内隐重复效应影响外显工作记忆的年龄差异*
2014-02-06刘英杰丁锦红郭春彦
刘英杰 魏 萍 丁锦红 郭春彦
(首都师范大学心理系,北京市“学习与认知”重点实验室,北京 100048)
1 引言
工作记忆是对有限容量的信息进行暂时贮存和保持,并对此信息进行操作加工和执行控制的系统。先前经验对记忆测验成绩的影响可以是有意识的,也可以是无意识的,表现为记忆的外显(explicit)或内隐(implicit)类型。在过去几十年里,大量来自实验心理学、神经心理学和认知神经科学的研究关注两种记忆类型的神经基础和相互作用。相对一致的结论认为,外显记忆与内隐记忆依赖于不同的脑内记忆系统(Gazzaniga,1998;Grill-Spector,Henson,&Martin,2006;Grill-Spector&Malach,2001;Jiang,Haxby,Martin,Ungerleider,&Parasuraman,2000;Markowitsch,2008;Meng&Guo,2007a;Paller,Voss,&Boehm,2007;Voss&Paller,2008)。但是,两种记忆系统随年龄变化而发生发展的机制仍不清楚。近期一些研究报告了人类有意识的记忆过程或外显记忆的年龄差异(Fabiani&Gratton,2005;Fleischman&Gabrieli,1998;Reuter-Lorenz,2000),但是对于内隐记忆、内隐与外显记忆的关系如何受到年龄变化的影响的研究较少,得到的结论也不一致。
一些研究发现,重复启动任务存在年龄差异(Cherry&St.Pierre,1998;Fleischman&Gabrieli,1998;La Voie&Light,1994;Lawson,Guo,&Jiang,2007;Liu,Guo,&Jiang,2006),老年被试的重复启动效应减小。重复启动效应指的是,被试对重复出现的刺激加工得更快、更好,对重复刺激的反应时加快、错误率降低,这体现了被试对该刺激的内隐学习和记忆。这种效应在遗忘症病人身上也存在,虽然病人并没有意识到这一点(Gabrieli,1998)。所以,重复启动效应被认为是一种不需要意识觉知的自动学习过程,不同于有意识的记忆或外显记忆。老年被试对三维物体(Jiang,Greenwood,&Parasuraman,1999;Jiang,Luo,&Parasuraman,2002,2009)、情绪性刺激(Jiang,Vagnini,Clark,&Zhang,2007)等的重复启动效应均较年轻被试明显降低,体现了老年人内隐学习能力一定程度的下降。Jiang等人(2009)使用事件相关电位技术(Event-Related Potential,ERP)证明,当启动刺激和目标刺激相隔时间短于400毫秒时,老年被试对目标刺激脑电信号反应强度减弱、时间进程减慢是导致老年被试启动效应降低的主要原因。但是,也有一些研究发现,老年被试只是在整体反应时上慢于年轻被试,但仍存在与年轻被试相当的重复启动效应量(Pfutze,Sommer,&Schweinberger,2002)。
多数这类研究使用了视觉启动范式,在这类范式中,被试只需被动地观察启动刺激而不进行反应,只对随后呈现的目标刺激进行判断。有研究者认为,这类范式可能导致外显记忆对内隐重复启动效应的污染(Mitchell&Bruss,2003),也是导致研究结果不一致的主要原因。采用延迟样本匹配任务(delayed-matching-to-sample task)并操纵被试对实验部分项目的有意识学习经验,则可以直接考察外显的学习经验对重复启动效应的影响(Guo,Lawson,&Jiang,2007;Guo,Lawson,Zhang,&Jiang,2008;Lawson et al.,2007)。被试首先对一些项目或图片进行有意识的学习。在之后的工作记忆任务中,这些学习过的图片和一些新图片作为工作记忆任务的靶或分心物。在一个试次(trial)中,首先呈现样本图片(学习过的或新的),之后呈现一系列需要判断的图片测验,包含靶、学习过的分心物、新的分心物等,这些图片可多次出现。被试需要判断每个测验图片是否是样本图片,即是不是靶刺激。在工作记忆中保持靶、排除分心物的过程中,多次出现的刺激会导致被试更快的加工,出现启动效应。Lawson等人(Lawson et al.,2007)使用该范式并采用事件相关电位技术发现,年轻被试拒绝干扰项的早期启动效应(200~550 ms)在脑电反应上显著受到之前外显学习经验的影响,而老年被试的该效应则不受影响。也就是说,年轻被试在脑电反应上区分出了学习过的分心物和新的分心物。在行为反应上,老年被试对学习过的分心物的反应时短于对新分心物的反应时,但仅限于首次呈现时。随着刺激重复出现,学习过的分心物可能逐渐建立起内隐的熟悉性,导致老年被试对其拒绝困难。而在年轻被试上,则不存在该效应。但是,老年被试在整体的重复启动效应(包括对靶的重复效应和对干扰项的启动效应)上并不比年轻被试小,甚至表现出了更大的启动受益。这说明,单纯考察重复效应不能全面考察老年被试的工作记忆加工特点,老年被试可能更难区分外显学习过的知识和内隐的熟悉性。
其次,虽然有多种方法可以对执行功能(executive function)进行分类,认知老龄化的研究一般将老年被试在工作记忆任务上的困难归结为加工资源不足和对分心物的抑制困难(Miyake,Friedman,Emerson,Witzki,&Howerter,2000)。例如,采用双任务范式,年轻被试在加工资源短缺情况下表现出与老年被试在单任务条件下类似的行为模式(Anderson,Craik,&Naveh-Benjamin,1998)。老年被试对分心物的抑制困难则表现在多种范式中,例如负启动、视觉搜索或Stroop任务(Andrés,Guerrini,Phillips,& Perfect,2008;Connelly&Hasher,1993;Darowski,Helder,Zacks,Hasheer,&Hambrick,2008),难以拒绝分心物可能导致过多的无关信息存储在工作记忆中,从而降低了整体的加工效率。但是到目前为止,还没有研究采用延迟样本匹配任务考察较大工作记忆负荷下,老年被试和青年被试对工作记忆的重复效应如何受到项目先前学习经验的影响。过去研究采用延迟样本匹配任务时均使用一个靶刺激,被试在工作记忆中只要保持该靶刺激并拒绝所有其他的刺激即可,不容易观察到老年被试的加工局限。其次,重复出现的分心物将建立起内隐的熟悉性,这种熟悉性容易跟工作记忆中持续保持的靶刺激产生混淆,老年被试抑制干扰刺激的能力更弱,更容易表现出对分心物(尤其是学习过的分心物)的拒绝困难。基于以上的假设,在当前的研究中,我们采用延迟样本匹配任务,考察老年被试的重复启动效应如何受到外显记忆的影响。在前期研究基础上进行了如下新的操纵。其一,增加了工作记忆中追逐靶的难度,将样本图片的数量从1个增加到2个。随着工作记忆中需要保持的靶刺激增多,提高了工作记忆的负载,老年被试的加工资源可能不足,保持靶和排除分心物更加困难,此时更利于考察老年被试和青年被试在外显工作记忆任务上和内隐启动效应上的差异;其二,考察靶和分心物的重复效应如何受到外显学习经验的影响。在对重复效应的考察上,除考察被试对测验项目第1次呈现和第2次呈现的反应,增加对第3和第4次呈现的反应时考察,被试对反复呈现的分心物建立起内隐的熟悉性,我们预测,由于老年被试无关信息抑制能力下降,可能更容易表现出对分心物的拒绝困难。
2 方法
2.1 被试
20名在校大学生或研究生(14名女生,6名男生,19~26岁,M
=21岁,SD
=1.85)以及20名老年被试(12名女性,8名男性,64~76岁,M
=68岁,SD
=3.43)参加了实验。被试均为右利手,视力或矫正视力正常,无色盲色弱。老年被试中2人为中专学历,1人为大专学历,其余均为本科学历,与青年被试在受教育程度上匹配。所有被试均自愿参加实验,实验后获得一定的报酬。2.2 刺激材料
实验材料选用了Snodgrass和Vanderwart(1980)设计的线条图片,共 144张。图片大小8.3 cm×5.8 cm,视距 65 cm,视角为 7.1°×5.1°。实验前先让被试学习36张图片,在工作记忆任务阶段该36张图片作为熟悉靶图片或熟悉分心物图片。另 108张未学习过的新图片中,36张用作新靶图片,72张用作新分心物图片。旧靶图片和新旧分心物图片在相似性和复杂性上均进行了匹配。
2.3 实验设计和程序
被试坐在照明柔和适度的电磁屏蔽室内完成整个实验程序。实验分为学习阶段和工作记忆任务阶段(实验程序见图1)。在学习阶段,被试学习并记住36张图片,每张图片呈现10 s。之后,被试对学习过的36个图片以及另外36个新图片(这些新图片在工作记忆任务中不再使用)进行再认任务,对旧图片的再认正确率在94%以上,保证这些图片在接下来的工作记忆阶段作为熟悉客体(熟悉靶或者熟悉分心物)。
在工作记忆任务阶段,采用延迟样本匹配任务范式。一个序列(trial)由2个靶和12个测验图片组成。两个靶并排呈现在屏幕中央,靶图片的外框用绿色标记,呈现时间为4000 ms。这两个靶或者都是新靶,或者都是熟悉靶。测验图片有3种:靶、已学习过的熟悉分心物、未学习过的新分心物。每次呈现一张测验图片,每个测验图片呈现时间在700 ms,随后呈现注视加号,时间在1000~1200 ms间随机变化,每种测验图片重复出现3~4次。要求被试对测验图片进行又快又准地按键反应,一半的被试对“靶”用左手食指按键,对“分心物”用右手食指按键,另一半被试相反。8个序列构成一个区组(block),共有9个区组。使用新靶的序列和使用熟悉靶的序列在区组内以“ABBAAB??”的方式平衡。在每个区组之间,被试有2~3 min的休息时间。
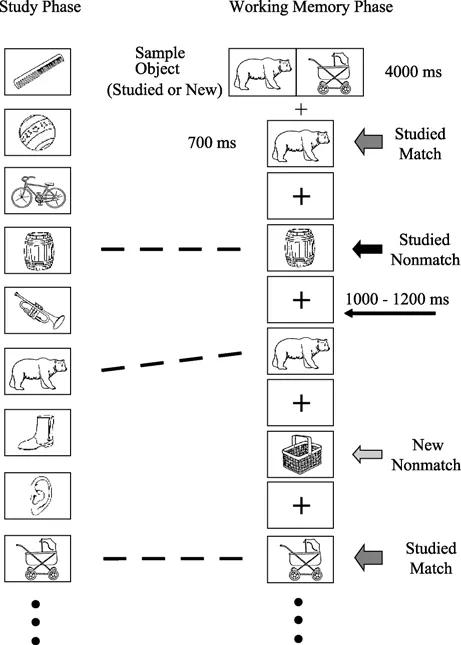
图1 实验流程示例
3 结果
对每个被试按照实验条件分别计算了平均反应时和正确率,熟悉靶、熟悉分心物、新靶、新分心物在第1次呈现、第2次呈现、第3和第4次呈现的平均值见表1和图2。
3.1 反应时
以被试(青年被试 vs.老年被试)为组间变量,以目标属性(靶 vs.分心物)、是否学习过(熟悉的vs.新的)、重复次数(第1次、第2次、第3和第4次的平均值)为组内变量对被试的反应时进行重复测量方差分析。结果发现,被试的主效应显著,F
(1,38)=47.18,p
<0.001,partial η
²=
0.993(关于η²
,partial η
²的区别请见Levine&Hullett,2002), 青年被试的反应时显著快于老年被试的反应时(514 vs.618 ms)。目标属性的主效应显著,F
(1,38)=195.74,p
<0.001,partial η
²=0.837,被试对靶的反应时快于对分心物的反应时(544 vs.588 ms);是否学习过的主效应显著,F
(1,38)=4.05,p
=0.051,partial η
²=0.096,被试对熟悉的图片的反应时快于对新图片的反应时(564 vs.567 ms);重复次数的主效应显著,F
(2,76)=29.54,p
<0.001,partial η
²=0.437,被试对图片第1次出现时的反应时显著慢于对第2次出现的反应时以及第3和第4次的平均值(574 vs.562 vs.561 ms),ps
<0.001。目标属性与是否学习过的交互作用显著,F
(1,38)=104.18,p
<0.001,partial η
²=0.733。事后检验表明,对熟悉靶的反应时快于对新靶的反应时(536 vs.550 ms),p
<0.001,但是,对熟悉分心物的反应时慢于对新分心物的反应时(592 vs.584 ms),p
<0.001。目标属性与重复次数的交互作用显著,F
(2,76)=25.07,p
<0.001,partial η
²=0.397,是否学习过与重复次数的交互作用也显著,F
(2,76)=7.90,p
<0.005,partial η
²=0.172,这说明,被试对熟悉靶、新靶、熟悉分心物、新分心物等的反应时受重复次数的影响有差异。这个差异进一步由三重交互作用证实,F
(2,76)=5.30,p
<0.01,partial η
²=0.122。重要的是, 组间变量与上述因素或因素的二重交互作用没有交互,但是与三因素的交互作用显著,F
(2,76)=3.54,p
<0.05,partial η
²=0.085,说明青年被试和老年被试对熟悉靶、新靶、熟悉分心物、新分心物的重复效应模式上有差异。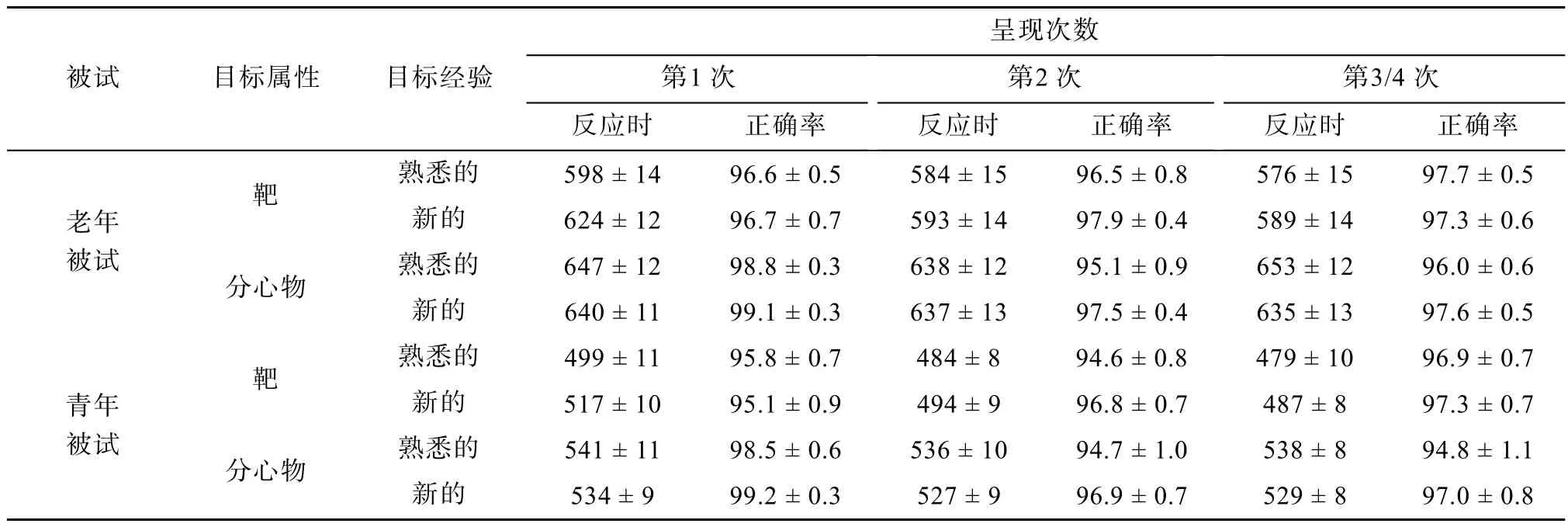
表1 老年被试和青年被试在各个实验条件下的反应时±标准误以及正确率(%)±标准误
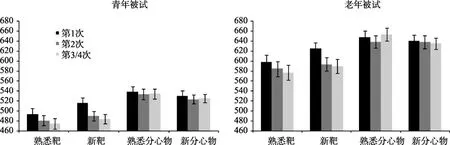
图2 两组被试在不同刺激条件下(熟悉靶、熟悉分心物、新靶、新分心物)的反应时和标准误
为进一步分析青年被试与老年被试在不同条件下重复效应的差异,对青年被试或老年被试在熟悉靶、新靶、熟悉分心物、新分心物的不同呈现次数上反应时进行部分方差分析。以被试为组间变量、重复次数为组内变量,对被试对熟悉靶的反应时进行方差分析显示,重复次数的主效应显著,F
(2,76)=27.00,p
<0.001,partial η
²=0.415。Bonferroni
矫正的事后检验表明,被试对第1次出现的图片的反应时显著快于对第2次的反应时,也快于第3和第4次的平均值(548 vs.534 vs.527 ms),ps
<0.001,被试对图片第2次出现的反应时显著快于对第3和第4次出现的反应时,p
=0.051。组间变量与重复次数没有交互作用,F
(2,76)<1,这说明两组被试对熟悉靶的重复效应的模式相同。对新靶做同样的方差分析,结果显示,重复次数的主效应显著,F
(2,76)=55.64,p
<0.001,partial η
²=0.594, 被试对图片第1次呈现的反应时显著慢于重复出现的反应时(571 vs.543 vs.538 ms),ps
<0.001,对第2次和第3、第4次的反应时没有差异。组间变量与该因素的交互作用也不显著,说明两组被试对新靶的重复效应模式没有差异。对熟悉分心物做同样的方差分析,结果显示,重复次数的主效应显著,F
(2,76)=3.04,p
=0.054,partial η
²=0.074,对图片第1次呈现的反应时与第2次、第3第4次的没有差异,p
>0.1,但是被试对第3第4次呈现的反应时显著慢于对第2次呈现的反应时,p
=0.05。同时,组间变量虽与重复次数的交互作用不显著,F
(2,76)=1.74,p
>0.05,partial η
²=0.044,但是,组间变量与重复次数的组内比较(within-subjects contrasts)交互显著,F
(1,38)=3.28,p
=0.078,partial η
²=0.323, 即两组被试在重复效应上的模式变化有差异。后续分析显示,青年被试在不同呈现次数上的反应时没有差异(541 vs.536 vs.538 ms),F
(2,38)<1;而老年被试在不同呈现次数上的反应时有差异,F
(2,38)=3.55,p
<0.05,Bonferroni
矫正的事后检验表明,老年被试在第1次和第2次呈现的反应时没有差异(647 vs.638 ms),p
>0.1,但是第3第4次呈现的反应时显著慢于对第2次呈现的反应时(653 vs.638 ms),p
<0.05。对新分心物的同样检验发现,重复次数的主效应不显著,F
(2,76)=2.05,p
>0.1,partial η
²=0.051, 组间变量与该因素也没有交互作用,F
(2,76)<1,p
>0.1,partial η
²=0.013,说明两组被试对新分心物均没有重复效应。接着,我们分析了两组被试在重复效应变化趋势上的差异。将重复效应I(项目第1次呈现时被试的反应时减第2次呈现时被试的反应时)和重复效应II(项目第2次呈现时被试的反应时减第3、4次呈现时的反应时)进行以被试(青年被试 vs.老年被试)为组间变量,以目标属性(靶 vs.分心物)、是否学习过(熟悉的 vs.新的)、重复效应次数(重复效应I vs.重复效应II)为组内变量的重复测量方差分析。结果显示,目标属性的主效应显著,F
(1,38)=29.16,p
<0.001,partial η
²=0.434, 被试对靶的重复效应显著大于对分心物的重复效应(13 vs.1 ms);是否学习过的主效应显著,F
(1,38)=16.90,p
<0.001,partial η²
=0.308,被试对熟悉的图片的重复效应小于对新图片的重复效应(5 vs.9 ms);重复效应次数的主效应显著,F
(1,38)=26.80,p
<0.001,partial η
²=0.414,重复效应I显著大于重复效应II(13 vs.1 ms)。上述因素的两两交互作用均不显著,ps
>0.1,同时,组间变量与上述单个因素或两因素交互作用的交互作用不显著。但是,目标属性、项目是否学习过、重复效应次数的三重交互作用显著,F
(1,38)=15.77,p
<0.001,partial η
²=0.293。更重要的是,组间变量与该三重交互作用的交互作用显著,F
(1,38)=4.57,p
<0.05,partial η
²=0.107。这说明,是否学习过对靶或分心物两次重复效应的影响在两组被试间存在差异。为进一步分析两组被试对靶或分心物的重复效应(见图3)的差异如何受到项目先前学习经验的差异性影响,对青年被试或老年被试在熟悉靶、新靶、熟悉分心物、新分心物上的两次重复效应变化进行了部分方差分析。以被试为组间变量、重复效应次数为组内变量,对两组被试对熟悉靶的重复效应进行方差分析显示,重复效应次数的主效应边缘显著,F
(1,38)=3.55,p
=0.06,partial η
²=0.085,被试的重复效应I显著大于重复效应II(14 vs.6 ms)。但是,组间变量与该因素的交互作用不显著,F
(1,38)<1,即两种被试对熟悉靶的重复效应变化趋势相同。对新靶的相同分析显示,重复效应次数的主效应显著,F
(1,38)=49.53,p
<0.001,partial η
²=0.566,被试的重复效应I显著大于重复效应II(27 vs.4 ms)。类似地,组间变量与该因素的交互作用也不显著,F
(1,38)=1.81,p
>0.1,partial η
²=0.045,即两种被试对新靶的重复效应变化趋势相同。对熟悉分析物做同样的方差分析,结果显示,重复效应次数的主效应显著,F
(1,38)=11.86,p
<0.005,partial η
²=0.238,被试的重复效应I显著大于重复效应II(7 vs.-9 ms)。同时,组间变量与重复效应次数的交互作用边缘显著,F
(1,38)=2.92,p
=0.09,partial η
²=0.071,说明两组被试对熟悉分心物的重复效应变化趋势有差异。进一步的独立样本T检验显示,老年被试和青年被试在熟悉分心物的重复效应I上没有差异(9 vs.5 ms),t
(38)<1,但是,老年被试对熟悉分心物的重复效应II显著小于青年被试的该效应(-15 vs.-3 ms),t
(38)=-1.8,p
=0.08。对新分心物做同样的方差分析,结果显示,重复效应次数的主效应不显著,F
(1,38)<1,重复效应I和重复效应II的量没有差异(4 vs.2 ms);此外,组间变量与重复效应次数的交互作用不显著,F
(1,38)<1,两组被试在两次重复效应上的模式没有差异。3.2 正确率
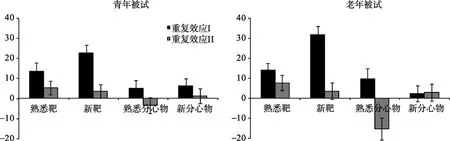
图3 两组被试在不同刺激条件下(熟悉靶、熟悉分心物、新靶、新分心物)的重复效应和标准误
以被试(青年被试 vs.老年被试)为组间变量,以目标属性(靶 vs.分心物)、是否学习过(熟悉的vs.新的)、重复次数(第1次、第2次、第3和第4次的平均值)为组内变量对被试的正确率进行重复测量方差分析。结果显示,组间变量的主效应显著,F
(1,38)=1.34,p
>0.1,partial η
²=0.034, 老年被试与青年被试的正确率没有差异(97.2%vs.96.5%);是否学习过的主效应显著,F
(1,38)=28.16,p
<0.001,partial η
²=0.426,被试对熟悉图片的正确率低于对新图片的正确率(96.4%vs.97.4%);重复次数的主效应显著,F
(2,76)=9.74,p
<0.001,partial η
²=0.204。Bonferroni
矫正的事后检验表明,被试对第1次出现的图片的正确率显著高于第2次呈现的正确率(97.5%vs.96.3%),p
<0.001,第2次呈现与第3第4次呈现的正确率没有差异(96.3%vs.96.8%),p
>0.1。目标属性与是否学习过的交互作用显著,F
(1,38)=5.95,p
<0.05,partial η
²=0.135。后续检验表明,对熟悉靶和新靶的正确率没有差异(96.4%vs.96.8%),p
>0.1,但是对新分心物的正确率显著高于对熟悉分心物的正确率(97.9%vs.96.3%),p
<0.001。目标属性与重复次数的交互作用显著,F
(2,76)=26.94,p
<0.001,对目标的正确率随重复逐渐上升(96.0%vs.96.4%vs.97.3%),p
<0.01,而对分心物的正确率随重复逐渐下降(98.9%vs.96.1%vs.96.4%),p
<0.001。是否学习过与重复次数的交互作用也显著,F
(2,76)=12.04,p
<0.001,对熟悉项目的正确率先降再升(97.4%vs.95.2%vs.96.4%),p
=0.005,而对新项目的正确率保持不变(97.5%vs.97.3%vs.97.3%),p
>0.1。以上结果说明正确率随重复次数的变化在靶与分心物之间、以及在熟悉图片和新图片间存在差异。但是三重交互作用不显著,F
(2,76)=1.50,p
>0.1,partial η
²=0.038。组间因素与上述任何因素或因素的交互作用均不显著,这说明两组被试正确率的模式没有差异。4 讨论
采用延迟样本匹配任务并控制被试对部分项目的有意识学习经验,当前研究考察了老年被试与青年被试在追逐靶、排除分心物的过程中,重复启动效应如何受到项目外显学习经验的影响。首先,老年被试与青年被试追逐靶和排除分心物的过程均受到项目之前学习经验的影响。无论是老年被试还是青年被试,对靶的反应时均快于对分心物的反应时,对外显学习过的靶(即熟悉靶)的反应时快于对未学习过的靶(即新靶)的反应时,而拒绝熟悉的分心物需要的时间长于拒绝新分心物的时间。其次,老年被试与青年被试均表现出对靶的重复效应,即当靶(无论是熟悉的还是新的)在任务中重复出现时,对其的反应时加快;对分心物的重复效应小于对靶的重复效应。随着项目多次重复,重复效应整体上减小,但该效应的变化受到项目属性(靶或分心物)以及项目之前学习经验的影响。重要的是,当熟悉的分心物多次出现时,老年被试不但没有出现重复效应,反而在拒绝该熟悉分心物上表现出困难,反应时显著延长,而在青年被试上没有该表现。
该研究结果首先说明,工作记忆任务中追逐靶和排除分心物的效率存在差异。无论是老年被试或青年被试,对靶的反应时快于对分心物的反应时。随着靶或分心物多次呈现,反应内隐学习与记忆的重复效应在靶和分心物之间也存在差异。对靶来讲,无论是熟悉靶还是新靶,无论是老年被试还是青年被试,均表现出重复效应,即对多次出现的靶刺激反应时加快。但是,分心物的重复效应显著减小甚至无重复效应。而在反应正确率上,对多次重复出现的靶的正确率提高,但是对多次重复出现的分心物的正确率下降。这些结果与前人的相关研究一致。例如,Caggiano,Jiang和Parasuraman(2006)采用单靶延迟样本匹配任务(但是没有控制项目的前期学习经验),让被试判断测试项目是否是靶刺激。靶和分心物可能重复多次出现(最多5次)。结果发现,随着项目多次出现,被试对靶的反应时逐渐加快,而对分心物的正确率逐渐下降。对靶的反应加快反应了内隐熟悉性造成的促进效应,而重复出现引起的熟悉性则干扰了拒绝分心物的过程。与过去研究有所不同的是,Caggiano等人(2006)以及Guo等人(2007,2008)均发现被试对靶的反应时慢于对分心物的反应时,而当前研究中两组被试均对靶的反应时快于对分心物的反应时。这可能与实验中使用的样本图片数量有关。上述提到的前人研究均使用单样本刺激,被试在进行匹配任务时可能采用排除法,只要测试图片与样本刺激有不同,即可快速判定为分心物,而当遇到靶图片时则需要与样本图片进行匹配。当前研究采用双样本图片,对分心物的拒绝要完成与两个样本的对比才能拒绝,而对靶图片的确认只要与其中一个样本匹配即可。这个研究结果可能与视觉搜索过程中的双目标搜索过程类似(Menneer et al.,2012),当需要在视觉工作记忆中保持多个目标时,对分心物的排除效率显著低于只在工作记忆中保持一个项目时。
其次,工作记忆任务中追逐靶和排除分心物的效率受到了项目之前学习经验的影响。一方面,在确认靶的过程中,无论是老年被试还是青年被试,对熟悉靶的反应时均快于对新靶的反应时。另一方面,在排除分心物的过程中,老年被试与青年被试均对熟悉分心物比对新分心物反应更慢。学习过的客体图片具有较高的熟悉性,作为靶时更容易被确认,而作为分心物时则更容易引起混淆,造成对其的反应时减慢。过去研究发现,老年被试对面孔(Bartlett&Fulton,1991;Bastin&van der Linden,2003)或文字的记忆(Balota,Burgess,Cortese,&Adams,2002;Jennings&Jacoby,1997)更依赖于熟悉性(即该项目是不是刚刚出现过),而在特定的回忆任务中容易出现行为缺陷。而青年被试,虽然整体的反应时快于老年被试,但是在目标属性(靶 vs.分心物)与项目学习经验的交互作用模式上与老年被试相似。这暗示,这种外显学习经验导致的熟悉性对老年被试和青年被试的影响是存在相似性的。
再次,当前研究发现,对靶或分心物的内隐重复启动效应也受到项目之前外显学习经验的影响。两组被试对靶,无论是新靶还是熟悉靶,均产生显著的重复效应,并且重复效应II要小于重复效应I,即项目第2次呈现时相对于首次呈现时的重复效应大于第3、4次相对于第2次的重复效应。而当分心物多次呈现时,青年被试在熟悉分心物和新分心物上均没有出现显著的重复效应,老年被试对熟悉分心物非但没有重复效应,反而在其多次出现时反应时显著变慢。当前研究首次报告了老年被试对熟悉分心物的拒绝困难。Lawson等人(2007)采用了类似的实验范式,但是被试只需在工作记忆任务中追逐一个靶刺激,结果并没有发现老年被试对重复出现的熟悉分心物的拒绝困难,老年被试在熟悉的分心物第2次出现和第3次出现的反应时相当,并显著快于第1次呈现的反应时,重复效应显著。而在当前研究中,被试需要在工作记忆中保持两个靶刺激,熟悉的分心物在第2次呈现时的反应时即与第1次的反应时相当,没有重复效应;而当熟悉的分心物多次呈现时,老年被试对其的拒绝更加困难。当前结果与Lawson等人(2007)结果说明两个问题。一方面,青年被试在工作记忆任务中对靶的保持优于老年被试。当工作记忆的负荷较低时,老年被试对靶的保持不存在困难,此时拒绝分心物只需与靶进行一次对比即可完成。即使分心物曾经学习过,具有较高的熟悉性,但是对靶的清晰保持保证了对分心物的快速拒绝。但是,当工作记忆的负荷变高时,对靶的保持难度增加,拒绝分心物需要完成与两个靶的对比才能正确拒绝。此时学习过的分心物因具有较高的熟悉性而干扰了老年被试,尤其是在该分心物多次出现时。另一方面,重复启动效应受到工作记忆负荷的调控。当工作记忆的负荷低时,Lawson等人(2007)发现了对熟悉分心物的重复效应;而当前研究的工作记忆负荷较高,老年被试和青年被试对熟悉分心物的重复效应均消失。但是,目前还没有研究在延迟样本匹配任务中直接操纵和比较不同记忆负荷条件下重复效应如何受到项目外显学习经验的影响。孟迎芳和郭春彦(2007b)采用学习-再认范式,报告了学习阶段的加工负荷对再认阶段内隐记忆效应的影响,只有在学习阶段处于低负荷的面孔刺激才在再认阶段的事件相关电位上表现出了内隐记忆效应,说明内隐记忆效应确实可以受到编码阶段加工负荷的调控。今后研究可直接控制再认阶段的工作记忆负荷,来进一步探讨内隐记忆与外显工作记忆任务负荷的关系。
增加记忆负荷显著影响老年被试的工作记忆表现的根本原因是,工作记忆的容量随年龄显著下降(Myerson,Emery,White,&Hale,2003;Chen,Hale,&Myerson,2003)。但是,这种下降的具体机制还需要进一步的证据。一种可能是老年被试对靶的保持出现困难,另一种可能则是老年被试更难抑制分心物的干扰。采用事件相关电位技术,Gazzaley等人(2008)发现,老年被试在工作记忆的编码(encoding)阶段难以抑制任务无关信息;而Vogel等人(Vogel,McCollough,&Machizawa,2005)则发现,低工作记忆容量的被试(老年被试)比高工作记忆容量的被试(青年被试)在延迟阶段(delay period)表征了更多的任务无关信息。虽然抑制任务无关信息的关键阶段的早晚还存在争议,这些研究和当前研究均说明,排除分心物的效率可能是决定个体工作记忆容量和在工作记忆任务中的行为表现的关键因素(Gazzaley et al.,2008;Jost,Bryck,Vogel,&Mayr,2011;Vogel et al.,2005)。但是,需要注意的是,当前研究与上述研究采用了不同的范式。Vogel等人(Vogel et al.,2005;Jost et al.,2011)的研究采用了变化探测任务(change detection task),被试在编码阶段需要记住红色项目的朝向,并忽略绿色或蓝色项目的朝向,在之后的测验阶段判断红色项目的朝向是否发生了变化。ERP结果显示,在项目被编码后的延迟阶段,青年被试能够更好更快地集中在任务相关的红色项目上,而老年被试则对任务无关的绿色或蓝色项目也进行了表征,对分心物的抑制晚于青年被试。而在当前研究中,被试的任务是保持靶刺激并拒绝分心物刺激,对靶或非靶进行辨别判断。此时的分心物也可以说是任务相关的。换句话说,正确地拒绝分心物也是成功保持靶刺激的一种表现。此时,分心物,尤其是重复出现的熟悉分心物在老年被试与青年被试脑内的反应可能存在差异,值得采用脑电技术和功能磁共振成像技术进行进一步的研究。老年被试可能需要较多的前额叶活动来补偿抑制分心物能力下降带来的干扰(近期关于认知老龄化与前额叶功能的综述请见Cabeza&Dennis,2012)。另一方面,在当前的研究范式中,青年被试对多次出现的熟悉分心物虽没有反应减慢,但是也没有显著的重复效应,这说明青年被试在拒绝多次出现的熟悉分心物上也并没有显示出来自内隐学习的加工优势。在正确率的模式上,老年被试和青年被试均对多次重复出现的靶的正确率提高,但是对多次重复出现的分心物的正确率下降。这说明青年被试对重复出现的分心物的加工效果也逐渐变差。未来研究可采用ERP技术来考察青年被试和老年被试在拒绝熟悉的分心物过程中的脑电差异只是量的不同,还是具有时程早晚和头皮分布等的差异。
此外,当前研究表明,老年被试虽然在拒绝熟悉分心物时效率下降,但是正确率并不低。整体来讲,虽未达到显著,老年被试的正确率略高于青年被试(97.2%vs.96.5%)。同时,老年被试与青年被试在整体的正确率模式上相同。这可能说明,老年被试在反应策略上更慎重、更保守,努力保证能够做到更高的正确率。此时,在遇到重复出现的熟悉分心物上表现出的迟疑确实体现了老年被试对该项目的加工困难。当然,当前研究采用的老年被试虽年纪较大,但认知功能仍属正常范围。将来研究可考虑采用轻度认知障碍(Mild Cognitive Impairment,MCI)的老年被试或老年痴呆症(Alzheimer’s Disease,AD)患者来进一步检验其拒绝重复出现的分心物如何受到项目先前学习经验的影响。由于认知功能的损害和工作记忆容量的进一步降低,这些人群则可能在反应时和正确率上均表现出困难。
Anderson,N.D.,Craik,F.I.M.,&Naveh-Benjamin,M.(1998).The attentional demands of encoding and retrieval in youngerand olderadults:Evidence from divided attention costs.Psychology and Aging,13
,405−423.Andrés,P.,Guerrini,C.,Phillips,L.H.,&Perfect,T.J.(2008).Differential effects of aging on executive and automatic inhibition.Developmental Neuropsychology,33
,101−123.Balota,D.A.,Burgess,G.C.,Cortese,M.J.,&Adams,D.R.(2002).The word-frequency mirror effect in young,old,and early-stage Alzheimer’s disease:Evidence for two processes in episodic recognition performance.Journal of Memory and Language,46
,199−226.Bartlett,J.C.,& Fulton,A.(1991).Familiarity and recognition of faces in old age.Memory and Cognition,19
,229−238.Bastin,C.,&van der Linden,M.(2003).The contribution of recollection and familiarity to recognition memory:A study of the effects of test format and aging.Neuropsychologia,17
,14−24.Cabeza,R.&Dennis,N.A.(2012).Frontal lobes and aging:Deterioration and compensation.In D.T.Stuss&R.T.Knight(Eds.),Principles of Frontal Lobe Function
(2nd ed.).Oxford University Press,New York.Caggiano,D.M.,Jiang,Y.,&Parasuraman,R.(2006).Aging and repetition priming for targets and distracters in a working memory task.Aging,Neuropsychology,and Cognition,13
,552−573.Chen,J.,Hale.S.,&Myerson,J.(2003).Effects of domain,retention interval,and information load on young and older adults’ visuospatial working memory.Aging,
Neuropsychology,
and
Cognition
(Neuropsychology,Development and Cognition:Section B),10
,122−133.Cherry,K.E.,&St.Pierre,C.(1998).Age-related differences in pictorial implicit memory:Role of perceptualand conceptual processes.Experimental Aging Research,24
,53−62.Connelly,S.L.,&Hasher,L.(1993).Aging and the inhibition of spatial location.Journal of Experimental Psychology:Human Perception and Performance,19
,1238−1250.Darowski,E.S.,Helder,E.,Zacks,R.T.,Hasher,L.,&Hambrick,D.Z.(2008).Age-related differences in cognition:The role of distraction control.Neuropsychology,22
,638−644.Fabiani,M.,&Gratton,G.(2005).Electrophysiological and optical measures of cognitive aging.In R.Cabeza,L.Nyberg,&D.Park(Eds.),Cognitive Neuroscience of Aging:Linking Cognitive&Cerebral Aging
(pp.85−106).New York:Oxford University Press.Fleischman,D.A.,&Gabrieli,J.D.E.(1998).Repetetion priming in normal aging and Alzheimer’s disease:A review offindingsand theories.Pshochology
&
Aging,13
,88−119.Gabrieli,J.D.E.(1998).Cognitive neuroscience of human memory.Annual Review of Psychology,49
,87–115.Gazzaley,A.,Clapp,W.,Kelley,J.,McEvoy,K.,Knight,R.T.,& D’Esposito, M. (2008). Age-related top-down suppression de fi cit in the early stages of cortical visual memory processing.Proceedings of the National Academy ofSciences
ofthe
United
States
ofAmerica,105
,13122–13126.Gazzaniga,M.S.(1998).Cognitive
Neuroscience
(pp.469–534).Shanghai,China:Shanghai Education Press.[Gazzaniga,M.S.(1998).认知神经科学
(pp.469–534).上海:上海教育出版社.]Grill-Spector,K.,Henson,R.,&Martin,A.(2006).Repetition and the brain:Neural models of stimulus-specific effects.Trends in Cognitive Science,10
(1),14−23.Grill-Spector,K.,&Malach,R.(2001).fMR-adaptation:A toolforstudying the functionalpropertiesofhuman cortical neurons.Acta Psychology,107
,293−321.Guo,C.Y.,Lawson,A.,&Jiang,Y.(2007).Distinct neural mechanisms for repetition effects of visualobjects.Neuroscience,149
,747−759.Guo,C.Y.,Lawson,A.,Zhang,Q.,&Jiang,Y.(2008).Brain potentials of new and studied objects during working memory.Human Brain Mapping,29
(4),441−452.Jennings,J.M.,& Jacoby,L.L.(1997).An opposition procedure for detecting age-related deficits in recollection:Telling effects of repetition.Psychology and Aging,12
,352−361.Jiang,Y.,Greenwood,P.M.,&Parasuraman,R.(1999).Age-related reduction in 3-D visualmotion priming.Psychology and Aging,14
(4),619−626.Jiang,Y.,Haxby,J.V.,Martin,A.,Ungerleider,L.G.,&Parasuraman, R. (2000). Complementary neural mechanisms for tracking items in human working memory.Science,287
,643−646.Jiang, Y., Luo, Y. J., & Parasuraman, R. (2002).Two-dimensional visual motion priming is reduced in older adults.Neuropsychology,16
(2),140−145.Jiang,Y,Luo,Y.J.,&Parasuraman,R.(2009).Neural correlates of age-related reduction in visual motion priming.Aging,Neuropsychology,and Cognition,16
(2),164−182.Jiang,Y.,Vagnini,V.,Clark,J.,&Zhang,Q.(2007).Reduced sensitivity of older adults to affective mismatches.The Scientific World Journal,7
,641−648.Jost,K.,Bryck,R.L.,Vogel,E.K.,&Mayr,U.(2011).Are old adults just like low working memory young adults?Filtering efficiency and age differences in visual working memory.Cerebral Cortex,21
,1147−1154.La Voie,D.,&Light,L.L.(1994).Adult age differences in repetition priming:A meta-analysis.Psychology&Aging,9
,539−553.Lawson,A.L.,Guo,C.,&Jiang,Y.(2007).Age effects on brain activity during repetition priming of targets and distracters.Neuropsychologia,45
,1223−1231.Levine,T.R.,&Hullett,C.R.(2002).Eta squared,partial eta squared and the misreporting of effect size in communication research.Human Communication Research,28
,612−625.Liu,R.,Guo,C.Y.,&Jiang,Y.(2006).An event-related potential study of working memory in children.Chinese Science Bulletin,51
(12),1467−1475.Markowitsch,H.J.(2008).Autobiographical memory:A biocultural relais between subject and environment.European
Archives
of
Psychiatry
and
Clinical Neuroscience,258
,98−103.Meng,Y.F.,&Guo,C.Y.(2007a).ERP dissociation and connection between implicit and explicit memory.Chinese Science Bulletin,52
(21),2945−2953.Meng,Y.F.,&Guo,C.Y.(2007b).The asymmetric effect of interference at encoding or retrieval on implicit and explicit memory.Acta Psychologica Sinica,39
(4),579−588.[孟迎芳,郭春彦.(2007b).编码与提取干扰对内隐和外显记忆的非对称性影响.心理学报,39
(4),579−588.]Menneer,T.,Stroud,M.J.,Cave,K.R.,Li,X.,Godwin,H.J.,Liversedge,S.P.,&Donnelly,N.(2012).Search for two categories of target produces fewer fixations to target-color items.Journal of Experimental Psychology:Applied,18
,404−418.Mitchell,D.B.,&Bruss,P.J.(2003).Age differences in implicit memory: Conceptual, perceptual, or methodological?Psychology&Aging,18
,807−822.Miyake,A.,Friedman,N.P.,Emerson,M.J.,Witzki,A.H.,&Howerter,A.(2000).The unity and diversity of executive functions and their contributions to complex"frontal lobe"tasks:A latent variable analysis.Cognitive Psychology,41
,49−100.Myerson,J.,Emery,L.,White,D.A.,&Hale,S.(2003).Effectsofage,domain,and processing demandson memory span:evidence for differential decline.Aging,Neuropsychology,
and
Cognition
(Neuropsychology,Development and Cognition:Section B),10
,20−27.Paller,K.A.,Voss,J.L.,&Boehm,S.G.(2007).Validating neuralcorrelates offamiliarity.Trends
in
Cognitive Sciences,11
(6),243−250.Pfutze,E.M.,Sommer,W.,&Schweinberger,S.R.(2002).Age-related slowing in face and name recognition:Evidence from event-related brain potentials.Psychology&Aging,17
,140−160.Reuter-Lorenz,P.A.(2000).Cognitive neuropsychology of the aging brain.In D.C.Park,&N.Schwarz(Eds.),Cognitive aging:A primer
(pp.93−114).New York:Psychology Press.Snodgrass,J.G.,&Vanderwart,M.(1980).A standardized set of260 pictures:Normsfornameagreement,image agreement,familiarity,and visual complexity.Journal of Experimental Psychology:Human Learning and Memory,6
,174−215.Vogel,E.K.,McCollough,A.W.,&Machizawa,M.G.(2005).Neural measures reveal individual differences in controlling access to working memory.Nature,438
,500−503.Voss,J.L.,&Paller,K.A.(2008).Neural substrates of Remembering−Electroencephalographic studies. In H.Eich-enbaum(Ed.),Memory Systems
(pp.79−98).Oxford:Elsevier Press.