microRNA657与新疆维吾尔族高脂血症相关性研究
2014-07-13王丽凤毛新民米娜瓦尔胡加艾朱曼丽李琳琳
杜 宪, 焦 谊, 龙 梅, 王丽凤, 毛新民, 米娜瓦尔·胡加艾, 武 云,朱曼丽, 李琳琳, 王 烨
(新疆医科大学1基础医学院生物化学教研室, 2基础医学院机能教研室, 3基础医学院生理学教研室,4中医学院,5第一附属医院综合内科, 6协同实验中心, 7基础医学院药理学教研室, 乌鲁木齐 830011)
高脂血症是各种原因导致的血浆中胆固醇和(或)甘油三酯水平升高的一种疾病,是引发和加重动脉粥样硬化的病理基础及导致动脉硬化、心绞痛、心肌梗死、脑梗塞、肾损害等动脉栓塞性疾病的独立危害因素[1]。我国高脂血症的发生以高甘油三酯居多,主要与饮食、体质指数(BMI)和生活方式关系较大[2]。孙玉萍等[3]调查新疆维吾尔族高脂血症的分析研究发现:随年龄增加,维吾尔族人群脂代谢水平紊乱与高脂血症的患病率增加,而高脂血症主要表现为高甘油三酯血症和高胆固醇血症患病率增加。近几年的研究发现,微小RNA(microRNA)与脂代谢调节密切相关,microRNA对脂代谢的调节是一个复杂的网络调节的过程,不同microRNA之间的靶点具有重叠性,相关的作用机制涉及到胆固醇、甘油三酯、脂肪酸等脂质的合成、代谢和转化。microRNA生物学功能是在转录后水平调节负调控基因的表达,其发生在mRNA水平和蛋白质水平。因此本研究采用前期芯片初筛结果及实时荧光定量PCR方法验证,初步探讨microRNA657与维吾尔族高脂血症的关系。
1 材料与方法
1.1材料
1.1.1 主要试剂与仪器 异丙醇,氯仿(SIGMA),TRIzol REAGENT BD(MRC),miScript SYBR®Green PCR kit、miScript Primer Assays、10×miScript Universal Primer microRNANeasy mini KitmiScript Ⅱ RT Kit (德国Qiagen公司)。涡旋混合器,低温高速离心机,实时荧光定量PCR热循环仪购自ABI公司。
1.1.2 标本采集 样本采自新疆医科大学第一附属医院内分泌VIP二病区维吾尔族高脂血症组和血脂正常组抗凝全血样本各14例。样本均为受调查者早晨空腹的新鲜抗凝血样。本方案经医院伦理委员会批准,患者签署知情同意书。高脂血症纳入标准:血脂水平参照中国成人血脂异常防治指南规定的标准[4],总胆固醇(TC)浓度≥5.18 mmol/L,甘油三酯(TG)浓度≥1.7 mmol/L,高密度脂蛋白(high-density lipoprotein,HDL)浓度<1.04 mmol/L,低密度脂蛋白(low-density lipoprotein,LDL)浓度≥3.37 mmol/L。排除标准:(1)妊娠或哺乳期妇女;(2)半年内曾患急性心肌梗死、脑血管意外、严重创伤或重大手术后的患者;(3)在被调查期间患有感冒、急性胃肠炎等急性疾病而对证候判断产生干扰者。
1.2方法
1.2.1 临床指标的测定 受试者脱鞋、免冠,测量身高、体质量、臀围、腰围,并计算体质指数(BMI)。测量2次收缩压与舒张压,取平均值;采集肘静脉血5 mL,分别测空腹血糖(FPG)、甘油三酯(TG)、总胆固醇(TC)、高密度脂蛋白胆固醇(HDL-C)、低密度脂蛋白胆固醇(LDL-C)等生化指标。
1.2.2 抗凝血中提取总RNA及逆转录 用TRI REAGENT BD的方法提取总RNA样品;在冰上准备反转录体系,根据miScript Ⅱ RT Kit试剂盒说明书进行逆转录,在37℃孵育60 min及95℃孵育5 min的条件下合成单链cDNA。
1.2.3 实时荧光定量PCR 取逆转录产物cDNA模板2 μL,配制PCR总反应体系20 μL。参照miScript SYBR®Green PCR kit试剂盒说明书操作,进行实时荧光定量PCR扩增。反应条件:预变性95℃ 15 min,变性94℃ 15 s,退火55℃ 30 s,延伸70℃ 35 s,共40个循环。每个样本重复2次。

2 结果
2.1维吾尔族高脂血症组和血脂正常组间体检及生化指标的比较维吾尔族高脂血症组患者年龄、FPG、TG水平高于血脂正常组,HDL-C水平低于血脂正常组,差异有统计学意义(P<0.05)。两组其余指标比较差异均无统计学意义,见表1、2。

表1 维吾尔族高脂血症和血脂正常组间体检指标的比较
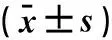
表2 维吾尔族高脂血症和血脂正常组间生化指标的比较
2.2实时RT-PCR检测microRNA657在维吾尔族高脂血症组和血脂正常组抗凝全血中的表达实时RT-PCR分析结果显示:内参U6和microRNA657的扩增曲线展示了RT-PCR产物好的扩增效率;其熔解曲线为单峰,出峰位置为U6和microRNA657的退火温度,特异性好。microRNA657在维吾尔族血脂正常组的表达量为(2.664±0.73),较高脂血症组的表达量(0.883±0.32)显著升高,差异具有统计学意义(t=2.237,P=0.038),见图1~5。

图1 microRNA657在维吾尔族血脂正常组和高脂血症组表达量的比较

图2 内参U6熔解曲线

图3 内参U6扩增曲线

图4 microRNA657 熔解曲线

图5 microRNA657 扩增曲线
2.3microRNA657与维吾尔族高脂血症组和血脂正常组临床指标的相关分析在维吾尔族高脂血症组和血脂正常组中,microRNA657与临床的相关指标无相关性(P>0.05)。在控制年龄的影响后,偏相关性分析发现microRNA657的表达量与收缩压呈负相关(r=-0.518,P<0.05),与舒张压呈负相关(r=-0.529,P<0.05),而与身高、体质量、腰围、臀围、腰臀比、FPG、TC、TG、HDL-C、LDL-C等无相关性。提示在控制年龄的影响后,随着microRNA657的表达量增加,收缩压和舒张压的水平会逐渐下降。
3 讨论
高脂血症又称脂质代谢紊乱或异常,是导致脂肪肝、动脉粥样硬化等多种疾病的因素。高血脂症的病因学预防和治疗已经成为目前的研究热点,但至今药物治疗尚未明显突破。而近期研究发现,microRNA与人类脂质代谢及心血管疾病等密切相关[6-7]。microRNAs是一种内源性、高度保守的、大小约为22个核苷酸的单链非编码RNA,通过促进靶mRNA的降解或抑制其翻译而致基因表达沉默[8-10]。据报道人类所有microRNA约占基因总数的3%,可能调控人体细胞中约30%的蛋白质编码基因[11],参与生命中许多复杂的生理过程。
研究发现通过下调microRNA143的表达可以使甘油三酯聚集减少,并且抑制脂肪细胞的分化,下调葡萄糖转运蛋白-4和PPARγ编码基因的表达,间接调控糖、脂的代谢[12]。在肝脏中特异表达的microRNA21可正性调控胆固醇生物合成关键酶的表达,维持正常脂代谢[ 13-14]。microRNA122和microRNA370通过调控脂肪酸的β氧化反应进程调节血脂的代谢[15]。有研究通过向小鼠尾静脉内注射固定剂量的microRNA122拮抗剂发现microRNA122作用被抑制后,实验组小鼠血浆胆固醇水平比对照组降低40%以上[16]。诸多研究表明microRNA与脂代谢息息相关。
目前国内外对微小RNA的研究有很多,但是对microRNA657的研究尚不多。研究发现microRNA657可以调控Hep G2细胞中的IGF2R的表达水平,有助于解释2型糖尿病的发生机制[17]。microRNA657作用于靶基因TLE1通过NFKB途径促进肿瘤发生肝癌[18]。但是目前的研究尚未发现microRNA657与脂质代谢及心血管等疾病相关。本研究通过对维吾尔族高脂血症和血脂正常组的抗凝全血中microRNA表达量的研究发现,microRNA657在维吾尔族的高脂血症和非高脂血症人群中抗凝全血存在差异表达,microRNA657高脂血症组的表达低于血脂正常组(P<0.05)。而Xu等[19]在2003年就发现microRNA与血脂存在关联,将microRNA14沉默后,动物体内的甘油三酯和甘油二酯水平都升高,而增加microRNA14的表达后,血脂水平则恢复正常。关于microRNA657的表达上调是否同样具有降低血脂的作用,有待于在后续的试验中继续研究。
高血脂与高血压在心血管病发病危险中有着重要作用,且二者之间又是相互促进[20]。高血脂人群血清总胆固醇、甘油三酯、低密度脂蛋白胆固醇均可作用于细胞膜,使其通透性改变,并与细胞Ca2+内流呈正相关。Ca2+内流增加导致血管平滑肌收缩增加、外周血管阻力升高,导致血压升高。本研究发现,在偏相关性分析中,microRNA657与收缩压和舒张压呈负相关,说明microRNA657与血压具有相关性,也提示若降低microRNA657在维吾尔族高脂血症组的表达水平,则其血压状况也可能会升高。
另外,本研究实验结束后,又利用targetscan、mirBase和miranda3个靶基因软件综合预测了microRNA657的靶基因,发现有CYTL1、 KIAA1324、PCYT1B等多个靶基因,这些靶基因分别涉及细胞的生长代谢、免疫调节、信号转导、脂代谢等多种功能。目前,用实验方法和计算机生物信息学手段明确了在维吾尔族高脂血症人群microRNA657的差异表达及其靶标,这也为揭示它的功能奠定了基础。因此,在此基础之上探明microRNA657具体调节途径以及作用机制将是对研究维吾尔族高脂血症调控的相关理论提供新的理论依据。后续的试验研究将针对microRNA657的具体作用靶点以及在脂代谢中如何发挥其生物学功能进行深入的研究。
综上所述,本研究分别从新疆维吾尔族高脂血症组和血脂正常组人群的抗凝全血中检测到了microRNA657的差异表达,其表达水平的高低与维吾尔族高脂血症相关。通过对microRNA657的研究,以期在microRNA分子研究水平上,为新一代的调血脂药物提供一个新的切入点。本研究结果为进一步研究microRNA657在高脂血症的作用及其机制奠定了基础,同时也为microRNA657的生物学功能及其作用机制的研究提供了参考依据。
参考文献:
[1] 茹秋梅,张建华.高脂血症的健康教育[J].中国健康教育,2002,18(10):630-631.
[2] 陈灏珠.实用内科学[M].10版.北京:人民卫生出版社,1998:877-890.
[3] 孙玉萍,李清,艾森拉江·买买提艾力,等.维吾尔族居民血脂及高脂血症分析[J].中国公共卫生,2010,26(3):271-272.
[4] 中国成人血脂异常防治指南制定联合委员会.中国成人血脂异常防治指南[J].中华心血管病杂志,2007,35(5):390-419.
[5] Kong L,Zhu J,Han W,et al.significance of serum microRNAs in pre-diabetes and newly diagnosed type 2 diabetes:aclinical study[J].Acta Diabetetol,2011,48(1):61.
[6] Tam STJ,Bastian I,Zhou XF,et al.MicroRNA 143 expression in dorsal root ganglion neurons[J].Cell Tissue Res,201l,346(2):163-173.
[7] 马兰,张基昌,崔燕,等.MicroRNAs与冠心病发生发展关系的研究进展[J].中国实验诊断学,2013,17(7):1354-1357.
[8] Lu Y, Zhang Y, Yang B, et al. MicroRNA-328 contributes to adverse electrical remodeling in atrial fibrillation[J]. Circulation, 2010, 122(23): 2378-2387.
[9] Pillai RS, Bhattacharyya SN, Filipowicz W. Repression of protein synthesis by microRNAs: how many mechanisms[J].Trends Cell Biol, 2007, 17(3): 118-126.
[10] Billonl N,Monteiro MC,Dani C.Developmental origin of adipocytes:new insights into a pending question[J].Biol Cell,2008,100(10):563-575.
[11] Schickel R,Boyerinas B,Park SM,et al.MicroRNAs:key players in the immune system,differentiation,tumorigenesis and cell death[J].Oncogene,2008,27(45):5959-5974.
[12] 凌宏艳,胡小波,奉水东,等.小鼠间充质干细胞脂向分化过程microRNA-143的表达[J].基础医学与临床,2012,32(1):12-15.
[13] Kim YJJ ,Hwang SH,Cho HJ,et al.MicroRNA21 regulates the proliferation of human adipose tissue-derived mesenchymal stem cells and high-fat diet-induced obesity alters microRNA 21 expression in white adipose tissues[J].Cell Physiol,2012,227(1):183-193.
[14] 王伟新,张勇,李晓宇,等.脂肪基质细胞脂向分化过程中MicroRNA21的表达变化[J].现代生物医学进展,2009,9(23):4404-4406.
[15] Iliopoulos D,Drosatos K, Hiyama Y,et al.MicroRNA-370 controls the expression of MicroRNA-122 and Cpt1α and affects lipid metabolism[J].Lipid Res,2010,51(6) :1513-1523.
[16] Kruetzfeldt J, Rajewsky N, Braich R,et al.Silencing of microRNAs in vivo with antagomicroRNAs[J]. Nature,2005,438(7068):685-689.
[17] Lv KL,Guo Y,Zhang Y.Allele-specific targeting of hsa-miR-657 to human IGF2R creates a potential mechanism underlying the association of ACAA-insertion/deletion polymorphism with type 2 diabetes[J]. Biochem Biophys Res Commun,2008,374(1):101-105.
[18] Zhang LL,Yang L,Liu X,et al.microRNA-657 promotes tumorigenesis in hepatocellular carcinoma by targeting transducin-like enhancer protein 1 through nuclear factor kappa B pathways[J]. Hepatology,2013,57(5):1919-1930.
[19] XU Ping,Vernooy SY,Guo M,et al.The Drosophila microRNA14 suppresses cell death and is required for normal fat metabolism [J].Curr Biol,2003,13,(9):790-795.
[20] 王颖,马莉,邹飒枫,等.海岛成年居民高血压现状与高血脂现状分析[J].中国健康杂志,2012,6(3):216.
