表达白介素-23基因的重组纯化及其对妊娠期哮喘气道炎症状态下免疫应答产生的影响
2014-05-19马佳佳NickLu陈必良
马佳佳,Nick Lu,陈必良
(1.第四军医大学西京医院妇产科,陕西西安 710032;2.Department of Allergy-immunology,Northwestern University,USA Chicago 60611)
妊娠合并哮喘是妊娠合并症中一种炎症潜伏与反复发作免疫应答过程尚未明确的常见疾病[1]。由于妊娠期母体特有的生理状态和炎症的持续感染反复迁延,引发免疫应答异常导致的孕期并发症,表现为肺组织病理损伤、胎儿低出生体重、早产危险性[2]等,均对母婴安全和机体免疫系统的自身稳定产生影响。白细胞介素-23(interleukin-23,IL-23)具有前炎症反应活性[3],通过结合靶细胞膜上IL-23受体,依赖致病CD4+T细胞亚群,广泛参与体内免疫过程,调节免疫细胞分化,刺激并特异性促进相关因子表达[4],因在多种肺部疾病炎症形成以及靶组织病理生理反应中的作用而成为近年来的研究热点[5]。本研究利用大肠杆菌表达的 IL-23重组蛋白,通过转导细胞聚集定位和体外转移鉴定,探讨IL-23-IL-23R-IL-17炎性通路激活及其Th细胞线性家族平衡偏移特异性靶分子参与妊娠期哮喘发病的可能机制,为以IL-23为靶向,寻找针对其引发炎症反应临床免疫干预治疗的新策略提供新思路。
1 材料
1.1 主要试剂 pMD18-T、pCEP4、E.coilDH5a、BL21(DE3)、肥大细胞株P815均由美国芝加哥西北大学过敏和免疫学系Nick Lu博士馈赠;pEGFPN1质粒(美国 Invitrogen);T4 DNA连接酶(大连TaKaRa);限制性内切酶、异丙基-β-D-硫代半乳糖苷(isopropylβ-D-1-thiongalactopyranoside,IPTG)(美国 Amresco);Glutathione Sepharose 4B(美国 Pharmacia Biotech);凝胶电泳迁移率变动(EMSA)试剂盒(美国 Pierce);荧光标记抗 FITC-CD4、CD25,抗PE-IFN-γ、IL-4、IL-17A,抗 PE-Cy5-FoxP3 mAb及同型对照(美国 BD/Pharmingen);小鼠 IL-17A ELISA检测试剂盒(武汉博士德生物工程有限公司)。
1.2 动物与标本 SPF级♀ BALB/c小鼠48只,6~8周龄,体质量18~20 g/只(第四军医大学实验动物中心提供);妊娠期哮喘妇女清晨空腹外周血采自第四军医大学西京医院妇产科就诊自愿者。本研究获得医院伦理委员会讨论通过。
2 方法
2.1 目的基因扩增及p40+p19双亚基表达载体构建 根据GeneBank中IL-23 cDNA序列和p40/p19 mRNA参考序列设计引物,并委托上海生工生物工程技术服务公司合成。p19F:5′-GAGCACCGGTCGCACCAGCAACCCTGAGTCCCTA-3′(下划线为KpnⅠ酶切位点,双下划线为表达增强序列),p19R:5′-CAAAGCTCGAGTTCCCTTCCCATCTAATA A-3′(下 划 线为 XhoⅠ 酶 切位 点);p40F:5′-GCTAGTCGATGGTGAGCCGTGAT-3′(下划线为NotⅠ 酶 切 位 点 ),p40R:5′-CATGACCGGCTAATGAGAAAGGGATT-3′(下划线为KpnⅠ酶切位点,双下划线为表达增强序列)。
以 p19F/p19R、p40F/p40R为引物 PCR扩增IL-23基因p19和p40双亚基cDNA全长。反应条件:94℃预变性 5 min;94℃30 s、61℃30 s、72℃1 min,30个循环后72℃延伸10 min。凝胶电泳鉴定并以两者扩增产物为模板,p19F/p40R为引物完成重叠PCR扩增,反应条件同上。将纯化产物分别克隆于pMD18-T载体,PmeⅠ酶切pMD18-T-p19/p40 DNA凝胶回收片段先后插入pCEP4载体,T4连接酶过夜连接后经转化、铺板筛选、挑取克隆菌落扩增并提取质粒,Pac I酶切和测序鉴定最终获得重组pCEP4-IL-23(p19+p40)命名为 pCEP4-IL-23。
2.2 重组蛋白诱导表达及产物纯化鉴定 挑取重组质粒和空载质粒转化E.coilBL21单菌落,于卡那霉素LB液体培养基中220 r·min-1,37℃振摇过夜,至菌液波长A600nm值为0.5~0.6时,取1 ml菌液作为诱导前对照,剩余菌液中加入IPTG终浓度0.5 mmol·L-122℃诱导表达4 h,离心收集超声裂解菌体上清0.22μm滤膜过滤,与预包被GST蛋白的Glutathione Sepharose 4B凝胶颗粒4℃反应过夜,3次重复上样。以预冷PBS洗脱未结合蛋白15 min×4次,收集梯度透析复性过滤产物进行SDS-PAGE电泳和蛋白质印迹鉴定。
2.3 重组蛋白介导转染及结合效应检测 对数生长期P815细胞以1×106/孔接种24孔板,37℃培养过夜至细胞丰度达80%~90%。转染前1 h吸去培养液,用5%葡萄糖和去离子水分别溶解纯化的重组蛋白与pEGFP-N1质粒,调浓度0.2 g·L-1。蛋白/质粒按 0、0.5、1.0、3.0、5.0、7.0不同质量比轻柔混均反应24 h,37℃水浴后将收集的蛋白/DNA复合液瞬时转入实验细胞继续培养,每一质量比设3个平行孔。12 h后加入1 mg·L-1LPS,设LPS刺激前为0 d。分别于转染后d 2、d 3、d 4荧光显微镜观察并收集图像。提取细胞总蛋白以anti-GST抗体检测目的蛋白表达;比较两者EMSA反应按试剂盒说明书操作进行,以单纯pEGFP-N1质粒与细胞共培养作为对照。
2.4 重组蛋白生物学功能检测
2.4.1 体内表达检测 妊娠哮喘小鼠模型制备参照文献[6]的方法完成并随机分为4组。以尾静脉快速注射方式,按预实验确定的方案,单纯模型组(A组)注入50μl生理盐水;LPS组(B组)注入1 mg·L-1LPS;空载蛋白+LPS组(C组)和纯化重组蛋白+LPS组(D组)分别注入 50μl GST/pCEP4和GST/pCEP4-IL-23后再给予 1 mg·L-1LPS刺激。分别于注射后d 2、d 4、d 7、d 14和d 21行小鼠眼眶静脉丛取血(n=6),血清IL-17A含量测定按ELISA试剂盒说明书进行;7 d时切取小鼠肺组织(n=6)4%多聚甲醛固定后石蜡包埋、切片(4~6μm)、常规HE染色后光镜观察并拍照。
2.4.2 体外表达检测 完全RPMI 1640培养液悬浮Ficoll法分离获得的患者外周血单个核细胞(2×106·L-1),置于预包被 CD3 mAb 24孔板与 20μg·L-1佛波酯、10μg·L-1离子霉素混均 37℃孵育 6 h,实验组加入纯化重组蛋白或空载蛋白4℃条件共培养5 d,对照细胞不做任何处理。按试剂说明每孔先后加入饱和浓度的 FITC-CD4、CD25,PE-IFN-γ、IL-4、IL-17A和PE-Cy5-Foxp3抗体及同型对照,4℃避光反应30 min,PBS/多聚甲醛重悬细胞后进行流式细胞术(FCM)检测,以上步骤重复3次。
2.5 统计学分析 重复实验获得的数据以¯x±s表示并使用SPSS 11.5统计软件进行分析。数据多组间均数比较采用单因素方差分析;两组间样本均数比较采用LSD-t检验。
3 结果
3.1 重组穿梭载体构建与鉴定 p19和p40重组克隆的PCR扩增产物大小为250 bp,均与预期cDNA相符。两者先后插入pMD18-T载体并在分子量约240 bp(p19)和 240 bp(p40)处产生的清晰特异条带与p19和p40亚基DNA长度吻合(Fig 1)。
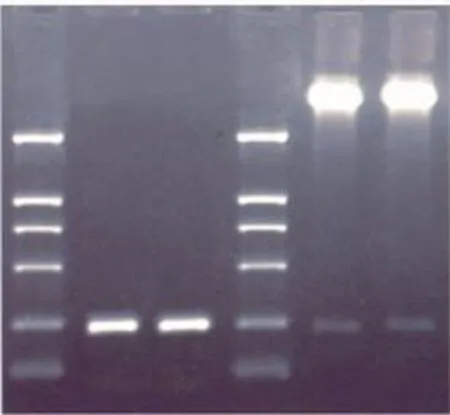
Fig 1 Cloning the target gene and enzyme digestion of the shuttle plasmid
3.2 重组真核双表达载体构建与鉴定 将p19和p40全长基因混合后与pCEP4载体连接,经重叠PCR及酶切鉴定,连接成功的阳性克隆能够释放大小约250 bp(p40+p19)片段(Fig 2),测序结果表明重组真核表达质粒中插入的目的基因序列无突变。
3.3 重组蛋白的表达和纯化 经IPTG诱导表达,含有重组质粒的菌液粗蛋白可溶性较好,GST/pCEP4-IL-23表达的分子质量在约36 000出现的显影条带与预期分子理论值相符(Fig 3A),层析纯度达95%以上;其表达产物与GST发生的特异性结合反应,对应了96 ku处浓重清晰显色条带的分子质量大小(Fig 3B),未诱导对照及GST/pCEP4显色较弱,可用于后续实验。
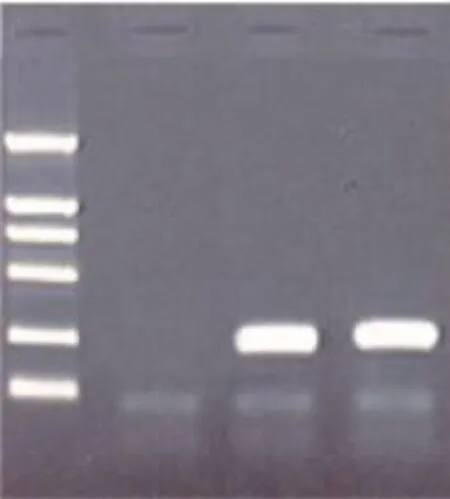
Fig 2 PCR identification of recombinant expression plasmid of cloning

Fig 3 Genetically engineered bacteria and protein chromatography induced effects
3.4 重组蛋白转导活性及DNA竞争结合迁移变化 显微镜下观察到重组蛋白能够将质粒DNA高效携入靶细胞。LPS刺激前EGFP基因以弥散状态分布,刺激后随时间的延长,EGFP表达形成多个荧光聚集且强度增高明显(Fig 4A);Western blot显示分子质量约96 ku的蛋白条带 (Fig 4B),单纯质粒转染细胞LPS刺激前后EGFP及蛋白表达均未见明显变化。
EMSA显示,LPS刺激指示出转染细胞核蛋白的DNA结合活性随蛋白/质粒相对质量比的增加而明显变化。核蛋白提取物中的大分子复合体含量越高,其显影位置凝胶迁移速率越低,区带电泳滞后可将质粒DNA完全阻滞于加样孔中,表明蛋白中和DNA两者的结合是特异的;单纯质粒的DNA结合活性很弱,由于不能与标记探针竞争抑制核蛋白,而呈现清晰的显影条带(Fig 5)。

Fig 4 Expression validation of cell activity after gene transfection

Fig 5 Electrophoretic mobility shift assay
3.5 重组蛋白的体内持续表达对局部炎性改变影响 D组注射4~6 d后血清中IL-17A含量明显高于A、B、C 3组,之后开始回落且逐渐稳定,至21 d时几乎为0(P<0.05,Fig 6);其肺组织炎性细胞浸润、肺泡间隔扩大、粘膜上皮增生等过度炎症损伤最为明显。B、C组血清IL-17A水平及肺组织炎性病理改变明显小于D组但强于A组(Fig 7)。
3.6 重组蛋白体外诱导对T细胞亚群分布及比例变化影响 与未经处理的对照细胞相比较,GST/pCEP4-IL-23组除 IFN-γ、FoxP3细胞数下降,IL-4细胞数以及 Th2/Th1比例升高外,IL-17A细胞和Th17/Treg比例亦明显增多、增高(P<0.05,Fig 8);GST/pCEP4组与对照细胞间的差异无显著性。

Fig 6 Detection of IL-17A level in serum by ELISA analysis(n=6,ng·L-1,¯x±s)
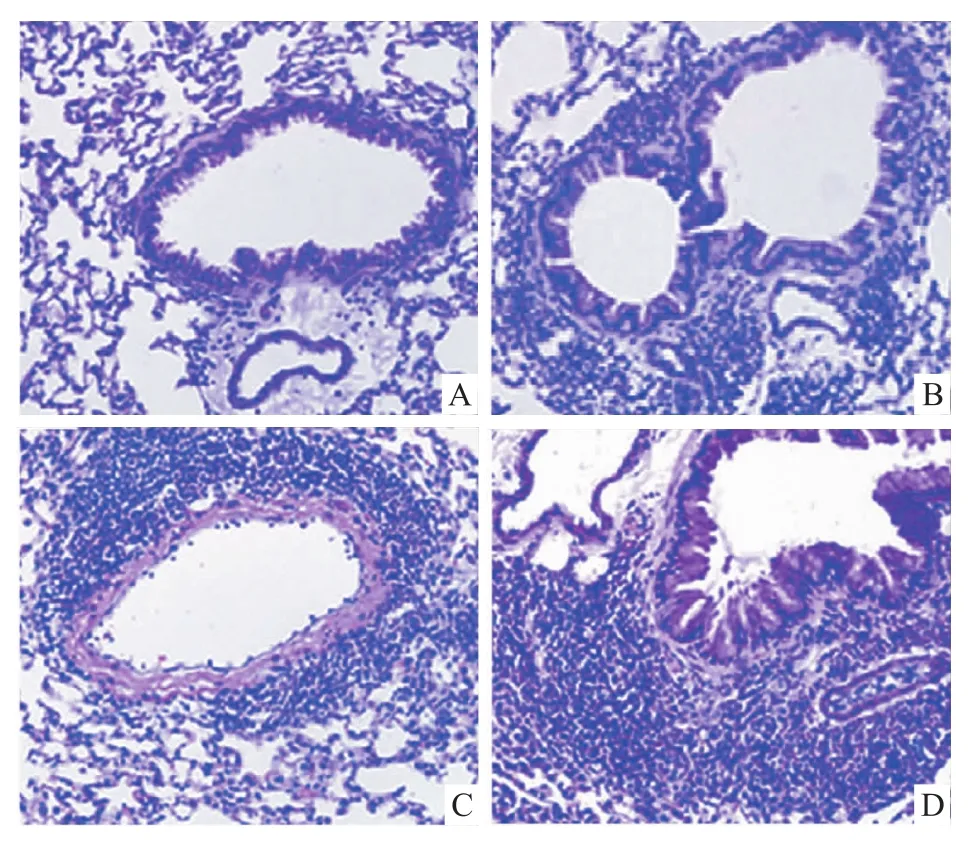
Fig 7 Morphology of lung tissue in different mice groups(HE ×100)
4 讨论
长期以来的观点认为,支气管哮喘发病过程中最重要的免疫异常是Th1/Th2相互拮抗和自身促进的平衡状态被打破。然而,Th1/Th2失衡学说这一模式化理论并不能解释哮喘免疫学基础与临床研究中的全部现象[7]。随着哮喘发病机制研究的深入以及免疫、分子生物学的发展,一种新的假说,即炎症和自身免疫性疾病的发生和发展可能是T辅助细胞反应从Th2、Treg向Th17或者Th1偏移所引起;而阻断关键性细胞因子可使免疫反应从Th17/Th1表型向 Treg和 Th2转换[8]。近来研究[5]证实,Th17的上游调控因子IL-23是具有与IL-12功能相似但更为有效、复杂的效应分子。通过趋化炎症细胞聚集与迁移,导致T细胞反应扩增以及过表达产生的IL-23/IL-17式瀑布效应等,均可造成母体多组织炎症损害、矮小综合症、不育和(或)早产死亡[1]。而这一现象能够解释在发现Th17前的某些自身免疫性疾病模型中出现的理论偏差现象。
IL-23的免疫调控机制十分复杂,其异源二聚体产生以及生物学作用的发挥需要编码p40和p19两种亚基的同时表达[3]。为此,本研究首先采用基因分子拼接技术通过连接肽基因,成功扩增并将p40/p19亚基片段融合拼接在pCEP4载体上;预期编码抗原基因IL-23的重组蛋白表达纯化后,可穿透胞膜将外源质粒携入细胞并转位于胞核,在相关DNA结合位点上形成稳定复合体,通过大分子转运以及调节炎症反应过程来抑制核蛋白的迁移。上述结果确证作者选择载体的多克隆位点不仅便于目的基因的插入,蛋白介导基因跨膜转运直接或间接的功能作用,实现EGFP基因的稳定表达,增加外源物质的DNA结合活性,为探讨重组蛋白免疫功能对IL-23基因表达影响,从细胞转录水平阐明其与妊娠哮喘抗感染免疫的可能机制奠定实验基础,也为生物大分子药物进入组织细胞内发挥治疗作用提供新的思路。
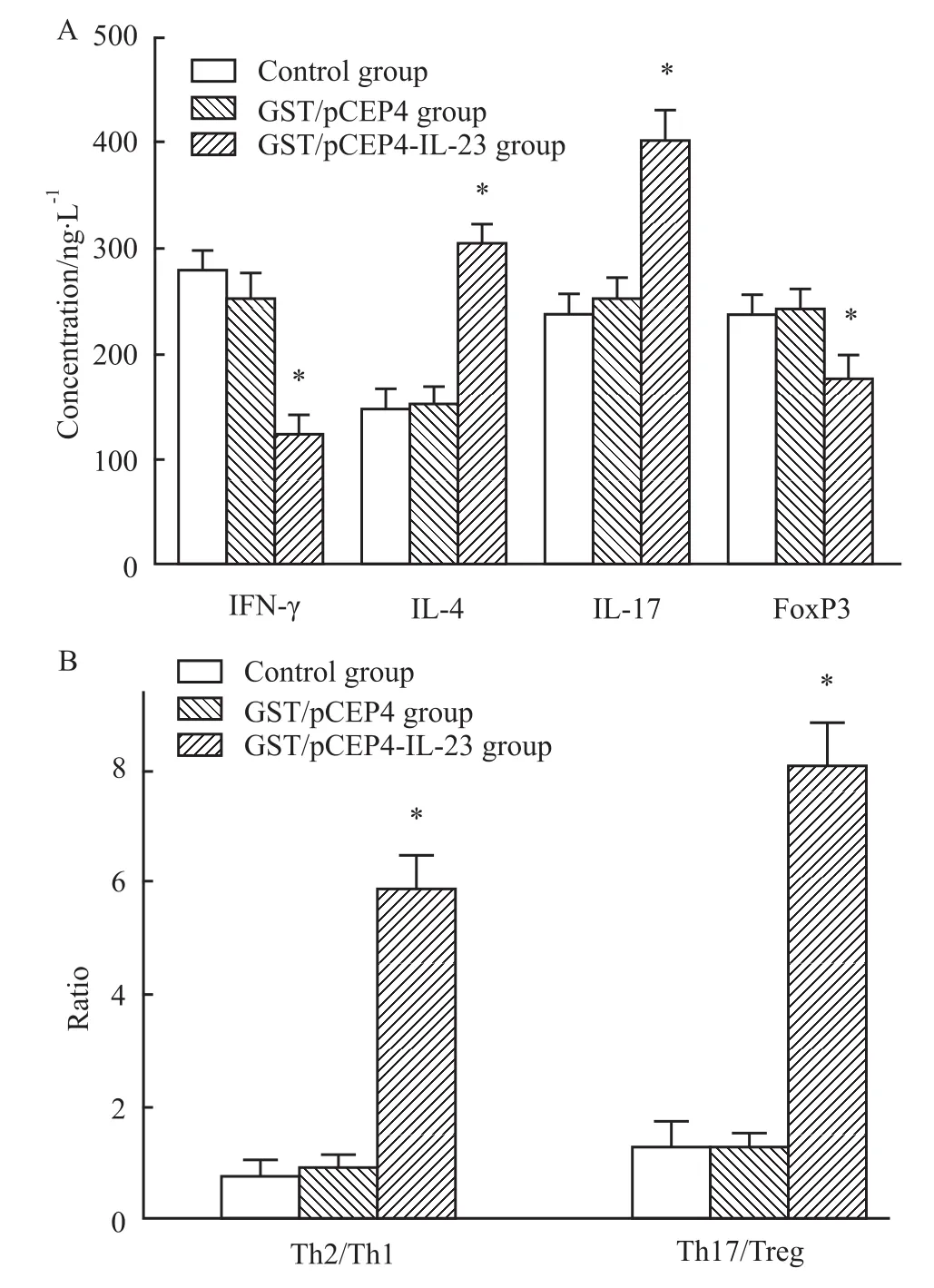
Fig 8 Evaluation of recombinant protein in vitro by FCM(¯x±s,n=6)
在阐明IL-23/IL-17为主轴的免疫调节途径与妊娠期哮喘发病的可能作用及机制方面,实验制作妊娠哮喘小鼠模型,通过体内转导活性检测发现所表达的重组蛋白表现出比对照蛋白更高的活性,经尾静脉注射14 d后小鼠血清中仍可观察到IL-17A的存在,由此产生免疫网络调节系统功能紊乱引发肺组织病理损伤改变增多等,可能是外源性蛋白激活IL-23R信号位点并有效在宿主染色体上整合而不易被降解清除,导致Th17分化增强和IL-17表达增多。除此之外实验进一步观察发现,增加小剂量的LPS,使刺激升高的IL-23R磷酸化水平通过增强p19/p40亚基转录活性,上调了IL-23基因病理性表达[9],而空载蛋白即使增加LPS及单纯LPS诱导,均未能达到相同效果。笔者认为,活化IL-23启动的炎症通路导致免疫平衡向Th17偏移具有一定的相关性。在一定程度上加剧了炎症细胞的浸润与黏附,改变了妊娠哮喘孕妇的局部微环境,是导致母体慢性炎症反复发生的关键所在。笔者的研究表明细胞反应的调节是可诱导的[10];IL-23-IL-23R-Th17-IL-17炎症轴在妊娠哮喘发生、发展过程中存在功能改变,破坏了妊娠哮喘母体自身组织和抗原的免疫耐受,证实其活化和优势应答可致妊娠期哮喘发病这一机制。
本研究体外细胞实验测定IL-23促炎症效应发现,患者外周血中除存在Th2/Th1失平衡外,还同时存在Th17表达增高和Treg数量减少,提示纯化重组蛋白发挥的免疫干扰特性抑制Th1、Treg细胞复制,促进Th2、Th17细胞生成。由此推测,Th1反应向Th2反应转化与Th17/Treg比值升高变化共同参与妊娠期哮喘异常免疫反应调解,使母体和胎儿自体反应性CD4+T细胞抑制功能降低[11]和(或)呈低反应性;激活的相关致炎因子以过度释放方式造成炎症损伤扩大并持久存在,增加妊娠哮喘母体对病毒的易感性[12],促发孕妇罹患哮喘的几率,加重感染后疾病的严重程度;而免疫功能亢进和免疫应答失平衡的双向性变化,最终导致妊娠期哮喘的多种并发症。作者的研究结果及观点进一步证实CD4+T细胞不同亚群在分化过程中和功能上存在相互抑制,支持Th1/Th2/Th17/Treg细胞线性家族间精细而复杂的平衡状态决定疾病形成和进展的发现。
总之,阐明IL-23与其他免疫细胞亚群之间平衡的调节机制以及对机体免疫状态的影响,对于改善甚至逆转妊娠哮喘慢性气道炎症反应的药物开发和探索新的治疗靶点具有重要意义。
参考文献:
[1]Namazy J A,Schatz M.Asthma and rhinitis during pregnancy[J].Mt Sinai J Med,2011,78(5):661-70.
[2]McCallister J W.Asthma in pregnancy:management strategies[J].Curr Opin Pulm Med,2013,19(1):13-7.
[3]Onari Y,Yokoyama A,Haruta Y,et al.IL-12p40 is essential for the down-regulation of airway hyperresponsiveness in a mouse model of bronchial asthma with prolonged antigen exposure[J].Clin Exp Allergy,2009,39(2):290-8.
[4]Guan Q,Ma Y,Aboud L,et al.Targeting IL-23 by employing a p40 peptide-based vaccine ameliorates murine allergic skin and airway inflammation[J].Clin Exp Allergy,2012,42(9):1397-405.
[5]Wakashin H,Hirose K,Iwamoto I,Nakajima H.Role of IL-23-Th17 cell axis in allergic airway inflammation[J].Int Arch Allergy Immunol,2009,149 Suppl 1:108-12.
[6]马佳佳,Nick Lu,陈必良.转录因子RORγt对妊娠期哮喘模型小鼠Th17/Treg平衡的调节作用[J].解放军医学杂志,2012,37(6):561-8.
[6]Ma JJ,Nick Lu,Chen B L.Regulatory effect of transcription factors RORγt on the Th17/Treg balances in pregnant asthma mice[J].Med J Chin PLA,2012,37(6):561-8.
[7]Kim Y M,Kim Y S,Jeon SG,Kim Y K.Immunopathogenesis of allergic asthma:more than the th2 hypothesis[J].Allergy Asthma Immunol Res,2013,5(4):189-96.
[8]Jang SO,Kim H J,Kim Y J,et al.Asthma prevention by lactobacillus rhamnosus in a mouse model is associated with CD4(+)CD25(+)Foxp3(+)T cells[J].Allergy Asthma Immunol Res,2012,4(3):150-6.
[9]Cho Y C,Lee SH,Lee M,et al.Enhanced IL-12p40 production in LPS-stimulated macrophages by inhibiting JNK activation by artemisinin[J].Arch Pharm Res,2012,35(11):1961-8.
[10]Rafa H,Saoula H,Belkhelfa M,et al.IL-23/IL-17A Axis Correlates with the Nitric Oxide Pathway in Inflammatory Bowel Disease:Immunomodulatory Effect of Retinoic Acid[J].J Interferon Cytokine Res,2013,33(7):355-68.
[11]Toldi G,Molvarec A,Stenczer B,et al.Peripheral T(h)1/T(h)2/T(h)17/regulatory T-cell balance in asthmatic pregnancy[J].Int Immunol,2011,23(11):669-77.
[12]Eszes N,Bohács A,Cseh A,et al.Relation of circulating T cell profiles to airway inflammation and asthma control in asthmatic pregnancy[J].Acta Physiol Hung,2012,99(3):302-10.