正性情绪诱导下的自我参照加工:来自ERPs的证据*
2014-02-06钟毅平蔡荣华谭千保肖丽辉占友龙秦敏辉
钟毅平 范 伟 蔡荣华 谭千保 肖丽辉 占友龙 罗 西 秦敏辉
(湖南师范大学教育科学学院,长沙 410082)
1 引言
自我参照效应(self-reference effect)是指当信息与自我概念有关时,个体会进行快速的加工和更好的记忆(Rogers,Kuiper,&Kirker,1977;Kim,2012)。心理学家们普遍认为,在潜意识下的自我,它不仅仅对人类的情绪情感产生以及需要、动机等个性心理倾向的激发起着重要作用,而且还对人类的认知活动有着最直接影响。早在1977年,心理学家Rogers在扩展了前人研究的基础上,第一次用实验证实了自我相关联的刺激信息和其它刺激信息进行记忆时,自我相关联的刺激信息记忆成绩比其它刺激信息好,他把这种记忆的优势效应归纳为自我参照效应(Rogers et al.,1977)。从此以后,越来越多的研究者对自我、记忆等心理活动产生了浓厚的兴趣,越来越多的研究者将这两个概念结合起来进行实验研究。
大量的中西方研究关注的是自我的类型,同一类型的自我参照加工应该有着同样的神经机制(Berlad&Pratt,1995;Miyakoshi,Nomura,&Ohira,2007;Ninomiya,Onitsuka,Chen,Sato,&Tashiro,1998;Schneider et al.,2012;Symons&Johnson,1997;Zhao,Wu,Zimmer,&Fu,2011)。对自我四肢归属的加工(Ehrsson,Spence,&Passingham,2004;Feinberg,1997;Meador,Loring,Feinberg,Lee,&Nichols,2000),还有面孔的再认(Platek et al.,2006;Turk et al.,2002;Liew,Ma,Han,&Aziz-Zadeh,2011;Ma&Han,2012a;Ma&Han,2012b;Sui,Hong,Hong Liu,Humphreys,&Han,2013),手的再认(Ferri et al.,2012;Su et al.,2010)等探索的是身体的自我;此外,以及使用自己的声音(Kaplan,Aziz-Zadeh,Uddin,&Iacoboni,2008),自己的笔迹(Chen et al.,2008),特征形容词(Craik et al.,1999;Kelley et al.,2002;Schneider et al.,2012;Serino,Pizzoferrato,&Làdavas,2008;Han et al.,2009)和自传体记忆(Finketal.,1996;Gray,Ambady,Lowenthal,&Deldin,2004;Kim,2012;Summerfield,Hassabis,&Maguire,2009)等探索的是心理的自我。虽然自我概念包含不同的层面,但是心理自我被认为是自我的核心层面(Northoff et al.,2006)。同时,还有学者认为自我类别应该包括个体自我(individualself)和集体自我(collectiveself)(Trafimow,Triandis,&Goto,1991)。个体自我包括自己独有的状态、特征和行为等(如快乐的,聪明的,勤奋的),集体自我包括集体成员身份、社会联系和社会地位等(如兄长,儿子,公民)(Gaertner et al.,2012;Northoff et al.,2006)。身体四肢归属的加工(Ehrsson et al.,2004;Feinberg,1997;Meador et al.,2000),对于自己的面孔、手的再认等(Ferri et al.,2012;Platek et al.,2006;Su et al.,2010;Turk et al.,2002)也被认为探索的是个体自我。类似地,也有一系列的研究探索了集体自我的存在(Brewer,1996;Sedikides&Brewer,2001;Trafimow 等,1991)。例如,在前人的研究中,被试的大学和家庭被当作集体自我参照刺激,相对于语义参照编码,集体自我参照编码被试再认成绩更好(Johnson et al.,2002);集体自我参照的记忆优势效应也得到实验证明,集体自我有关的信息比非集体自我有关信息有更好的记忆(Wagar&Cohen,2003);张力等(2006)用自我参照效应范式完成了一项fMRI研究,发现中国人的母亲参照与自我参照共同激活了内侧前额叶(mPFC),从而推论母亲是中国人集体自我的一个组成部分(Zhang et al.,2006)。赵科等人对个体自我参照效应和集体自我参照效应进行了研究,发现个体自我参照效应要大于集体参照效应(Zhao et al.,2009)。
通过对文献的回顾,我们发现,前人的研究大多是以自我相关和非自我刺激去激活被试行为和神经层面上的差异来考察自我相关效应,但并没有考虑到这些刺激与自我的相关程度如何。在现实生活中,刺激与自我的接近程度不同,对于个体而言往往意义也会不一样,对高自我相关刺激的加工比低自我相关刺激的加工具有更大的生理和社会意义。这就是为什么同样都是名字,而且都能比无关自我刺激引起更大的注意,但是听到自己的名字会比听到朋友的名字引起更大的警觉性。大脑不同的反应,可能反映了面对不同自我相关程度的刺激时,大脑的活动是存在差异的。
在以往的研究中,有一些关于自我面孔加工的研究与此相关。Ma和Han比较了被试自己的面孔和朋友的面孔,结果发现左右梭状回在自我面孔的物理属性和自我面孔识别上的功能是不一样的(Ma&Han,2012a)。他们还发现在低威胁状态下,自我面孔的优势效应是存在的,但是在高威胁状态下,自我面孔就不存在优势效应了(Ma&Han,2009)。Keyes和他的同事比较了被试自己的面孔,朋友的面孔以及陌生人的面孔,发现被试对于自己面孔的加工更为特别一些(Keyes,Brady,Reilly,&Foxe,2010)。而Scott等人比较了成年人和四岁的儿童对于面孔加工的电生理差别,结果发现儿童对于自己面孔的加工表现出更多的弥漫性电活动(Scott,Luciana,Wewerka,&Nelson,2005)。Sui等人的研究和Caharel等人的研究也表明了人脑对自己面孔的认知加工是一种自动的加工,发生在面孔结构编码之后,并且这种加工是独立进行的,与任务无关(Caharel,Courtay,Bernard,Lalonde,&Rebai,2005;Sui,Zhu,&Han,2006)。另外,还有一些关于自己名字的研究也与不同自我相关程度的刺激加工有关。Tacikowski等人考察了在预实验中名字和面孔的重复效应,研究结果发现对比名字来说,被试对面孔的学习更容易,这可能是由于面孔携带了更多了语义信息(Tacikowski,Jednoróg,Marchewka,&Nowicka,2011)。Tacikowski和同事还发现对比名人和陌生人来说,无论在听觉模式下,还是视觉模式下,个体自己相关的信息总能激活内侧前额叶皮质(MPFC)(Tacikowski,Brechmann,& Nowicka,2012)。Tateuchi等人也发现,相比其他的名字,被试听到自己的名字时有一个早期注意的加工(Tateuchi,Itoh,&Nakada,2012)。另外,Höller等人也发现,相比其它条件,被试自己名字所诱发的α-β波的活动更弱。但他们认为这一结果与熟悉度有关,而不是与自我相关程度有关(Höller et al.,2011)。Chen等人直接考查了自我相关加工的程度效应,结果发现被试自己的名字比所在的省份的名字和国家的名字激发了更大的P3波幅(Chen et al.,2011)。但是个人的名字与省份的名字概念内涵不同,这种比较可能并不是在同一层面上的,更大的P3波幅可能是由于特异程度不同引起的,而不是真正意义上的自我相关程度的不同。基于这些发现和证据,研究者认为与自我相关的刺激应该还有着程度的不同,与核心自我的接近程度不同,大脑对其加工深度也不同。即与低自我相关的刺激相比,高自我相关的刺激会得到大脑更好更精细的加工。 然而,自我参照加工的程度效应是可以直接被观察到的。尤其是高低不同的自我相关刺激,大脑对其加工的时空特性可能是不一样的,这是一个值得考察的命题。
更有意思的是,在情绪的研究中也有类似的程度效应存在。有研究发现虽然对比中性刺激,情绪刺激都能吸引更多的注意力,但是极端情绪刺激总能比中等程度的情绪刺激诱发更多的大脑注意偏向(Yuan,Yang,Meng,Yu,&Li,2008;Yuan et al.,2007)。何媛媛等人也发现,外倾被试组在极端正性刺激、中等正性刺激、中性刺激三种条件下 P2波幅两两差异显著,且极端正性刺激诱发的P2波幅最大,中等正性刺激诱发的P2波幅次之,中性刺激诱发的P2波幅最小(何媛媛,袁加锦,伍泽莲,李红,2008)。还有研究表明相比低负性情绪,高负性情绪对个体的生活影响则更大。高负性的情绪会严重影响个体的记忆,阻碍创造力的发挥,也很容易导致个体做出不明智的决策(Coon&Mitterer,2010;Watkins,Vache,Verney,&Mathews,1996;黄宇霞,罗跃嘉,2004)。事实上,自我相关刺激和情绪唤醒刺激也有相似的地方,尤其是在生理意义这一方面(Gray et al.,2004;Phan et al.,2004)。有研究表明对自我相关的刺激和情绪刺激进行加工能够激活一些相似的神经机制,例如,伏隔核(nucleus acumbens),脑岛(insula),腹内侧前额叶(VMPFC)等(Phan et al.,2004;Taylor&Fragopanagos,2005)。还有研究表明,自我相关刺激和情绪刺激的加工相互有着重要的影响。例如,Watson和Dritschel等人运用自我参照范式,在被试对情绪性字词进行自我相关判断(像我VS不像我)时记录其脑电,发现N400上自我相关信息加工与负性情绪效价加工之间存在交互作用(Watson,Dritschel,Obonsawin,&Jentzsch,2007)。Watson和Dritschel等人比较了抑郁被试和正常被试对情绪字词的自我相关判断,结果,发现正常被试身上存在自我正面偏见,而抑郁被试虽然可以准确识别字词的情绪效价,但是却不能把情绪效价与其自我联系起来,他们认为情绪状态的改变(如抑郁)跟自我与情绪的关系的改变是相联系的(Watson,Dritschel,Jentzsch,&Obonsawin,2008)。陈芸、钟毅平等人采用了阈下启动范式,给定被试情绪词判断任务,这个核心反应与无关反应相冲突,所诱发的N2反映了内隐层面上自我与情绪存在交互作用(陈芸、 钟毅平、 周海波、 周路平和王小艳,2012)。
另外,自我相关的刺激和情绪刺激的加工不但相互有着重要的影响,能够激活一些相似的神经机制,而且能够激发同样的ERPs成分。有大量的研究发现情绪刺激总是能够引起P3的变化(Chen,Yuan,Huang,Chen,&Li,2008;Huang&Luo,2006;Ito&Cacioppo,2000;Ito,Larsen,Smith,&Cacioppo,1998),带情绪色彩的刺激总是能比中性刺激诱发更大的P3波幅和更长的潜伏期。而且前人的研究也发现,自我参照刺激总是比控制刺激能够诱发更大的P3波幅(Miyakoshi et al.,2007;Ninomiya et al.,1998)。在我们实验室也有两个研究发现了类似的效应,一个研究发现个体自己的名字和被试母校的名字比其他学校的名字诱发了更大的P3波幅和更长的P3潜伏期(Zhao et al.,2009);另一个研究发现以本国国旗为例的自我参照刺激比其他国家的国旗诱发了更大的P3波幅和更长的P3潜伏期(Fan et al.,2011)。既然情绪与自我参照加工不但有着重叠的神经机制,还能诱发相同的ERPs波形指标,那么可以预测,情绪与自我参照加工是有着某种重要联系的,只是这种联系的内部机制我们还不太清楚。
前人的研究表明,除了P3成分以外,N2成分也被认为与自我加工高度相关(Chen et al.,2011;Chen et al.,2012;Fan et al.,2011;Zhao et al,2011)。Fan等人的研究表明早在刺激呈现后200 ms左右,人脑已经对自我参照刺激进行加工了,而不是大部分研究中认为的P3阶段(Fan et al.,2011)。Zhao等人的研究表明人脑对自己名字的加工更为敏感,比其他名字的加工更早一些,即自我名字加工存在N250效应(Zhao et al.,2011)。研究者预测在当前的研究中,早在N2阶段自我参照效应就会出现,高自我相关的刺激会比其它刺激得到更快的加工,但这一加工应该是粗糙的,更为精细的认知加工会发生在稍晚的阶段(比如P3阶段)。
综上所述,前人对于情绪与自我加工的关系研究主要集中在负性情绪对自我加工的影响上,较少有研究考察正性情绪对自我加工的影响。但人脑对于正性与负性情绪的加工肯定有所不同,例如Pourtois,Grandjean,Sander,&Vuilleumier(2004)观察到恐惧表情相对高兴表情引起较大的C1成分;恐惧表情消失后在同一位置呈现的刺激引起较大P1波幅,而高兴表情不能引起这种效应(Pourtois et al.,2004)。Pourtois和他的同事在随后的实验中运用相似实验范式做了一项fMRI研究(Pourtois,Schwartz,Seghier,Lazeyras,&Vuilleumier,2006),研究发现,恐惧面孔比高兴面孔引起大脑双侧颞顶区和右侧枕顶区更强的激活。根据这些证据,可以预测正性情绪刺激对自我参照加工的影响不同于负性情绪刺激,它的作用机制与神经基础也可能是不同的。而且,有研究表明自我相关信息是与正性情绪效价联系在一起的,个体往往把正面的结果或特质归因于自我内部稳定的人格特征,而认为负面的结果或特质与自己的人格特征不相关(Mezulis,Hyde,&Clark,2004;Pahl&Eiser,2005)。所以, 研究者认为探索正性情绪刺激与自我参照加工的关系更有意义,这不仅有助于验证和完善自我相关信息与情绪信息的功能加工层次模型,有助于从内隐层面上了解自我与情绪的复杂关系,揭示不同情绪状态下内隐自我参照加工的性质与特征,而且还能将对其神经基础做出重要探索,以达到深入解读自我的目的。
2 方法
2.1 实验目的与实验假设
采用图片启动范式,以探讨正性情绪对自我参照加工程度效应的影响。实验假设是人脑对正性情绪与自我参照刺激的加工可能是相互独立的系统,正性情绪的诱导较少地影响到自我参照加工的程度效应;早在N2阶段就会出现自我参照效应,高自我相关的名字比其他名字加工更快;而自我参照加工的程度效应可能会出现在P3阶段,在这一阶段高自我相关的名字比中等自我相关的名字和非自我名字诱发了更大的P3波幅,而中等自我相关的名字又比非自我名字诱发了更大的P3波幅。
2.2 被试
20名在校大学生和研究生参与了脑电实验,男生9名,女生11名,年龄在19~25岁之间,平均为21.58岁。被试均为右利手,身体健康,无神经系统疾病,没有脑部损伤史,视力正常或校正后正常。实验完成后付给一定报酬。
2.3 实验材料
采用正性图片与中性图片作为启动刺激,不同相关程度的自我参照刺激作为目标刺激。
120张正性与中性图片组成启动刺激。采用图片启动范式,从中国情绪图片系统(CAPS)中选取60张正性情绪图片和60张中性情绪图片。正性和中性图片在效价强度上相匹配,正性图片效价强度平均为7.56±0.27,中性图片效价强度为5.14±0.11,两者在效价强度上有显著的差异,F
(1,59)=3.49,p
<0.05;正性和中性图片在唤醒度上也相匹配,正性图片唤醒度平均为5.69±0.60,中性图片的唤醒度平均为3.93±0.97,两者在唤醒度上有显著的差异,F
(1,59)=4.12,p
<0.05;正性和中性图片在熟悉度上保持一致,两者在熟悉度上的差异不显著,F
(1,59)=0.21,p
>0.05。实验时,将两种图片都重复使用两次,共计360张图片次,然后将正性情绪和中性图片平均分配成3组,每组有120张图片次,包括取60张正性情绪图片次和60张中性情绪图片次。匹配好各组图片的效价强度、唤醒度和熟悉度,使其没有显著的差异。 统计结果显示启动刺激平衡控制较好,3组正性图片之间的效价强度[F(
1,59)=0.31,p
>0.05]、熟悉度[F(
1,59)=0.39,p
>0.05]和唤醒度[F
(1,59)=0.69,p
>0.05]都不存在显著的差异,三组中性图片之间的效价强度[F(
1,59)=0.27,p
>0.05]、熟悉度[F
(1,59)=0.19,p
>0.05]和唤醒度[F
(1,59)=0.11,p
>0.05]也不存在显著的差异。自我参照刺激按相关程度分为三类:一是与自我高相关的材料,即被试自己的名字(例如:秦敏辉);二是与自我中度相关的材料,即被试父亲的名字(例如:秦某某);三是熟悉刺激,即熟悉但不具备归属感的外国国家领导人的名字作为熟悉的刺激(例如:奥巴马)。每类自我参照刺激呈现60次,其中30次蓝色字体,30次为绿色字体。
2.4 实验设计
3(自我相关类型:自我高相关、自我中度相关和熟悉刺激)×2(情绪效价:正、中性)两因素被试内设计,因变量为ERP数据(P2和N2的波幅与潜伏期,P3平均波幅)。
2.5 实验程序
正式实验时,首先在屏幕中央呈现一个200 ms的”十”字注视点,紧接着随机呈现500~1000 ms的黑屏,之后呈现500 ms的一种情绪图片,然后黑屏随机呈现150~300 ms,再呈现500 ms的一种自我参照刺激,最后再呈现1000 ms的黑屏。在呈现自我参照刺激时,被试的任务是认真看刺激并进行分类按键反应,且要求又快又准。如果被试认为自我参照刺激字体是蓝色,则按“1”键;如果被试认为自我参照刺激字体是绿色,则按“2”键;练习实验之后,后跟360试次的正式实验,分3组,每组120试次(见图1)。

图1 实验程序中一个试次的流程图
为了测量刺激的熟悉度和有效性,要求被试在实验后对三类自我相关刺激的自我相关程度和熟悉度进行评定,采用口头报告的9点量表。在自我相关程度的评定中,“1”代表完全不相关,“9”代表非常相关。在熟悉度的评定中,“1”代表完全不熟悉,“9”代表非常熟悉。
2.6 脑电记录
使用 NeuroScan ERP记录与分析系统,按国际10~20系统扩展的64导电极帽记录EEG。在线纪录时以左侧乳突连线为参考电极,离线后转为双侧乳突为参考电极,离线双眼外侧安置电极记录水平眼电(HEOG),左眼上下安置电极记录垂直眼电(VEOG)。滤波带通为0.05~40 Hz,采样频率为500 Hz/导, 头皮阻抗<5 kΩ。
2.7 ERP数据处理与统计
完成连续记录EEG后,离线(off-line)处理数据,用NeuroScan软件校正VEOG,并充分排除其它伪迹。对不同相关程度的自我参照刺激呈现后的脑电数据进行分析与EEG叠加。波幅大于±80 μV 者视为伪迹而自动剔除。分析时程(epoch)为刺激后550 ms,基线为刺激呈现前100 ms。根据已有研究结果与本研究目的,对ERP数据(P2、N200波幅与潜伏期、LPP平均波幅)进行统计分析。选择15个电极位置,进行四因素重复测量方差分析,3(自我相关类型:自我高相关、自我中度相关和熟悉刺激)×2(情绪效价:正、中性)×3单侧化(左脑:F3,FC3,C3,CP3,P3;中脑:Fz,FCz,Cz,CPz,Pz;右脑:F4,FC4,C4,CP4,P4)×5电极(头皮前部:F3,Fz,F4;头皮前中:FC3,FCz,FC4;头皮中部:C3,Cz,C4;头皮后中:CP3,CPz,CP4;头皮后部:P3,Pz,P4)。方差分析的P值采用 Greenhouse-Geisser法校正。
3 结果
3.1 行为结果
实验后的评定表明自我相关刺激存在着一个显著的主效应,F
(2,38)=38.05,p
<0.001;事后比较表明高自我相关名字比中等自我相关名字以及非自我名字评定得分更高,allt
(19)>3.31,allp
<0.05;而中等自我相关名字比非自我名字要高一些,t
(19)=9.5,p
<0.01。此外,三类自我相关刺激在熟悉度方面的评定没有发现显著的差异,F
(2,38)=1.03,p
>0.05。各条件下的正确率见表1。研究者对各条件下的正确率进行了方差分析,但并没有发现任何显著的主效应或交互作用。
3.2 ERPs结果
见图2和图3,在三类刺激中都发现了N1,P2,N2和P3成分。重复测量方差分析表明,N1在潜伏期和波幅上都没有发现显著的差异。
3.2.1 P2
在P2的波幅上,重复测量方差分析表明,情绪类型主效应显著,中性情绪条件比正性情绪条件激发了更大的P2波幅,F
(1,19)=28.77,p
<0.001。重复测量方差分析还发现,情绪类型与电极位置交互作用显著,F
(4,76)=13.20,p
<0.001。简单效应分析表明,在电极前中部位置,中性情绪条件所诱发的P2波幅显著高于正性情绪条件,F
(1,19)=3.82,p
<0.05。另外,自我参照类型与脑区位置交互作用显著,F
(4,76)=3.79,p
<0.05;简单效应分析表明,高自我相关的名字在右半球激发的P2波幅显著高于左半球,F
(1,19)=3.97,p
<0.05。在P2的潜伏期上,重复测量方差分析表明,自我参照类型与电极位置交互作用显著,F
(8,152)=3.41,p
<0.05;简单效应分析表明在电极后部位置,高自我相关的刺激比其它刺激诱发了更短的P2潜伏期,F
(1,19)=5.69,p
<0.01。
表1 各条件下正确率(%,M±SE)
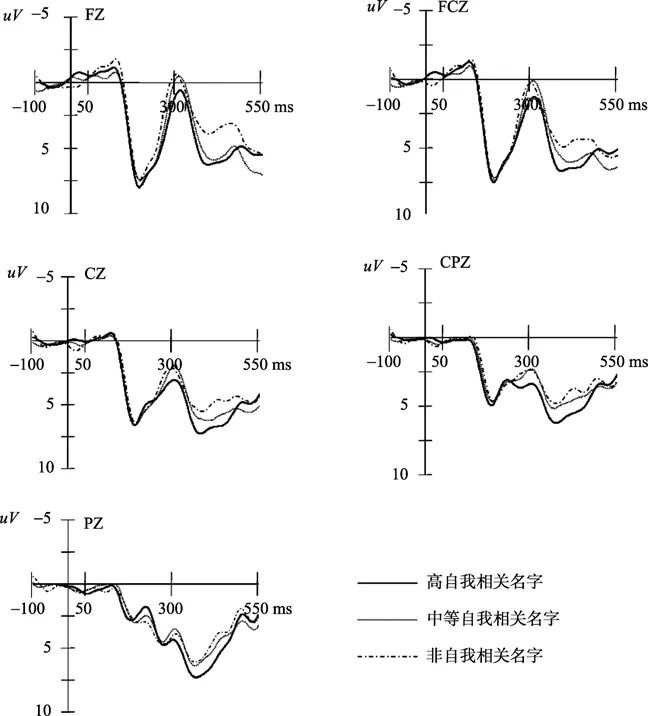
图2 正性图片启动下高自我相关名字、 中等自我相关名字和非自我相关名字在FZ、FCZ、CZ、CPZ和PZ上诱发的ERPs总平均图。
3.2.2 N2
在N2的波幅上,重复测量方差分析表明,自我参照刺激主效应边缘显著,F
(2,38)=3.58,p
=0.075。事后比较表明高自我相关名字比中等自我相关名字(p
<0.05)和非自我相关名字(p
<0.05)诱发了更小的N2的波幅。重复测量方差分析还显示,自我参照类型与电极位置交互作用显著,F
(8,152)=4.76,p
<0.01。简单效应分析表明高自我相关名字所诱发N2的波幅在前部额区电极位置[F
(1,19)=4.11,p
<0.05]、前中部电极[F
(1,19)=4.38,p
<0.05]、中部电极[F
(1,19)=6.17,p
<0.01]和头皮中后部电极[F
(1,19)=2.94,p
<0.05]显著小于其它条件。在N2的潜伏期上,重复测量方差分析表明,情绪类型与电极位置交互作用边缘显著,F
(4,76)=2.61,p
=0.06。简单效应分析表明,中性刺激所诱发的N2的潜伏期在额区电极位要长于其他的电极位置,F
(1,19)=2.31,p
<0.05。重复测量方差分析表明,自我参照类型与电极位置交互作用显著,F
(8,152)=10.66,p
<0.001。简单效应分析表明,高自我相关刺激在电极前部位置[F
(1,19)=3.49,p
<0.05]和电极后部位置[F
(1,19)=18.10,p
<0.001]所诱发的N2的潜伏期要显著长于其它刺激条件。3.2.3 P3(350~500 ms)
在P3平均波幅上,重复测量方差分析表明,自我参照刺激主效应显著[F
(2,38)=13.92,p
<0.001]。事后比较表明高自我相关名字比中等自我相关名字(p
<0.05)和非自我相关名字(p
<0.001)诱发了更大的P3波幅,中等自我相关名字比非自我相关名字诱发了更大的P3波幅(p
<0.01)。4 讨论
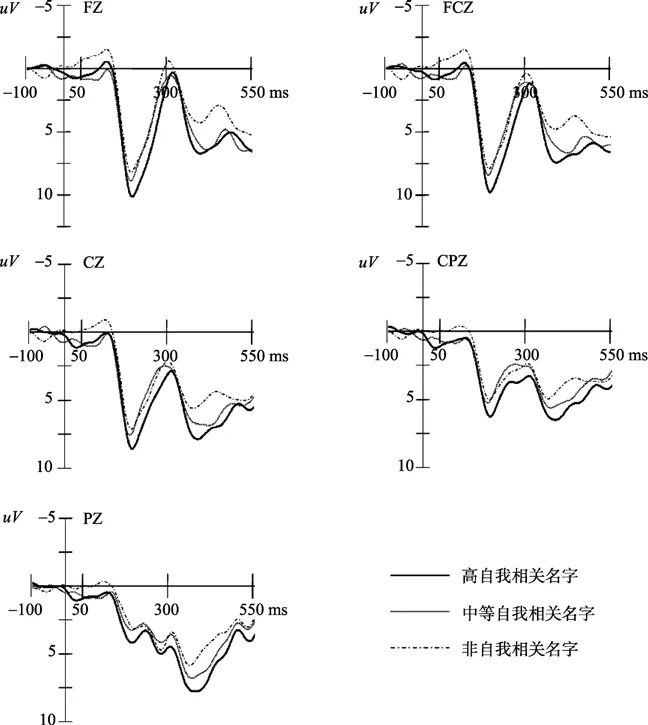
图3 中性图片启动下高自我相关名字,中等自我相关名字和非自我相关名字在FZ、FCZ、CZ、CPZ和PZ上诱发的ERPs总平均图。
在当前的研究中,研究结果发现三类自我参照刺激在N1的波幅和潜伏期上均没有发现显著的差异,这可能是由于对刺激类型的尺寸大小、长度和复杂性等物理属性上做到了均衡处理。在这一时间阶段,正性与中性情绪图片启动也没有影响到被试对自我参照刺激的加工,我们估计这种影响可能会出现在稍晚一点的阶段。
而在刺激呈现后,研究结果发现在额区电极观察到了明显的P2成分.对比正性图片,中性图片启动后的自我参照加工得到了更大的P2波幅。有研究表明,在额区位置发现的P2成分可能是对刺激特征的快速侦查,易敏感的刺激会得更多的关注(Karayanidis&Michie,1996;Thorpe,Fize,&Marlot,1996)。此外,还有研究表明与情绪有关的刺激可能比中性刺激激发更大的P2波幅,生物重要性更强的刺激容易得到注意(Carretié,Mercado,Tapia,&Hinojosa,2001)。但是这种注意缺乏高级的认知加工和控制资源的分配(Del Cul,Baillet,&Dehaene,2007;Hu,Wu,&Fu,2011)。因此,中性图片启动后的自我参照加工更大,可能是由于正性图片呈现时,吸引了大量的注意资源,而人脑对中性图片的加工消耗的认知资源较小,对后面呈现的自我参照刺激加工影响也较小,自我参照刺激得到了充分的加工。还有证据表明,个体的名字直接代表了他(她)自己,因此有着很强的重要性(Chen et al.,2011;Perrin et al.,2005;Zhao et al.,2011)。在当前的研究中,对比其他的名字,高自我相关的名字所诱发的P2潜伏期更短,这也可能是由于其生物重要性,大脑识别高自我相关的名字更为容易,消耗的认知资源更少一些(Campanella et al.,2002)。中等自我相关名字和低自我相关名字的自我参照加工在这一阶段并没有发现显著的差异,这可能是由于人脑对不同程度的自我参照刺激的加工和提取发生在稍晚的阶段。
在额区位置的电极还发现了比较明显的N2成分,高自我相关的名字比其他名字诱发了更小的N2波幅。有研究表明被试自我的面孔比其它刺激诱发了更小的N2波幅(Herzmann,Schweinberger,Sommer,&Jentzsch,2004)。N2通常被认为是一种非特殊的成分,它与注意的转换机制相联系,随后一般会有P3成分出现(Kiehl,Laurens,Duty,Forster,&Liddle,2001)。在被动接受刺激的情况下,只要刺激足够突出或者发生了注意的转换,N2b/P3a都会被观察到(Näätänen,Simpson,&Loveless,1982)。因此,在当前的研究中,被试自己的名字由于其突出的生物和社会意义上的重要性很快抓住了大脑的注意力。中等自我相关名字和非自我相关名字的自我参照加工在这一阶段都没有发现显著的差异,这可能是由于在这一阶段人脑对不同程度的自我参照刺激的加工也是粗糙的,更为精细的认知加工会发生在稍晚的阶段。另外,研究结果发现高自我相关名字比其他名字所诱发的N2潜伏期更短,这可能是由于高自我相关的名字可能具有更好的心理突出性(psychologically salient)和生理重要性(biologically important),大脑识别起来更为容易,消耗的认知资源更少一些(Campanella et al.,2002)。
在两种情绪图片启动下,都能观察到自我参照刺激具有显著差异的P3成分。P3通常被认为是人脑在晚期阶段对中枢控制加工的朝向反应(Campanella et al., 2002; Carretié, Hinojosa,Martín‐Loeches,Mercado,&Tapia,2004;Yuan et al.,2008)。对比低自我相关的刺激,人脑在加工高自我相关的刺激时会投入更多的认知和控制加工资源。因此,在实验中高自我相关的名字比中等自我相关的名字诱发了更大的P3波幅,而中等自我相关的名字又比非自我相关的名字诱发了更大的P3波幅。这些结果证明了一个显著的自我参照加工程度效应,即高自我相关的刺激会比低自我相关的刺激得到更为深入和精细的加工。这些结果还表明P3成分不同于P2和N2成分,它可能反应了自我信息的总体和精细的加工过程,能够作为自我参照加工程度效应的有效ERPs指标。
另外,前人的研究报告了熟悉刺激会比不熟悉的刺激诱发更大的P3波幅(Beauchemin et al.,2006;Bobes,Martín,Olivares,&Valdés-Sosa,2000;Zhao et al.,2009)。如果刺激的熟悉程度不一样,可能会对自我相关程度的研究造成污染。但行为学评定结果已经将这种可能排除了,所有的自我相关刺激和非自我相关刺激在熟悉程度上没有显著的差异。因此,实验中所发现的P3可能无关熟悉度。而且,研究结果表明P3可能与自我相关程度有着显著的相关,自我相关程度越高,P3的波幅越大,而N2成分不存在这种相关和影响,所以实验中发现的P3可能不是特征刺激或者新异刺激的指标,而是反应了自我相关程度不同刺激的特征指标。
研究结果还表明,无论在正性情绪启动还是中性启动的影响下,自我参照加工的程度效应都存在,表现出稳定的特征。另外,在早期阶段(150~250 ms),对比正性图片启动后的自我参照加工,中性图片启动后的自我参照加工得到了更大的P2波幅。而在稍晚的N2和更晚的P3阶段,却没有发现这一效应。这可能表明正性图片启动所起到的干扰作用是有限的,这与前人的研究结果相似。Yuan等人的研究表明,在内隐情绪任务中,被试的 ERP波幅在极端正性、 中等正性和中性三种效价强度图片刺激的条件下无显著差异(Yuan et al.,2007)。当被试在执行外显的情绪分类任务时,增加强度的愉悦表情也不能导致 ERP波幅明显的变化(Leppänen,Kauppinen,Peltola,&Hietanen,2007)。这些研究得出这样一个结论,人类大脑对正性情绪刺激的加工并不敏感。所以,无论在正性情绪启动还是中性启动的影响下,自我参照加工的程度效应都能展现出稳定的特征。
总的来说,当前的研究借助高时间进程的精密仪器ERPs,不但重复了自我参照加工的程度效应,而且拓展了前人的研究,揭示了正性情绪对自我参照加工程度效应的影响。正性情绪对自我参照加工程度效应的影响是有限的,二者可能有着独立的加工系统,互不干扰。无论在正性情绪还是中性情绪的诱导下,高自我相关的刺激都得到了更为深入和精细的加工,自我参照加工的程度效应表现出了稳定的特征。
Beauchemin,M.,De Beaumont,L.,Vannasing,P.,Turcotte,A.,Arcand, C., Belin, P., & Lassonde, M. (2006).Electrophysiological markers of voice familiarity.European Journal of Neuroscience,23
(11),3081–3086.Berlad,I.,&Pratt,H.(1995).P300 in response to the subject's own name.Electroencephalography
and
Clinical Neurophysiology/Evoked
Potentials
Section,
96
(5),472–474.Bobes,M.A.,Martín,M.,Olivares,E.,&Valdés-Sosa,M.(2000).Different scalp topography of brain potentials related to expression and identity matching offaces.Cognitive Brain Research,9
,249–260.Brewer,M.(1996).In-group favoritism:The subtle side of intergroup discrimination.In D.Messick&A.Tenbrunsel(Eds.),Codes
ofconduct:BehavioralResearch
Into Business Ethics
(pp.160–170).New York:Russell Sage Foundation.Caharel,S.,Courtay,N.,Bernard,C.,Lalonde,R.,&Rebai,M.(2005).Familiarity and emotional expression influence an early stage of face processing:An electrophysiological study.[Clinical Trial].Brain and Cognition,59
(1),96–100.Campanella,S.,Gaspard,C.,Debatisse,D.,Bruyer,R.,Crommelinck,M.,&Guerit,J.(2002).Discrimination of emotional facial expressions in a visual oddball task:An ERP study.Biological Psychology,59
(3),171–186.Carretié,L.,Hinojosa,J.A.,Martín-Loeches,M.,Mercado,F.,&Tapia,M.(2004).Automatic attention to emotional stimuli:Neural correlates.Human Brain Mapping,22
(4),290–299.Carretié,L.,Mercado,F.,Tapia,M.,&Hinojosa,J.A.(2001).Emotion,attention and the "negativity bias",studied through event-related potentials.International Journal of Psychophysiology,41
(57),75–85.Chen,A.,Weng,X.,Yuan,J.,Lei,X.,Qiu,J.,Yao,D.,&Li,H.(2008).The temporal features of self-referential processing evoked by Chinese handwriting.Journal of Cognitive Neuroscience,20
(5),816–827.Chen,J.,Yuan,J.,Feng,T.,Chen,A.,Gu,B.,&Li,H.(2011).Temporal features of the degree effect in self-relevance:Neural correlates.Biological Psychology,87
(2),290–295.Chen,J.,Yuan,J.,Huang,H.,Chen,C.M.,&Li,H.(2008).Music-induced mood modulates the strength of emotional negativity bias:An ERP study.Neuroscience
Letters,445
(2),135–139.Chen,J.,Zhong,J.,Zhang,Y.X.,Li,P.,Zhang,A.,Tan,Q.B.,& Li,H.(2012).Electrophysiologicalcorrelates of processing facialattractiveness and its influence on cooperative behavior.Neuroscience Letters,517
(2),65–70.Chen,Y.,Zhong,Y.P.,Zhou,H.B.,Zhou,L.P.,&Wang,X.Y.(2008).An ERP Study on Implicit Self-positivity Bias Effect.Chinese Journal of Clinical Psychology
,20
(3),297–300.[陈芸,钟毅平,周海波,周路平,王小艳.(2012).内隐自我正面偏见效应的 ERP研究.中国临床心理学杂志,20
(3),297–300.]Coon,D.,&Mitterer,J.O.(2010).Introduction to psychology:Gateways to mind and behavior
(12th ed.).
Belmont,CA:Wadsworth.Craik,F.I.M.,Moroz,T.M.,Moscovitch,M.,Stuss,D.T.,Winocur,G.,Tulving,E.,&Kapur,S.(1999).In search of the self: A positron emission tomography study.Psychological Science,10
(1),26–34.Del Cul,A.,Baillet,S.,&Dehaene,S.(2007).Brain dynamics underlying the nonlinear threshold for access to consciousness.PLoS Biology,5
(10),2408–2423.Ehrsson,H.,Spence,C.,&Passingham,R.(2004).That's my hand!Activity in premotorcortex reflectsfeeling of ownership of a limb.Science,305
(5685),875.Fan,W.,Zhang,Y.X.,Wang,X.,Wang,X.Y.,Zhang,X.Y.,&Zhong, Y. P. (2011). The temporal features of self-referential processing evoked by national flag.Neuroscience Letters,505
(3),233–237.Feinberg,T.(1997).Some interesting perturbations of the self in neurology.Seminars in Neurology,17
,129–134.Ferri,F.,Frassinetti,F.,Mastrangelo,F.,Salone,A.,Ferro,F.M.,&Gallese,V.(2012).Bodily self and schizophrenia:The loss of implicit self-body knowledge.Consciousness and Cognition,21
(3),1365–1374.Fink,G.,Markowitsch,H.,Reinkemeier,M.,Bruckbauer,T.,Kessler,J.,&Heiss,W.D.(1996).Cerebral representation of one's own past: Neural networks involved in autobiographical memory.Journal of Neuroscience,16
(13),4275–4282.Gaertner,L.,Sedikides,C.,Luke,M.,O'Mara,E.M.,Iuzzini,J.,Jackson,L.E.,...Wu,Q.(2012).A motivational hierarchy within:Primacy of the individual self,relational self,or collective self?Journal of Experimental Social Psychology,48
(5),997–1013.Gray,H.,Ambady,N.,Lowenthal,W.,&Deldin,P.(2004).P300 as an index of attention to self-relevant stimuli.Journal
of
Experimental
Social
Psychology,
40
(2),216–224.Höller,Y.,Kronbichler,M.,Bergmann,J.,Crone,J.S.,Schmid,E.V.,Golaszewski,S.,&Ladurner,G.(2011).Inter-individualvariability of oscillatory responses to subject's own name: A single-subject analysis.International
Journal
of
Psychophysiology,
80
(3),227–235.Han,S.,Gu,X.,Mao,L.,Ge,J.,Wang,G.,&Ma,Y.(2009).Neural substrates of self-referential processing in Chinese Buddhists.Social Cognitive and Affective Neuroscience,5
(2–3),332–339.He,Y.Y.,Yuan,J.J.,Wu,Z.L.,&Li,H.(2008).The valence strength of positive stimuli modulates the attention in extraverts study.Acta
Psychologica
Sinica,
40
(11),1158–1164.[何媛媛,袁加锦,伍泽莲,李红.(2008).正性情绪刺激效价强度的变化对外倾个体注意的调制作用.心理学报,40
(11),1158–1164.]Herzmann,G.,Schweinberger,S.R.,Sommer,W.,&Jentzsch,I.(2004).What's special about personally familiar faces?A multimodal approach.Psychophysiology,41
(5),688–701.Hu,X.,Wu,H.,& Fu,G.(2011).Temporal course of executive control when lying about self- and other-referential information: An ERP study.Brain Research,1369
,149–157.Huang,Y.X.,&Luo,Y.J.(2004).Emotion-related ERP components and their variety in mood disorder.Advances in Psychological Science,12
(1),10–17.[黄宇霞,罗跃嘉.(2004).情绪的ERP相关成分与心境障碍的ERP变化.心理科学进展,12
(1),10–17.]Huang,Y.X.,&Luo,Y.J.(2006).Temporal course of emotional negativity bias:An ERP study.Neuroscience Letters,398
(1–2),91–96.Ito,T.,&Cacioppo,J.(2000).Electrophysiological evidence of implicit and explicit categorization processes.Journal of Experimental Social Psychology,36
(6),660–676.Ito,T.,Larsen,J.T.,Smith,N.K.,&Cacioppo,J.T.(1998).Negative information weighs more heavily on the brain:The negativity bias in evaluative categorizations.Journal of Personality and Social Psychology,75
,887–900.Johnson,C.,Gadon,O.,Carlson,D.,Southwick,S.,Faith,M.,&Chalfin,J.(2002).Self reference and group membership:Evidence for a group reference effect.European Journal of Social Psychology,32
(2),261–274.Kaplan,J.,Aziz-Zadeh,L.,Uddin,L.,&Iacoboni,M.(2008).The self across the senses:An fMRI study of self-face and self-voice recognition.SocialCognitive
and
Affective Neuroscience,3
(3),218–223.Karayanidis,F.,& Michie,P.(1996).Frontal processing negativity in a visual selective attention task.Electroencephalography
and
ClinicalNeurophysiology,99
(1),38–56.Kelley,W.,Macrae,C.,Wyland,C.,Caglar,S.,Inati,S.,&Heatherton,T.(2002).Finding the self?An event-related fMRI study.Journal of Cognitive Neuroscience,14
(5),785–794.Keyes,H.,Brady,N.,Reilly,R.B.,&Foxe,J.J.(2010).My face or yours? Event-related potentialcorrelates of self-face processing.Brain and Cognition,72
(2),244–254.Kiehl,K.A.,Laurens,K.R.,Duty,T.L.,Forster,B.B.,&Liddle,P.F.(2001).Neural sources involved in auditory target detection and novelty processing:An event-related fMRI study.Psychophysiology,38
(1),133–142.Kim,H.(2012).A dual-subsystem model of the brain's default network:Self-referentialprocessing,memory retrieval processes, and autobiographical memory retrieval.Neuroimage,61
(4),966–977.Leppänen,J.M.,Kauppinen,P.,Peltola,M.J.,&Hietanen,J.K. (2007). Differential electrocortical responses to increasing intensities offearfuland happy emotional expressions.Brain Research,1166
,103–109.Liew,S.L.,Ma,Y.,Han,S.,&Aziz-Zadeh,L.(2011).Who's afraid of the boss:Cultural differences in social hierarchies modulate self-face recognition in Chinese and Americans.PLoS ONE,6
(2),e16901.Ma,Y.,&Han,S.(2009).Self-face advantage is modulated by social threat–Boss effect on self-face recognition.Journal of Experimental Social Psychology,45
(4),1048–1051.Ma,Y.,&Han,S.(2012a).Functional dissociation of the left and right fusiform gyrus in self-face recognition.Human Brain Mapping,33
(10),2255–2267.Ma,Y.,&Han,S.(2012b).Is the self always better than a friend?Self-Face recognition in Christians and Atheists.PLoS ONE,7
(5),e37824.Meador,K.J.,Loring,D.W.,Feinberg,T.E.,Lee,G.P.,&Nichols,M.E.(2000).Anosognosia and asomatognosia during intracarotid amobarbital inactivation.Neurology,55
(6),816–820.Mezulis,A.H.,Hyde,J.S.,&Clark,R.(2004).Father involvement moderates the effect of maternal depression during a child's infancy on child behavior problems in kindergarten.Journal
of
Family
Psychology
,18
(4),575–588.Miyakoshi,M.,Nomura,M.,&Ohira,H.(2007).An ERP study on self-relevantobjectrecognition.Brain
and Cognition,63
(2),182–189.Näätänen,R.,Simpson,M.,&Loveless,N.(1982).Stimulus deviance and evoked potentials.Biological Psychology,14
(1–2),53–98.Ninomiya,H.,Onitsuka,T.,Chen,C.,Sato,E.,&Tashiro,N.(1998).P300 in response to the subject'sown face.Psychiatry and Clinical Neurosciences,52
(5),519–522.Northoff,G.,Heinzel,A.,de Greck,M.,Bermpohl,F.,Dobrowolny,H.,&Panksepp,J.(2006).Self-referential processing in our brain--a meta-analysis of imaging studies on the self.Neuroimage,31
(1),440–457.Pahl,S.,&Eiser,J.R.(2005).Valence,comparison focus and self-positivity biases.Experimental Psychology(formerly Zeitschrift
für
Experimentelle
Psychologie),
52
(4),303–310.Perrin,F.,Maquet,P.,Peigneux,P.,Ruby,P.,Degueldre,C.,Balteau,E.,...Laureys,S.(2005).Neural mechanisms involved in the detection of our first name:A combined ERPs and PET study.Neuropsychologia,43
(1),12–19.Phan,K.L.,Taylor,S.F.,Welsh,R.C.,Ho,S.H.,Britton,J.C.,&Liberzon,I.(2004).Neural correlates of individual ratings of emotional salience:A trial-related fMRI study.Neuroimage,21
(2),768–780.Platek,S.,Loughead,J.,Gur,R.,Busch,S.,Ruparel,K.,Phend,N.,...Langleben,D.(2006).Neural substrates for functionally discriminating self-face from personally familiar faces.Human Brain Mapping,27
(2),91–98.Pourtois,G.,Grandjean,D.,Sander,D.,&Vuilleumier,P.(2004).Electrophysiological correlates of rapid spatial orienting towards fearful faces.Cerebral Cortex,14
(6),619–633.Pourtois,G.,Schwartz,S.,Seghier,M.L.,Lazeyras,F.,&Vuilleumier,P.(2006).Neuralsystems fororienting attention to the location of threat signals:An event-related fMRI study.Neuroimage,31
(2),920–933.Rogers,T.B.,Kuiper,N.A.,& Kirker,W.S.(1977).Self-reference and the encoding of personal information.JournalofPersonality
and
SocialPsychology,35
(9),677–688.Schneider,M.,Debbané,M.,Lagioia,A.,Salomon,R.,d'Argembeau,A.,&Eliez,S.(2012).Comparing the neural bases of self-referential processing in typically developing and 22q11.2 adolescents.Developmental
Cognitive Neuroscience,2
(2),277–289.Scott,L.S.,Luciana,M.,Wewerka,S.,&Nelson,C.A.(2005).Electrophysiological correlates of facial self-recognition in adults and children.Cogniţie,Creier,Comportament,IX(
3),211–238.Sedikides,C.,&Brewer,M.B.(Eds.).(2001).Individual self,relational self,collective self
.Philadelphia,PA:Psychology Press.Serino,A.,Pizzoferrato,F.,&Làdavas,E.(2008).Viewing a face(especially one's own face)being touched enhances tactile perception on the face.Psychological Science,19
(5),434–438.Su,Y.,Chen,A.,Yin,H.,Qiu,J.,Lv,J.,Wei,D.,...Wang,T.(2010). Spatiotemporal cortical activation underlying self-referencial processing evoked by self-hand.Biological Psychology,85
,219–225.Sui,J.,Hong,Y.Y.,Hong,L.C.,Humphreys,G.W.,&Han,S.(2013).Dynamic cultural modulation of neural responses to one's own and friend's faces.Social Cognitive Affective Neuroscience,8
(3),326–332.Sui,J.,Zhu,Y.,&Han,S.(2006).Self-face recognition in attended and unattended conditions:An event-related brain potential study.NeuroReport,17
(4),423–427.Summerfield,J.,Hassabis,D.,&Maguire,E.(2009).Cortical midline involvement in autobiographical memory.Neuroimage,44
(3),1188–1200.Symons,C.S.,&Johnson,B.T.(1997).The self-reference effect in memory:A meta-analysis.Psychological Bulletin,121
(3),371–394.Tacikowski,P.,Brechmann,A.,& Nowicka,A.(2012).Cross-modal pattern of brain activations associated with the processing of self-and significant other's name.Human Brain Mapping,34
,2069-2077.Tacikowski,P.,Jednoróg,K.,Marchewka,A.,&Nowicka,A.(2011).How multiple repetitions influence the processing of self-,famous and unknown names and faces:An ERP study.International Journal of Psychophysiology,79
(2),219–230.Tateuchi,T.,Itoh,K.,& Nakada,T.(2012).Neural mechanisms underlying the orienting response to subject's own name: An event-related potential study.Psychophysiology,49
(6),786–791.Taylor,J.G.,&Fragopanagos,N.F.(2005).The interaction of attention and emotion.Neural Networks,18
(4),353–369.Thorpe,S.,Fize,D.,&Marlot,C.(1996).Speed of processing in the human visual system.Nature,381
(6582),520–522.Trafimow,D.,Triandis,H.C.,&Goto,S.G.(1991).Some tests of the distinction between the private self and the collective self.Journal
of
Personality
and
Social Psychology,60
(5),649–655.Turk,D.,Heatherton,T.F.,Kelley,W.M.,Funnell,M.,Gazzaniga,M.S.,&Macrae,C.N.(2002).Mike or me?Self-recognition in a split-brain patient.Nature Neuroscience,5
(9),841–842.Wagar,B.,&Cohen,D.(2003).Culture,memory,and the self:An analysis of the personal and collective self in long-term memory.Journal of Experimental Social Psychology,39
(5),468–475.Watkins,P.C.,Vache,K.,Verney,S.P.,&Mathews,A.(1996).Unconscious mood-congruent memory bias in depression.Journal of Abnormal Psychology,105
(1),34–41.Watson,L.A.,Dritschel,B.,Jentzsch,I.,&Obonsawin,M.(2008). Changes in the relationship between self‐reference and emotional valence as a function of dysphoria.British Journal of Psychology,99
(1),143–152.Watson,L.A.,Dritschel,B.,Obonsawin,M.C.,&Jentzsch,I.(2007).Seeing yourself in a positive light:Brain correlates of the self-positivity bias.Brain Research,1152
,106–110.Yuan,J.,Yang,J.,Meng,X.,Yu,F.,&Li,H.(2008).The valence strength ofnegativestimulimodulatesvisual novelty processing:Electrophysiological evidence from an event-related potential study.Neuroscience,
157
(3),524–531.Yuan,J.,Zhang,Q.,Chen,A.,Li,H.,Wang,Q.,Zhuang,Z.,&Jia,S.(2007).Are we sensitive to valence differences in emotionally negative stimuli? Electrophysiological evidence from an ERP study.Neuropsychologia,45
,2764–2771.Zhang,L.,Zhou,T.,Zhang,J.,Liu,Z.,Fan,J.,&Zhu,Y.(2006).In search of the Chinese self:An fMRI study.Science in China Series C,49
(1),89–96.Zhao,K.,Wu,Q.,Zimmer,H.D.,& Fu,X.(2011).Electrophysiological correlates of visually processing subject's own name.Neuroscience
Letters,
491
(2),143–147.Zhao,K.,Yuan,J.,Zhong,Y.,Peng,Y.,Chen,J.,Zhou,L.,...Ding,D.(2009).Event-related potential correlates of the collective self-relevant effect.Neuroscience Letters,464
(1),57–61.