热休克蛋白70对骨骼肌细胞系(C2C12)葡萄糖代谢的影响
2014-03-16杨伟伟
王 磊,王 尊,杨伟伟
(南京中医药大学第二临床医学院康复医学系,江苏南京210001)
热休克蛋白70(heat shock protein,HSP70)是最重要的应激反应蛋白,其最基本功能是感受刺激和分子伴侣功能[1]。骨骼肌细胞正常状态下HSP70 表达水平很低,当受到各种内外环境刺激时,HSP70 的表达快速的升高,起多种保护作用维持细胞内环境的稳定。研究表明HSP70 的表达水平与骨骼肌细胞葡萄糖代谢的紊乱程度相关,包括ATP 的耗竭,储存的糖原的下降以及乳酸的积聚等[2],提示HSP70 对骨骼肌细胞葡萄糖代谢产生影响,但具体作用及相关机制尚不清楚。本研究通过构建高表达HSP70 的C2C12 细胞系,研究HSP70 对骨骼肌细胞葡萄糖代谢影响及相关机制,为最终阐明HSP70 在骨骼肌细胞葡萄糖代谢中的作用提供理论依据。
1 材料与方法
1.1 材料
真核表达载体pTRE2hyg(Novagen 公司);含有人HSP70cDNA(ATCC57494)的质粒pAT153(德国ULM 大学Yuefei Liu 教授惠赠);RNeasy Mini Kit及real-time PCR 试剂盒(QIAGEN 公司);HSP70 抗体和ATP biolumin-escence assay 试剂(Sigma 公司);C2C12 细胞系,DMEM 培养基,G418 和小牛血清(Gibico 公司)。葡萄糖测定试剂盒(北京普利莱基因技术有限公司);3H-2 -脱氧葡萄糖(放射性比活度5.4 Bq/mmol)(中国同位素总公司);HK、PK、PFK 和LDH 的活性测定试剂盒及Glut4 抗体(Sigma公司)。
1.2 方法
1.2.1 细胞分组:将C2C12 细胞分为HSP70 转染组与正常对照组,分别培养分化后进行相关指标检测。
1.2.2 构建重组pTRE2hyg-HSP70 质粒:1)用Hind Ⅲ限制性内切酶切开含有人HSP70 cDNA 的质粒pAT153,用NheⅠ限制内切酶切开载体pTRE2hyg,补平切出的黏性末端;2)经过电泳,纯化得到HSP70 cDNA 和pTRE2hyg 分子;3)将纯化的HSP70和pTRE2hyg 分子按1∶3 的比例用T4 连接酶进行连接,后转化到DH5α 菌株中,以含氨苄青霉素筛选抗性克隆。4)将挑选出的克隆进行BamHⅠ酶切鉴定后,大量制备重组质粒pTRE2hyg-HSP70。
1.2.3 C2C12 细胞培养:骨骼肌细胞系C2C12 细胞培养于5% CO2和37 ℃培养箱中,DMEM 培养基中含体积分数10%小牛血清1 ×105U/L 青霉素100 mg/L 链霉素。
1.2.4 重组pTRE2hyg-HSP70 质粒稳定转染C2C12细胞系:1)采用脂质体转染质粒的方法将pTRE2 hyg-HSP70 质粒转染C2C12 细胞:取5 μL Lipofectin®Reagent 与100 μLOpti ® -MEM 混合30 min,取0.7 μg重组pTRE2hyg-HSP70 质粒溶于100 μL 的无血清培养液中并加入PLUS 混合15 min。并转染C2C12 细胞3 h 后,置换为含10%血清的DMEM 培养液培养24 h。按1∶10 稀释细胞转至10 cm 培养皿,次日加入终浓度为500 mg/L 的G418 进行筛选。3)检测转染后的C2C12 细胞表达HSP70:选取抗性细胞克隆,采用1 μg/mL 的强力霉素诱导后,Western blot 方法检测HSP70 的表达情况。4)取细胞以5 × 105/mL 接种于将96 孔板中,换为含50 mL/L马血清的高糖DMEM 培养液诱导C2C12细胞分化成为肌小管(myotube),在倒置显微镜下观察诱导分化情况,当在镜下见80%~90%的细胞汇合时取用。
1.2.5 稳定转染C2C12 细胞葡萄糖消耗和乳酸产生的检测:将96 孔板中诱导分化的C2C12 骨骼肌细胞,以含1% BSA 低糖DMEM 培养基孵化8 h,然后以葡萄糖氧化酶法检测每孔培养基中的葡萄糖含量。用不含细胞的培养孔内培养基中的葡萄糖含量的均值减去各组培养孔培养基中剩余的葡萄糖含量,即为每孔细胞的葡萄糖消耗量。具体操作步骤见试剂盒说明。同时用YSI 2300 STAT PLUS 乳酸生化分析仪检测培养基中乳酸的浓度,并通过计算得出每8 h 葡萄糖的消耗及乳酸的产生量。
1.2.6 稳定转染C2C12 细胞葡萄糖摄取的测定:接种5 ×105/mL 细胞于孔板中,分化培养至80%~90%汇合,置换无血清5.5 mmol/L 葡萄糖的MEM培养液,让细胞处在无血清和低糖状态。每孔加入37 kBq3H-2-脱氧葡萄糖,在室温下放置10 min。每孔立即加入10 μL 浓度为0.48 g/L 细胞松弛素B(终浓度为10 μmol/L),终止摄取。用PBS 冲洗2次,每孔加入0.5 g/L NaOH 200 μL,室温静置2 h,裂解细胞,吸附后液烁计数检测。
1.2.7 稳定转染C2C12 细胞内ATP 水平的检测:收集孔板中分化C2C12 细胞,用冰PBS(pH 7.4)洗净后,加入Tris EDTA 缓冲液(100 mmol/L Tris-HCl和4 mmol/L EDTA,pH 7.55)在100 ℃孵育3 min后,10 000 ×g 离心2 min 取上清。采用ATP 生物发光检测试剂检测ATP 的含量。ATP 的水平将从ATP(1.56~800 nmol/L)标准的双对数曲线中计算出来,以nmol/mg pro.来表示。
1.2.8 稳定转染C2C12 细胞糖酵解关键酶活性的检测:收集孔板中分化C2C12 细胞,用PBS 重新悬浮,反复冻融3 次,4 ℃,2 000 ×g 离心20 min,取上清液检测PFK、LDH、HK 和PK 的活性,具体步骤见文献[3]。以上各种酶的活性通过耦合NADP 的减少或NADH 的氧化进行,并通过分光光度计检测在334 nm 处的吸光度值,酶活性的结果采用nmol/min·mg pro.来表示。
1.2.9 稳定转染C2C12 细胞膜Glut4 的表达检测:1)收集孔板中分化C2C12 细胞,用PBS 重新悬浮,反复冻融3 次,4 ℃,9 000 ×g 高速离心10 min,保留上清。将上清以4 ℃,190 000 × g 超高速离心1 h,取沉淀铺于25%、30%、35% 蔗糖梯度上,4 ℃,150 000 ×g 超离心16 h。于25%蔗糖层得到骨骼肌细胞膜悬浊液,35%层上得到骨骼肌细胞内质膜悬浊液。膜蛋白浓度用蛋白定量试剂盒测定。2)蛋白定量后,取25 μL 样品10%聚丙烯酰胺凝胶上进行SDS 凝胶电泳,随后转移至PVDF膜上,封闭,然后加入兔抗鼠Glut4 单克隆抗体过夜(4 ℃),洗涤后与HRP 标记的抗兔IgG 抗体室温孵育1 h(37 ℃),凝胶成像系统检测Western blot 蛋白条带,经β-actin 校正,gene 软件分析蛋白表达水平。
1.3 统计学分析
2 结果
2.1 质粒酶切鉴定及PCR 质粒鉴定
成功构建pTRE2hyg-HSP70 质粒。P1:pAT153质粒被HindIII 和BamHI 酶切(2.3 kb 和3.4 kb);P2:Hind Ⅲ酶切pAT153 (5.7 kb);P3:修饰后pTRE2hyg 载体(2.3 kb);P4:修饰后目标基因HSP70(5.3 kb);P5:重组pTRE2hyg-HSP70 质粒(7.6 kb);M:1 kb DNA marker(图1)。
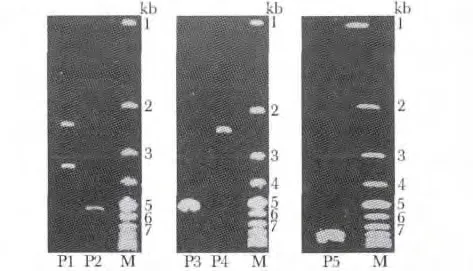
图1 pTRE2hyg-HSP70 质粒重组鉴定Fig 1 pTRE2hyg-HSP70 recombinant plasmid colony by PCR identified
2.2 C2C12 细胞培养及pTRE2hyg-HSP70 质粒转染C2C12 细胞
C2C12 细胞转染后筛选发现(No5 C 组和No12 D 组)克隆经过诱导后能够稳定高效的表达HSP70,说明质粒转染C2C12 成功(图2)。
2.3 高表达HSP70 的C2C12 细胞葡萄糖消耗、乳酸产生及葡萄糖摄取的检测
与对照组相比,在细胞分化后的3 和7 d,过表达HSP70 的C2C12 细胞每8 h 葡萄糖的消耗和乳酸的产生量都明显增加(P<0.01),胰岛素刺激的葡萄糖摄取量也明显增加(P<0.05)(表1)。
2.4 高表达HSP70 对C2C12 细胞内ATP 水平的影响
与对照组相比,在细胞分化后的3 和7 d,过表达HSP70 的C2C12 细胞ATP 水平明显升高(P<0.01)(表1)。

图2 pTRE2hyg-HSP70 转染C2C12 细胞后HSP70 的表达Fig 2 The expression of HSP70 in C2C12 cells after pTRE2hyg-HSP70 transfection
2.5 高表达HSP70 的C2C12 细胞糖酵解关键酶的检测
与对照组相比,在细胞分化后的3 和7 d,过表达HSP70 的C2C12 细胞PFK,HK,PK(P<0.01)和LDH(P<0.05)的活性均显著增加(表2)。
2.6 高表达HSP70 的C2C12 细胞膜Glut4 的表达检测
对照组相比,在细胞分化后的3 和7 d,过表达HSP70 的C2C12 细胞膜Glut4 蛋白的表达明显增高(P<0.05)(图3)。
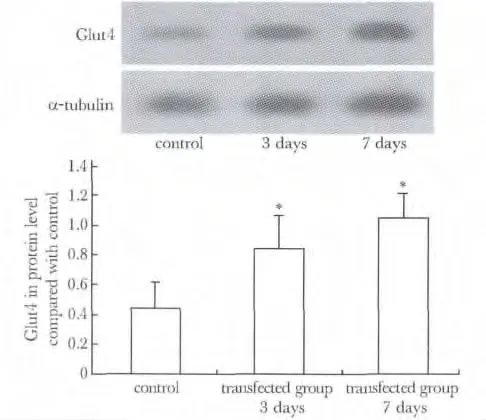
图3 高表达HSP70 的C2C12 细胞膜Glut4 的表达Fig 3 The expression of plasma membrane Glut4 in protein level in up-regulated HSP70 C2C12 cells
3 讨论
骨骼肌细胞葡萄糖代谢功能对其能量平衡十分重要,也同许多重要的疾病密切相关。HSP70 是重要的应激反应蛋白,具有保护细胞内环境稳定的功能。研究表明,许多刺激可以同时引起肌细胞糖代谢的紊乱及HSP70 表达的变化,且HSP70 功能的实现有赖于能量(ATP)的供给,提示HSP70 可能直接或间接对肌细胞的葡萄糖代谢产生影响[4],但目前尚无直接证据。
表1 稳定转染HSP70 后C2C12 细胞葡萄糖消耗、乳酸产生、葡萄糖摄取及细胞内ATP 水平变化Table 1 Changes of glucose consumption,glucose uptake,lactate production and ATP level in tansfected C2C12 cells (±s,n=3)

表1 稳定转染HSP70 后C2C12 细胞葡萄糖消耗、乳酸产生、葡萄糖摄取及细胞内ATP 水平变化Table 1 Changes of glucose consumption,glucose uptake,lactate production and ATP level in tansfected C2C12 cells (±s,n=3)
*P<0.05,**P<0.01 compared with control.
groupglucose consumption(mmol/L/107 cells/8 hours)lactate production(mmol/L/107 cells/8 hours)glucose uptake(nmol/L/107cells/h)ATP level(nmol/mg protein)normal control3.63 ±1.025.33 ±1.533.85 ±0.395.62 ±1.32 transfected group 3 days7.32 ±2.35**14.62 ±4.36**5.68 ±1.48*13.43 ±3.89**transfected group 7 days7.51 ±2.16**13.35 ±4.21**5.87 ±1.34*14.82 ±4.08**
表2 稳定转染HSP70 后C2C12 细胞葡萄糖酵解关键酶活性的变化Table 2 Changes of activities of enzymes involved in glycolysis in tansfected C2C12 cells(nmol/min·mg protein,±s,n=3)

表2 稳定转染HSP70 后C2C12 细胞葡萄糖酵解关键酶活性的变化Table 2 Changes of activities of enzymes involved in glycolysis in tansfected C2C12 cells(nmol/min·mg protein,±s,n=3)
*P<0.05,**P<0.01 compared with control.
groupPKFLDHHKPK normal control18.63 ±5.243 900 ±4682.61 ±0.6516.43 ±3.78 transfected group 3 days38.57 ±13.45**5 879 ±821*6.93 ±1.69**31.25 ±9.63**transfected group 7 days41.65 ±15.26**6 123 ±854*6.85 ±1.54**34.31 ±9.67**
本研究结果显示高表达HSP70 的骨骼肌细胞能够增加葡萄糖的摄取和利用,增加细胞内ATP 的含量,提高与葡萄糖酵解相关的酶的活性,显示了高表达的HSP70 能够加速骨骼肌细胞的糖酵解,明确了HSP70 对葡萄糖代谢的直接作用。相关研究也证实,如运动,缺血等刺激都可以引起骨骼肌细胞HSP70 的表达,并且HSP70 表达的水平同刺激的类型、强度和时间密切相关[5]。如骨骼肌细胞糖原和ATP 的耗竭,乳酸的堆积,氧自由基的产生等都可以促进细胞HSP70 的表达[6-7]。结合本实验结果,HSP70 可以通过加速葡萄糖的摄取和糖酵解,快速产生ATP 来满足细胞对能量的需求。之前的研究中也发现,低频率(10 Hz)长时间(90 min)的刺激可引起C2C12 细胞高表达HSP70,同时伴有培养液中葡萄糖含量的减少和乳酸生成的增多,及无氧代谢酶的活性增加等[8-9]。以上都进一步证明HSP70的表达可以提高糖酵解水平,维持骨骼肌细胞能量代谢的平衡。
葡萄糖进入骨骼肌细胞主要依赖葡萄糖载体的转运,Glut4 是存在肌细胞内和膜表面的主要葡萄糖转运体。研究证实,运动可以提高肌细胞Glut4蛋白的表达及转位[10-11],从而提高细胞对葡萄糖的摄取和利用。本实验的结果也显示,高表达HSP70能够明显提高骨骼肌细胞Glut4 蛋白的表达,提示HSP70 可以通过提高骨骼肌C2C12 细胞膜Glut4 的表达增加细胞对葡萄糖摄取。
综上所述,过表达HSP70 的骨骼肌细胞可以通过提高肌细胞膜Glut4 的表达增加葡萄糖的摄取,并提高糖酵解相关酶的活性加速肌细胞糖酵解和提高细胞内ATP 水平,维护细胞能量代谢的平衡。显示HSP70 对细胞葡萄糖代谢的直接作用,但HSP70 调节细胞葡萄糖代谢的具体机制,还有待于进一步研究。
[1]Liu Y,Steinacker JM.Changes in skeletal muscle heat shock proteins:Pathological significance[J].Front Biosci,2010,6:12-25.
[2]Liu Y,Gampert L,Nething K,et al.Response and function of skeletal muscle heat shock protein 70[J].Front Biosci,2006,11:2802-2827.
[3]Beutlar E.Red cell metabolism:A Manual of Biochemical Methods[M].3rd Edition.Orlando:Grune & Stratton,Inc.1984:195-203.
[4]Liu Y,Gampert L,Nething K,et al.Response and function of skeletal muscle heat shock protein 70[J].Front Biosci,2006,11:2802-2827.
[5]Liu Y,Lormes W,Wang L,et al.Different skeletal muscle HSP70 responses to high-intensity strength training and low-intensity endurance training[J].Eur J Appl Physiol,2004,91:330-335.
[6]Liu Y,Steinacker JM.Changes in skeletal muscle heat shock proteins:pathological significance[J].Front Biosci,2001,6:12-25.
[7]Febbraio MA,Steensberg A,Walsh R,et al.Reduced glycogen availability is associated with an elevation in HSP72 in contracting human skeletal muscle[J].J Physiol,2002,538:911-917.
[8]wang L,Liu Y,Steinacker JM.Hsp70 response to electrical stimulation in C2C12 cells[J].Exerc Immunol Rev,2010,18:91-102.
[9]Wang L,Schumann U,Liu Y.Heat shock protein 70(Hsp70)inhibits oxidative phosphorylation and compensates ATP balance through enhanced glycolytic activity[J].J Appl Physiol,2012,113:1669-1676.
[10]Rodnick KJ,Henriksen EJ,James DE,et al.Exercise training glucose transporter and glucose transport in rat skeletal muscle[J].Am J Physiol,1992,262:9-14.
[11]杨晓冰,吴毅,李益明,等.运动促进大鼠骨骼肌细胞葡萄糖运载体4 的转位[J].中国运动医学杂志,2000,19:37-38.
