1例有汗性外胚层发育不良患者的基因突变分析
2014-03-22蒋树娟黄丹何蓉
蒋树娟,黄丹,何蓉
(中国医科大学附属盛京医院临床遗传科,沈阳110004)
1例有汗性外胚层发育不良患者的基因突变分析
蒋树娟,黄丹,何蓉
(中国医科大学附属盛京医院临床遗传科,沈阳110004)
Mutation Analysisin Patientwith Hidrotic EctodermalDysplasia:ACase Report
利用DNA测序技术对1例中国汉族有汗性外胚层发育不良患者进行GJB6基因突变检测,并利用生物信息学软件对检测到的突变进行结构分析,明确有汗性外胚层发育不良的致病原因。结果发现,GJB6基因c.31G>A(p.G11R)突变是该有汗型外胚层发育不良患者的致病原因之一。
有汗型外胚层发育不良;基因突变;GJB6;DNA测序;结构分析
有汗型外胚层发育不良(hidrotic ectodermal dysplasia,HED),又称Clouston综合征,人类孟德尔遗传在线编号为129500,是一种罕见的常染色体显性遗传性皮肤疾病,发病率约为1/10万。临床主要特点为甲发育不良、掌跖角化、毛发稀疏或完全缺如,患者汗腺和皮脂腺功能正常,部分患者可有白内障、斜视、骨骼改变、听力减退、智力障碍及指(趾)畸形,临床表现具有多样性[1]。本病是由编码连接蛋白30(connexin 30,Cx30)的GJB6基因突变所致[2]。我们利用DNA测序技术对来门诊进行遗传咨询的1例中国汉族有汗型外胚层发育不良患者及其父亲进行了致病基因突变分析,报道如下:
1 材料与方法
1.1 临床资料
患者男,28岁,自出生就发现头发、指甲、趾甲发育不良。查体:无头发,头皮仅见少量毫毛;眉毛、睫毛、阴毛、腋毛等体毛缺如,甲板短小,指甲、趾甲发育不良、短缩、变薄,有深的纵纹(图1A和1B),牙齿发育正常,汗腺分泌正常,视力、听力无异常,智力发育正常,腋窝皮肤病理示表皮基本正常,真皮散在毛囊、皮脂腺结构,汗腺丰富(图2A)。患者父亲,50岁,临床表现比患者轻,头发、眉毛及睫毛正常,阴毛、腋毛稀少,指甲、趾甲增厚呈灰黄色(图1C和1D),右上肢及前胸可见散在红斑,牙齿发育正常,汗腺分泌正常,视力及听力正常,智力发育正常,腋窝皮肤病理显示表皮基本正常,真皮内未见毛囊、皮脂腺结构,真皮下部汗腺丰富(图2B)。
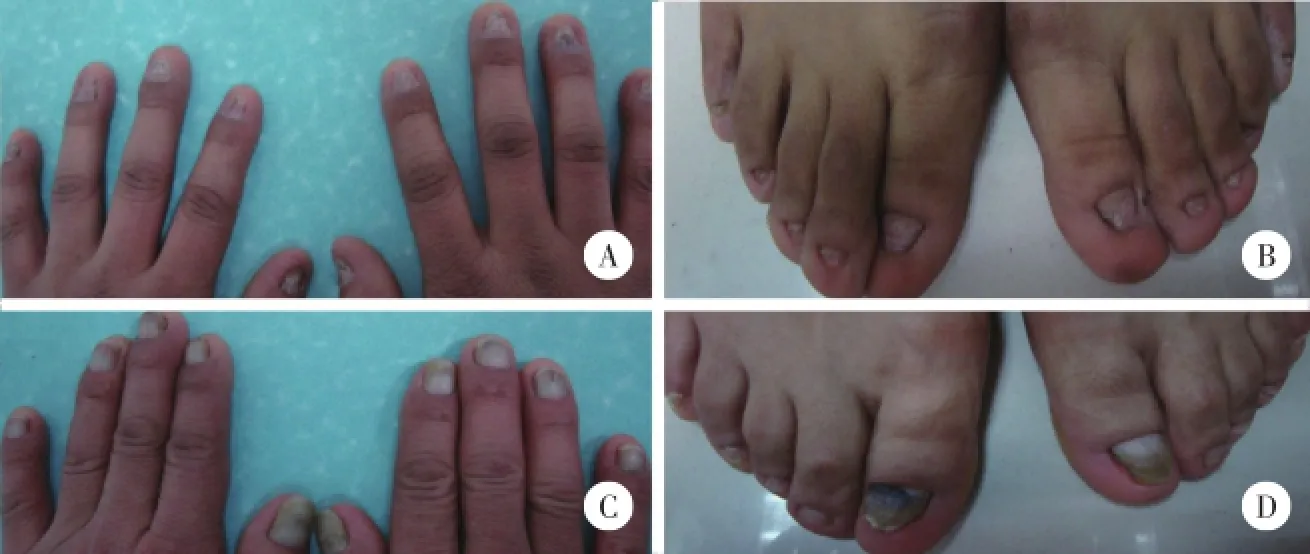
图1 患者及其父亲甲表现
1.2 方法
1.2.1 基因组DNA提取:经患者及家属知情同意,采集患者及其父亲的外周血,并获得了他们签署的知情同意书。采用上海华舜生物技术有限公司的外周血DNA抽提试剂盒提取基因组DNA。另外以同样的方法提取160例无亲缘关系的健康对照个体基因组DNA作为对照。
1.2.2 突变检测:GJB6含有一个外显子,我们参照文献[3]合成引物,对GJB6整个编码区进行扩增,产物长度990 bp。PCR反应体系为25 μL,含有基因组DNA 100 ng,引物0.5 μmol/L,1×PCR缓冲液,1 μmol/L dNTPs,1.5 μmol/L MgCl2,1U Taq DNA聚合酶(大连TaKaRa公司)。PCR反应条件:95℃变性5 min,95℃45 s,62℃45 s,72℃1 min进行35个循环,72℃延伸10 min。4 μL PCR产物用1%的琼脂糖凝胶电泳检测。PCR扩增的目的片段纯化回收后用ABI3730测序仪(美国Applied Biosystems by Life Technologies)进行双向测序,测序引物同前。测序结果用DNAStar 5.0和BioEdit软件与人类基因组GJB6序列(NG_008323.1)进行比对。
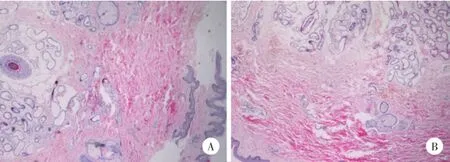
图2 患者及其父亲右腋下皮肤病理
1.2.3 Cx30结构分析:从NCBI数据库(http://www.ncbi.nlm. nih.gov/)下载不同种属的Cx30序列,利用BioEdit软件对下载的氨基酸序列进行比对,比较Cx30突变位点区域的种属差异。瑞士生物信息学研究所创建的蛋白质分析专家系统包含了蛋白质三级结构预测的SWISS-MODEL(http://swissmodel.expasy.org/)数据库,利用该网站的在线预测功能,以Cx26(PDB编码:2ZW3)为模板对野生型和突变后的Cx30进行蛋白质三级结构预测,并进一步利用生物信息学软件Py-MOL(http://www.pymol.org)对突变前后的蛋白质三级结构进行比较[4]。
2 结果
2.1 GJB6基因突变分析
测序结果表明,患者及其父亲携带GJB6基因杂合错义突变c.31G>A(p.G11R),该突变导致Cx30第11位氨基酸由甘氨酸(Gly)变成精氨酸(Arg),160例无亲缘关系的正常人均未见此改变(图3)。
2.2 Cx30结构分析
通过氨基酸序列比对可以看出Cx30的第11个氨基酸位置(p.G11)在各个种属(如人、猩猩、狗、鼠、牛、鸡)中高度保守(图4A)。三级结构分析显示,突变之前甘氨酸(Gly11)可以与Cx30氨基端的组氨酸(His7)和缬氨酸(Val13)形成氢键,从而维持Cx30正常的三级结构;突变成精氨酸(Arg11)之后,增加的侧链还可以与氨基端的苏氨酸(Thr8)和第一跨膜区的苏氨酸(Thr26)形成氢键(图4B、C),这可能会影响Cx30的正常折叠,从而影响其正常的空间构象。另外,突变为精氨酸之后Cx30氨基末端的局部区域还增加了正电荷,这可能会对Cx30正常结构的形成产生影响,或对形成的连接蛋白通道的通透性产生影响。
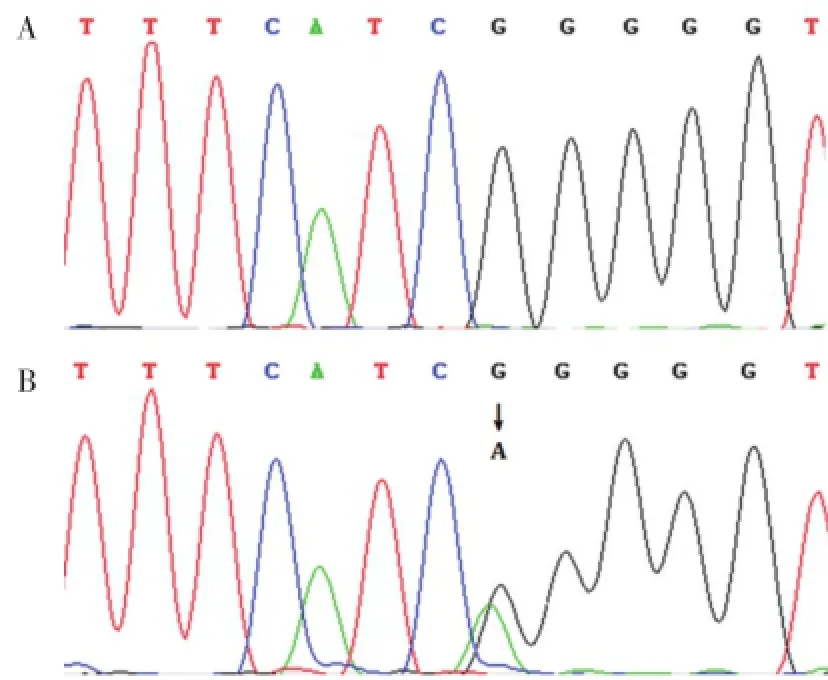
图3 GJB6基因测序图
3 讨论
有汗型外胚层发育不良是一种罕见的常染色体显性遗传性疾病,多见于法籍加拿大人。1996年Kibar等[5]利用基因连锁分析确定了HED2的染色体定位,2000年Lamartine等[6]进一步证实GJB6基因突变是该综合征的致病原因。迄今为止,在HED患者中已发现该基因的4种突变:(1)大多数HED患者发病的原因是c.31G>A(p.G11R)突变,该突变导致Cx30的氨基末端引入了一个正电荷的精氨酸。Lamartine等[6]研究了来自不同地理区域(苏格兰、非洲、西班牙、法裔加拿大人、法国、印度、马来群岛及威尔士)的12个患病家系,发现除了印度、马来群岛和威尔士的患者携带c.263C>T突变外,几乎所有的患者为c.31G>A突变,这一突变还在黎巴嫩和德国患者中被发现[7],中国也相继在5个家系中发现这个突变[8~12],本研究也在先证者和其父亲中检测到c.31G>A(p.G11R)突变的存在,进一步支持此突变是导致HED发病的主要原因。(2)错义突变c.263C>T(p.A88V)[2],该突变在Cx30的第2个跨膜区引入了1个高疏水性的氨基酸残基。李汶等[13]也在一中国家系中发现此突变,并在基因诊断基础上对该家系中一孕妇孕5个多月的胎儿进行了产前诊断。(3)第3种突变c.110T>A(p.V37E),该突变是Smith等[14]在研究一个散发的来自苏格兰的病例时发现的,它直接导致了Cx30第1个跨膜区域的氨基酸的改变,引入了一个带负电荷的谷氨酸。(4)第4种突变c.148G>A(p.D50N),该突变发生在Cx30的第一个胞外区,是Baris等[15]在患有HED的北欧犹太人中发现的,后两种突变尚未在中国患者中发现。
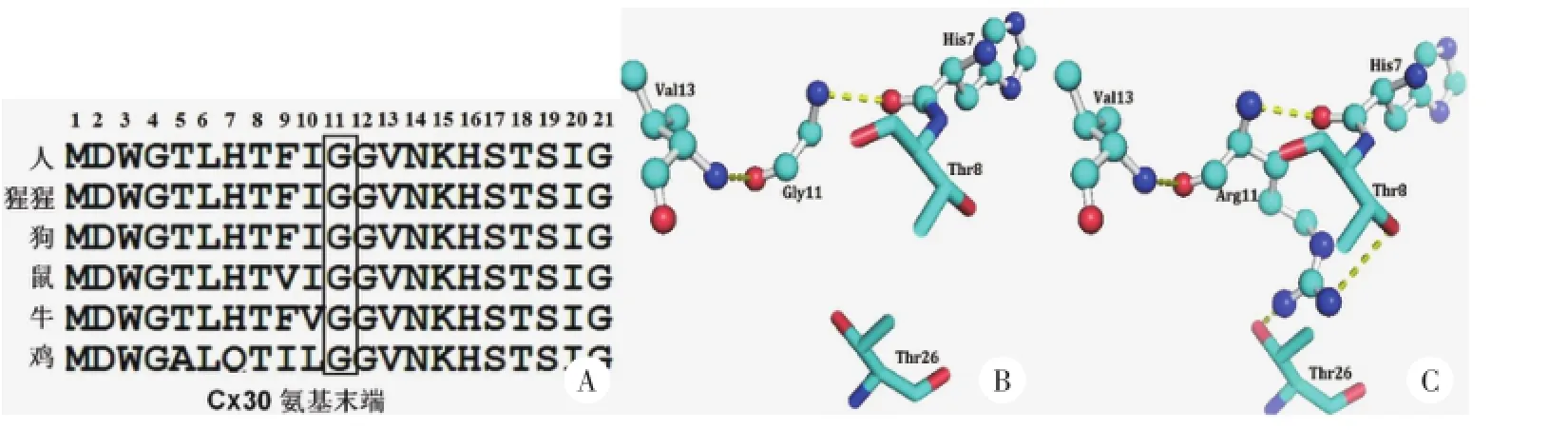
图4 Cx30氨基末端氨基酸序列比对及三级结构分析
GJB6基因编码Cx30蛋白,属于缝隙连接蛋白基因家族,与相邻细胞膜上的半通道或称连接子组成一个完整的缝隙连接通道,使相邻细胞间离子、代谢物质和小的信使分子直接转运,对细胞增殖、分化及机体的生长发育具有意义[16]。据报道,Cx30在特定的组织优先表达,在小鼠中,Cx30在成鼠的大脑和皮肤中高度表达[17]。Fujimoto等[7]利用免疫染色的方法证明Cx30在毛囊、表皮和指甲中高度表达,并证明其在表皮细胞的分化、毛囊和指甲的生长过程中起到了重要的作用,由此推测突变的Cx30可能影响这些组织的增殖和分化,从而导致HED的发生。连接蛋白结构上具有相似性,每种连接蛋白都具有4个跨膜区,2个胞外区,3个胞内区,人类Cx30编码261个氨基酸,与Cx26具有77%的同源性[18]。本研究在患者中检测到的突变发生在Cx30的氨基末端,为了进一步明确此突变的致病机制,我们对不同种属的氨基酸序列进行了比对,并对突变前后Cx30的三级结构进行了比较,通过氨基酸序列比对可以看出Cx30的第11个氨基酸位置(p. G11)在人、猩猩、狗、鼠、牛、鸡各个种属中高度保守。甘氨酸是只含有2个碳原子的脂肪族氨基酸,当甘氨酸被含有6个碳原子带有正电荷的精氨酸取代后,不仅增加了侧链的长度,还引入了能提供正电荷的氨基,三级结构分析显示,突变之前甘氨酸(Gly11)可以与Cx30氨基端的组氨酸(His7)和缬氨酸(Val13)形成氢键,从而维持Cx30正常的三级结构;突变成精氨酸(Arg11)之后,增加的侧链还可以与氨基端的苏氨酸(Thr8)和第一跨膜区的苏氨酸(Thr26)形成氢键,这可能会影响Cx30的正常折叠,从而影响其正常的空间构象。另外,突变为精氨酸之后Cx30氨基末端的局部区域还增加了正电荷,这可能会对Cx30正常结构的形成产生影响,或对形成的连接蛋白通道的通透性产生影响。
有汗型外胚层发育不良的临床表现具有多样性,本研究中患者主要表现为甲和毛发的异常,不伴有掌跖角化。患者和其父亲的表现不尽相同,患者指、趾甲表现为甲板短小,甲发育不良、短缩、变薄,有深的纵纹,而其父亲则表现为指、趾甲增厚、呈灰黄色;患者毛发损害较重,头发及各种体毛均缺如,而其父亲毛发损害较轻,除阴毛、腋毛稀少外头发、眉毛及睫毛等均正常。同为c.31G>A(p.G11R)突变导致的HED,文献[8,10,12]报道的家系中患者有典型的三大症状(甲发育不良、掌跖角化、毛发稀疏或完全缺如);陈楠[9]、刘宁等[11]报道的家系与本研究患者的表现相同,除了甲和毛发异常外,不伴有掌跖角化;除此之外此突变还可伴有杵状指和或智力低下等[8,9]。同一突变在相同种族人群中导致的表型差异可能与基因突变的外显度不同相关,或与某些未知的调节机制相关,也可能与环境因素相关。
HED临床表现的多样性和个体差异对临床诊断造成困难,利用GJB6基因突变检测方法对HED患者进行检测能够明确病因,从而更好地为患者提供遗传咨询。对Cx30蛋白结构的分析能使我们更好地理解HED的致病机制。
[1]Clouston HR.Major forms of hereditary ectodermal dysplasia[J]. Can Med Assoc J,1939,40(1):1-7.
[2]Lamartine J,Munhoz Essenfelder G,Kibar Z,et al.Mutations in GJB6 cause hidrotic ectodermal dysplasia[J].Nat Genet,2000,26(2):142-144.
[3]Chen P,Chen H,Fu S,et al.Prevalence of GJB6 mutations in Chinese patients with non-syndromic hearing loss[J].Int J Pediatr Otorhinolaryngol,2012,76(2):265-267.
[4]Maeda S,Nakagawa S,Suga M,et al.Structure of the connexin 26 gap junction channel at 3.5A resolution[J],Nature,2009,458(7238):597-602.
[5]Kibar Z,Der Kaloustian VM,Brais B,et al.The gene responsible for Clouston hidrotic ectodermal dysplasia maps to the pericentromeric region of chromosome 13q[J].Hum Mol Genet,1996,5(4):543-547.
[6]Lamartine J,Laoudj D,Blanchet-Bardon C,et al.Refind localization of the gene for Clouston syndrome(hidrotic ectoderm al dysplasia)in a large French family[J].Br J Dermatol,2000,142(2):248-252.
[7]Fujimoto A1,Kurban M,Nakamura M,et al.GJB6,of which mutations underlie Clouston syndrome,is a potential direct target gene of p63[J].J Dermatol Sci,2013,69(2):159-166.
[8]陈建军,杨森,崔勇,等.有汗性外胚层发育不良一家系的连接蛋白30基因突变检测[J].中华皮肤科杂志,2003,36(11):613-615.
[9]陈楠,张锐利,王震英,等.基因诊断有汗性外胚层发育不良一家系[J].中华皮肤科杂志,2009,42(11):745-747.
[10]王珍,曹丽华,李铁男,等.有汗性外胚层发育不良一家系基因突变[J].国际皮肤性病学杂志,2012,38(5):283-285.
[11]刘宁,史惠蓉,吴庆华,等.有汗型外胚层发育不良家系基因突变分析及孕早期产前诊断[J].中华医学遗传学杂志,2013,30(4):407-410.
[12]TANIA Mousumi,熊志敏,鹿丽娜,等.一个中国先天性有汗性外胚层发育不良家系的GJB6基因筛查[J].中南大学学报(医学版),2013,38(8):761-765.
[13]李汶,高伯笛,李麓芸,等.有汗型外胚层发育不良一家系的Cx30基因突变检测及产前诊断[J].中华医学遗传学杂志,2006,23(6):618-621.
[14]Smith FJ,Morley SM,McLean WH,et al.A novel Connexin 30 mutation in Clouston syndrome[J].J Invest Dermatol,2002,118(3):530-532.
[15]Baris HN,Zlotogorski A,Peretz-Amit G,et al.A novel GJB6 missense mutation in hidrotic ectodermal dysplasia 2(Clouston syndrome)broadens its genotypic basis[J].Br J Dermatol,2008,159(6):1373-1376.
[16]Langlois S,Maher AC,Manias JL,et al.Connexin levels regulate keratinocyte differentiation in the epidermis[J].J Biol Chem,2007,282(41):30171-30180.
[17]Dahl E,Manthey D,Chen Y,et al.Molecular cloning and functional expression of mouse connexin-30,a gap junction gene highly expressed in adult brain and skin[J].J Biol Chem,1996,271(30):17903-17910.
[18]Zonta F,Polles G,Zanotti G,et al.Permeation pathway of homomeric connexin 26 and connexin 30 channels investigated by molecular dynamics[J].J Biomol Struct Dyn,2012,29(5):985-998.
(编辑 陈姜)
R394.3
A
0258-4646(2014)10-0941-04
蒋树娟(1985-),女,博士研究生.
何蓉,E-mail:her@sj-hospital.org
2013-06-01
网络出版时间:
