.
[17]Raitsos D E,Korres G,Triantafyllou G,et al.Assessing chlorophyll variability in relation to the environmental regime in Pagasitikos Gulf,Greece[J].Journal of Marine Systems,2012,94:S16-S22.
[18]Tew-Kai E,Marsac F.Patterns of variability of sea surface chlorophyll in the Mozambique Channel:A quantitative approach[J].Journal of Marine Systems,2009,77(1/2):77-88.
[19]Chen B Z,Liu H B,Huang B Q.Environmental controlling mechanisms on bacterial abundance in the South China Sea inferred from generalized additive models(GAMs)[J].Journal of Sea Research,2012,72:69-76.
[20]Chen B Z,Landry M R,Huang B Q,et al.Does warming enhance the effect of microzooplankton grazing on marine phytoplankton in the ocean?[J].Li mnology and Oceanography,2012,57(2):519-526.
[21]郑波,陈新军,李纲.GL M和GA M模型研究东黄海鲐资源渔场与环境因子的关系[J].水产学报,2008,32(3):379-386.
[22]朱国平.基于广义可加模型研究时间和环境因子对南极半岛北部南极磷虾渔场的影响[J].水产学报,2012,36(12):1863-1871.
[23]GB/T 12763.6-2007,海洋调查规范 第6部分:海洋生物调查[S].北京:中国标准出版社,2008.
[24]GB/T 12763.4-2007,海洋调查规范 第4部分:海水化学要素调查[S].北京:中国标准出版社,2008.
[25]GB 17378.4-2007,海洋监测规范 第4部分:海水分析[S].北京:中国标准出版社,2008.
[26]Wood S N.Stable and efficient multiple smoothing parameter estimation for generalized additive models[J].Journal of the American Statistical Association,2004,99(467):673-686.
[27]Campbell J W.The lognor mal distribution as a model for bio-optical variability in the sea[J].Journal of Geophysical Research:Oceans,1995,100(C7):13237-13254.
[28]孙军,宋书群,栾青杉.长江口水域浮游植物生物量及结构特点[M]//俞志明,沈志良,陈亚瞿,等.长江口水域富营养化.北京:科学出版社,2011:184-203.
[29]Banse K.Grazing and zooplankton production as key controls of phytoplankton production in the open ocean[J].Oceanography,1994,7(1):13-20.
[30]宁修仁,史君贤,蔡昱明,等.长江口和杭州湾海域生物生产力锋面及其生态学效应[J].海洋学报,2004,26(6):96-106.
[31]孙军,宋书群,栾青杉.长江口水域浮游植物关键过程及其对富营养化的影响[M]//俞志明,沈志良,陈亚瞿,等.长江口水域富营养化.北京:科学出版社,2011:336-349.
[32]周名江,朱明远.“我国近海有害赤潮发生的生态学、海洋学机制及预测防治”研究进展[J].地球科学进展,2006,21(7):673-679.
[33]李瑞香,朱明远,王宗灵,等.东海两种赤潮生物种间竞争的围隔实验[J].应用生态学报,2003,14(7):1049-1054.
[34]孙军,刘东艳,陈宗涛,等.不同氮磷比率对青岛大扁藻、新月柱鞘藻和米氏凯伦藻生长影响及其生存策略研究[J].应用生态学报,2004,15(11):2122-2126.
[35]吕颂辉,李英.我国东海4种赤潮藻的细胞氮磷营养储存能力对比[J].过程工程学报,2006,6(3):439-444.
[36]孙军,宋书群,栾青杉.长江口水域浮游植物群集组成结构与特点[M]//俞志明,沈志良,陈亚瞿,等.长江口水域富营养化.北京:科学出版社,2011:144-184.
[37]李雁宾.长江口及邻近海域季节性赤潮生消过程控制机理研究[D].青岛:中国海洋大学,2008.
