杜氏盐藻鞭毛内运输蛋白88在鞭毛组装过程中的功能分析及其原核表达*
2013-12-17毛丽红李庆华龚方华蒋海丽薛乐勋1
韩 康,石 科,毛丽红,李庆华,龚方华,蒋海丽,薛乐勋1,
1)郑州大学生物工程系细胞生物学研究室 郑州450001 2)郑州大学第一附属医院细胞生物学研究室 郑州450052#通讯作者,男,1944年2月生,本科,教授,研究方向:肿瘤标志物与基因工程,E-mail:xuelx@zzu.edu.cn
鞭/纤毛内运输(intraflagellar transport,IFT)是存在于纤毛内部的一种运输机制,很多纤毛相关蛋白在纤毛内是借助这种机制而实现定向运动,这种运输主要依靠IFT 复合体,这些复合体能够通过马达蛋白和轴丝相连,并负责纤毛内部的多种成分的结合,从而达到运输的目的[1]。IFT 复合体包含20种以上IFT 蛋白,分别为IFT-A 和IFT-B。IFT 是一种双向运输机制,包括基部到顶端的正向运输和顶端到基部的逆向运输,其中,IFT-B 主要负责正向运输,而IFT-A 与货物的逆向运输相关[2],正是这种可控的双向运输机制,调控着纤毛有序的组装、维持和解聚过程。IFT88 是IFT-B 的成分之一,它是由IFT88 基因所编码的,这种基因能编码三四氨基酸重复序列(tetratricopeptide repeat,TPR)家族,此重复序列能介导蛋白质之间相互作用[3],对于IFT88 蛋白的运输功能起着极其重要的作用。在衣藻细胞内能够与IFT52、IFT46、HSP40 相互作用;在小鼠中,IFT88 同源蛋白Tg737 的突变会引起多囊肾疾病[4],且Tg737 的突变与小鼠的视觉、嗅觉的功能障碍也有一定关系[5-6]。杜氏盐藻是一种高度耐盐的单细胞且具有双鞭毛的真核生物,有一对约13 μm的等长鞭毛,生长周期短,培养方式简单,可作为研究鞭毛的模式生物。作者利用郑州大学生物工程系细胞生物学研究室盐藻转录组的已知片段,利用3'RACE 及5'扩增得到IFT88 基因序列,采用实时荧光定量PCR 方法观察IFT88 mRNA 在杜氏盐藻鞭毛再生过程中的表达情况。预测并扩增其开放阅读框后,成功构建其原核表达载体,为进一步的功能分析提供理论依据。
1 材料与方法
1.1 实验材料 杜氏盐藻UTEX LB-1644 购自美国德克萨斯州大学,大肠杆菌DH5α 为郑州大学生物工程系细胞生物学研究室保存菌种,载体pMD19-T、EcoR I、Hind Ⅲ等DNA 限制性内切酶及IPTG 均购自大连宝生物工程有限公司;PCR 特异性引物由北京华大基因生物技术有限公司合成;胶回收试剂盒、质粒DNA 小量制备试剂盒购自Axygen 公司,Real Master Mix(SYBR Green)购自天根生化科技(北京)有限公司;HPG400 型光照培养箱购自哈尔滨市东联电子技术开发有限公司,紫外分光光度计为Thermo 公司产品,7300 实时荧光定量PCR 仪购自System Applied Biosystems 公司。
1.2 杜氏盐藻IFT88 cDNA 3' 端序列扩增及拼接根据郑州大学生物工程系细胞生物学研究室转录组测序片段,用Primer Premier 5.0 设计2 个3'RACE上游引物:5'-TTGCGGTTGCTGGAGGTG-3'(18 bp)、5'-CAAGCGTCAGAATTACATCA-3'(20 bp)。下游引物为3'RACE 试剂盒中自带引物:3'RACE 内侧引物5'-CGCGGATCCTCCACTAGTGATTTCACTATAGG-3'(32 bp)和3'RACE 外侧引物5'-TACCGTCGTTC CACTAGTGATTT-3'(23 bp)。提取对数生长期杜氏盐藻的总RNA,按照First Choice RLM-RACE Kit 说明进行操作。PCR 反应体系:cDNA 1 μL,上、下游引物(50 μmol/L)各1 μL,dNTP(10 mmol/L)8 μL,10×LA Buffer 10 μL,dH20 补至100 μL。巢式PCR反应产物经10 g/L 的琼脂糖凝胶电泳,胶回收目的片段,与pMD19-T 连接,过夜并转化至大肠杆菌DH5α,用含有氨苄青霉素抗性、IPTG 和X-Gal 的平板进行蓝白斑筛选,挑取阳性菌落培养,提取质粒后经双酶切鉴定,随后将菌液送公司测序,将测序结果与原序列进行拼接。
1.3 杜氏盐藻IFT88 cDNA 5' 端序列扩增及拼接根据拼接的3'端序列,设计5'端下游引物序列,外侧引物5'-ATGCCAGCCCAATGTTCC-3'(18 bp),内侧引物5'-GTTTCGCACGAGGTCTGT-3'(18 bp)。上游引物消减杂交文库特有引物:5'-AAGCAGTGG TATCAACGCAGAGTACXXXXX-3'(X = SMARTer 机密的寡核苷酸序列)(30 bp),巢式PCR 扩增IFT88 cDNA 的5'端序列,PCR 产物用10 g/L 琼脂糖凝胶检测,回收后与pMD19-T 载体连接并转化至大肠杆菌DH5α,阳性克隆扩大培养。提质粒,EcoR Ⅰ、Hind Ⅲ双酶切鉴定,送公司测序后,将测序结果与已知序列进行拼接。
1.4 实时荧光定量PCR 使用机械研磨法去除杜氏盐藻鞭毛[7],在去除鞭毛后的180 min 内每30 min 取样1 次,用Trizol 法提取总RNA,逆转录为cDNA 作为模版,设计一对引物5'-ACAGACCTCGT GCGAAAC-3' (18 bp)、5'-ATGCCAGCCCAATGT TCC-3'(18 bp),以GAPDH 作为内参,进行实时荧光定量PCR,随后导出数据进行分析。
1.5 验证开放阅读框及其原核表达 将拼接序列输入ORF finder 预测其开放阅读框,设计原核表达上游引物:5'-CCGGAATTCATGTCGCGCAATGCTGACGA-3'(29 bp);原核表达下游引物:5'-CCCAAGCTTTTATC CCATTGGTAGCAAGT-3'(29 bp)。以制备的cDNA 为模板,利用合成的引物采用PCR 法扩增盐藻细胞预测的IFT88 开放阅读框的基因序列。PCR 反应程序:95℃5 min;94℃30 s,40~60℃30 s,72℃150 s,30 个循环;72℃10 min,4℃终止反应。PCR反应结束后,用10 g/L 琼脂糖凝胶电泳检测扩增结果。将PCR 产物胶回收,连接于pMD19-T 载体上,转化至大肠杆菌DH5α,挑取阳性菌落培养,双酶切鉴定后送公司测序。将测序正确的菌液质粒回收,EcoR I、Hind Ⅲ双酶切后,产物用10 g/L 琼脂糖凝胶检测并回收,随后与pET28a 载体连接,转化至大肠杆菌BL21(DE3)中,进行原核表达。并进一步利用1 mmol/L IPTG,37℃诱导4 h,进行聚丙烯酰胺凝胶电泳(SDS-PAGE),观察表达情况。
2 结果
2.1 杜氏盐藻IFT88 基因cDNA 3'RACE 结果3'RACE 所得片段胶回收后连接于pMD19-T 得到质粒pMD-I1,经EcoR I、Hind Ⅲ双酶切鉴定后(图1),送公司测序,测序长度约为2 275 bp。

图1 IFT88 3'RACE 及pMD-I1 的酶切鉴定结果
2.2 杜氏盐藻IFT88 基因cDNA 5'RACE 结果经5'扩增后得到一条约1 000 bp 的条带,胶回收后与pMD19-T 载体连接得到质粒pMD-I2,双酶切鉴定结果见图2,送公司测序后得到998 bp 核苷酸片段。
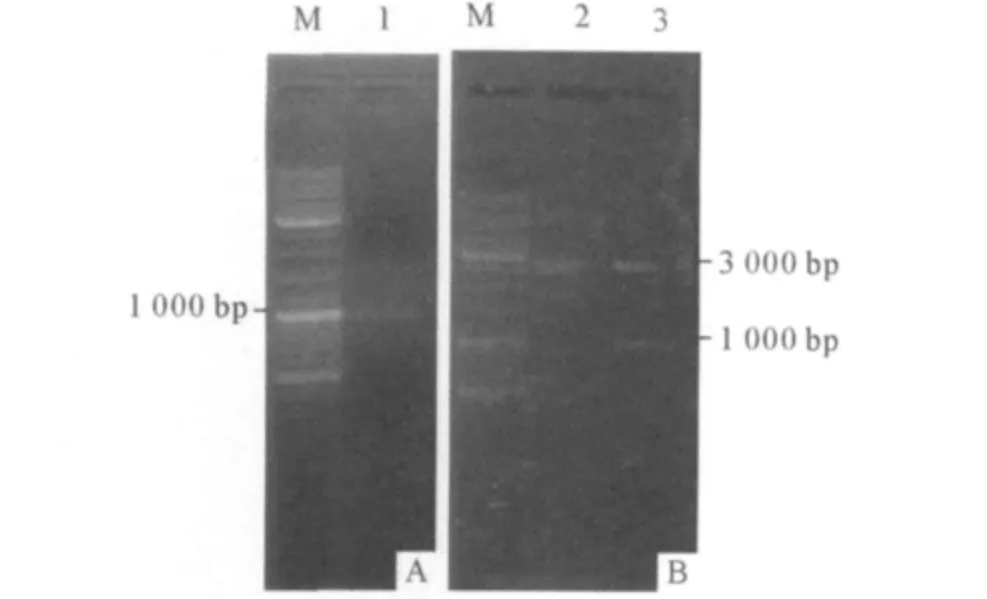
图2 IFT88 5'端扩增及pMD-I2 的酶切鉴定结果
2.3 实时荧光定量PCR 结果 结果见图3。与未去除鞭毛组比较,在去除鞭毛后的180 min 内,IFT88 基因的转录量在30 min 时达到了对照组的4倍,随后转录水平迅速降低,与盐藻鞭毛生长速率曲线基本一致。
2.4 pET28a-IFT88 载体构建及其原核表达结果扩增得到开放阅读框约2 400 bp(图4A),公司测序结果显示序列无误,编码799 个氨基酸,BLAST 显示该氨基酸序列与多个物种的IFT88 有较高同源性。胶回收片段,与pET28a 载体连接后,转化至大肠杆菌BL21(DE3)中,与pET28a 载体连接后得到质粒pET28a-IFT88,质粒双酶切鉴定结果见图4B。SDSPAGE 显示37℃、1 mmol/L IPTG 诱导4 h 后在约90 000处有明显高表达(图5)。

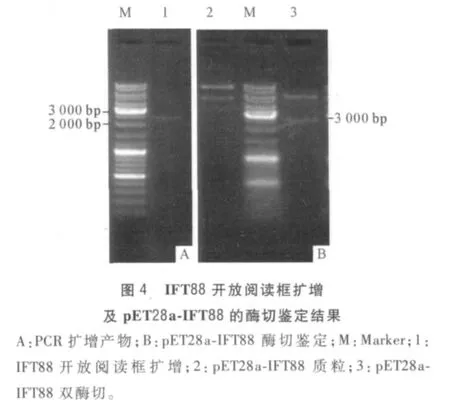
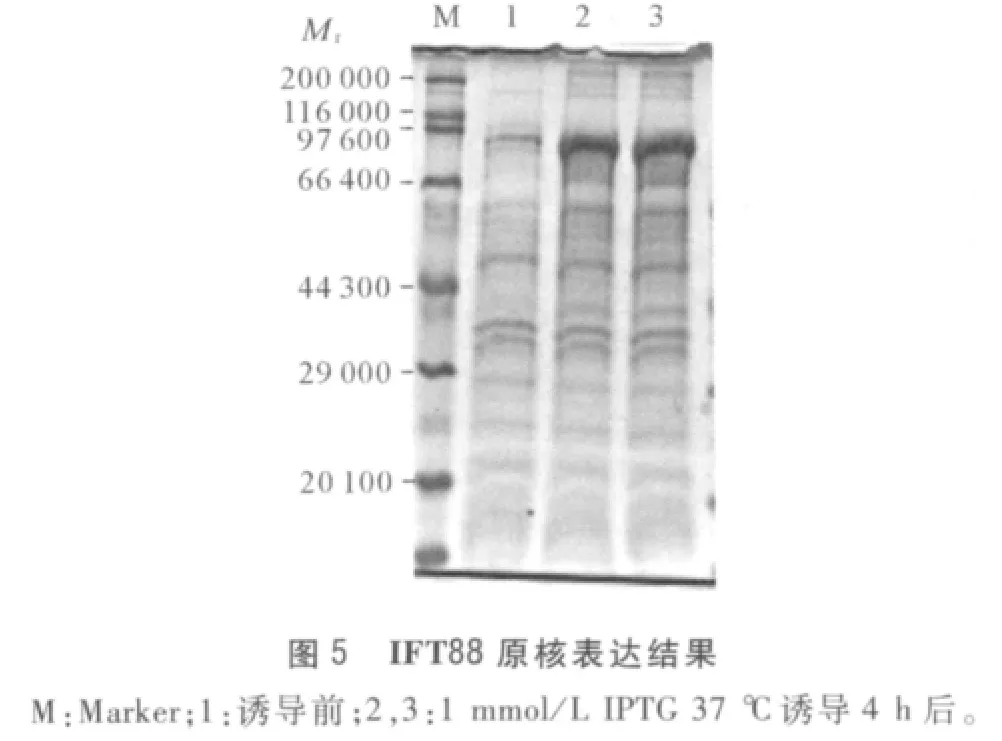
3 讨论
经测序拼接后,得到IFT88 开放阅读框2 400 bp,编码799 个氨基酸,通过DNAMAN 软件对该序列分析得到IFT88 蛋白的相关信息,其蛋白质的相对分子质量为87 907.4,等电点为5.08。在NCBI上比对IFT88 氨基酸序列,发现其具有IFT88 家族所特有的结构域(TPR 结构域),且与其他物种的IFT88 氨基酸序列有较高的同源性(衣藻58%、斑马鱼40%、短尾负鼠40%、黑猩猩43%、人类38%),可见此蛋白序列比较保守。
自Pazour 等[4]发现衣藻IFT88 小鼠同源蛋白Tg737 断裂可以缩短肾脏纤毛并引起多囊肾病以来,关于IFT88 研究越来越多。IFT88 的缺失会导致纤毛缺陷,这种缺陷会导致多囊肾疾病、视觉、嗅觉的功能障碍等多种纤毛疾病,此外,IFT88 在细胞有丝分裂中与纺锤体的定位相关联[8],并且在无纤毛细胞中,IFT88 还能调节细胞G1-S 的转变[9]。虽然关于IFT88 和纤毛疾病的研究已经很多,但是导致这种病变的机制仍不清楚,并且关于纤毛的组装、维持和解聚的机制也有待探究。作者不仅克隆了IFT88 的开放阅读框而且构建其原核表达载体对此蛋白进行诱导表达,又通过实时荧光定量PCR 观察在转录水平上IFT88 与杜氏盐藻鞭毛再生的关系,此实验为进一步纯化IFT88 蛋白、制备抗体及研究IFT88 在鞭毛中的相互作用奠定了基础。
[1]曹木青,潘俊敏.纤毛及纤毛相关疾病研究进展[J].中国细胞生物学学报,2012,34(9):849
[2]Cole DG,Snell WJ.Snapshot:intraflagellar transport[J].Cell,2009,137(4):784
[3]Hao L,Schole JM.Intraflagellar transport at a glance[J].J Cell Sci,2009,122(Pt 7):889
[4]Pazour GJ,Dickert BL,Vucica Y,et al.Chlamydomonas IFT88 and its mouse homologue,polycystic kidney disease gene tg737,are required for assembly of cilia and flagella[J].J Cell Biol,2000,151(3):709
[5]McIntyre JC,Davis EE,Joiner A,et al.Gene therapy rescues cilia defects and restores olfactory function in a mammalian ciliopathy model[J].Nat Med,2012,18(9):1423
[6]Pazour GJ,Baker SA,Deane TA,et al.The intraflagellar transport protein,IFT88,is essential for vertebrate photoreceptor assembly and maintenance[J].J Cell Biol,2002,157(1):103
[7]王翠,李杰,柳丽平,等.杜氏盐藻寡糖基转移酶亚基STT3α 功能结构域的克隆与表达分析[J].生物工程学报,2010,26(6):760
[8]Delaval B,Bright A,Lawson ND,et al.The cilia protein IFT88 is required for spindle orientation in mitosis[J].Nat Cell Biol,2011,13(4):461
[9]Robert A,Margall-Ducos G,Guidotti JE,et al.The intraflagellar transport component IFT88/polaris is a centrosomal protein regulating G1-S transition in non-ciliated cells[J].J Cell Sci,2007,120(Pt 4):628
