高温和低湿对干热河谷7个植被恢复树种光合参数的影响
2013-12-14段爱国张建国何彩云
段爱国,张建国,何彩云
(中国林业科学研究院林业研究所国家林业局林木培育重点实验室,北京100091)
干热河谷是对中国云南、贵州和四川等西南地区气候干燥炎热河谷的统称[1-3]。极端高温与低湿是干热河谷的典型气候特征,也是影响该区域植被恢复树种生长的2个主导环境因子[4-6]。探讨高温或低湿对植被恢复树种光合作用的影响,有助于揭示不同树种对高温或低湿的生理生态响应机制,进而评价和筛选适宜区域发展的抗逆性树种。
在自然条件下,植物光合作用常在温度、湿度和光照等因子协同作用下发生变化[7-8],难以确定其中发挥主导作用的因素[9-12],也不能将光合作用的变化归因于某一环境因子的作用,更难把握单一环境因子对光合作用的影响机制。光合作用对胁迫环境特别敏感,高温或低湿均能对光合作用产生明显的负面影响[13-14]。以往对干热河谷野生树种光合作用的研究通常针对高温和低湿的综合效应,缺乏光合作用对高温或低湿单一因子响应的相关研究。
鉴于此,作者以云南省境内干热河谷区的7个主要植被恢复树种为实验对象,通过人工气候室控制温度与湿度,测定各树种幼苗的光合参数,探讨供试树种光合作用对温度和湿度单一因子的响应规律,评价供试树种对高温的忍耐性差异及典型树种对低湿的生理生态响应规律,以期为区域植被恢复树种的选育及恢复措施的选择提供实验依据。
1 材料和方法
1.1 材料
供试树种为云南省境内干热河谷主要植被恢复树种印楝(Azadirachta indica A.Juss.)、木豆〔Cajanus cajan(Linn.)Huth〕、赤 桉 (Eucalyptus camaldulensis Dehn.)、蓝桉(Eucalyptus globulus Labillardière)、柠檬桉 (Eucalyptus citriodora Hook.)、大 叶 相 思 (Acacia auriculiformis A.Cunn.)和 麻 风 树 (Jatropha curcas Linn.),均为 1 年生幼苗。
1.2 方法
1.2.1 幼苗栽植及干旱处理方法 2006年秋季在云南元谋县采集供试树种的种子,2007年3月播种于中国林业科学研究院温室,培育1年后供试。在每一树种中选择生长较为一致的幼苗6株,栽植于长、宽、高均为20 cm的底部带孔塑料盆中,每盆1株,栽培基质为 W(河沙)∶W(腐殖质土)∶W(草炭土)=5∶3∶2。将同一树种的6盆幼苗分别标记,采取隔天浇水并逐步减少浇水盆数的方法,使10 d后同一树种的6盆幼苗的干旱天数分别为0、2、4、6、8和10 d。在干旱梯度形成后,于7月中旬将供试幼苗移至长3.5 m、宽2.2 m、高3.2 m 的 AGC-2 型人工气候室(浙江大学电器设备厂)内进行高温处理。
1.2.2 高温处理及光合参数测定方法 参照干热河谷干季典型环境条件[15],设置人工气候室的空气相对湿度为 20% 、光照强度为 600 mmol·m-2·s-1、CO2浓 度 为400 mmol·m-2·s-1、温度分别为 30 ℃ 和40℃。将供试幼苗移入人工气候室后,让幼苗在设定条件下放置1个白天,使幼苗完全适应人工气候室的环境条件;晚上关闭光源,于第2天凌晨摘取每盆幼苗的中上层鲜叶2~3片,参照文献[16]采用烘干法测定叶片含水量;重新打开光源并光照30 min后,选取每株幼苗中部叶片3片,用Licor-6400型便携式光合测定系统(美国LI-COR公司)记录叶片的净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr);同步采用OSI-FL便携式调制叶绿素荧光仪(美国Opti-Sciences公司)记录叶绿素荧光参数Fv/Fm(PSⅡ最大光能转换效率)[16],测定前对每一叶片均暗适应约 8 min[17]。
1.2.3 低湿处理及光合参数测定方法 选择麻风树幼苗为实验对象,参照上述方法在温室内进行干旱处理后,将幼苗移入人工气候室;设定人工气候室温度为 25 ℃、光照强度为 600 mmol·m-2·s-1、CO2浓度为 400 mmol·m-2·s-1,空气相对湿度分别设定为20%和50%;按照上述方法进行处理并测定叶片相对含水量,用Licor-6400型便携式光合测定系统分别测定每株幼苗的 Pn、Gs、Ci和 Tr。
1.3 数据处理
采用EXCEL 2003统计分析软件进行线性拟合与数据的统计分析。
2 结果和分析
2.1 高温处理对供试7个树种幼苗叶片光合参数的影响
2.1.1 不同温度条件下叶片净光合速率(Pn)与叶片含水量的关系 在干季或干热季,干热河谷植被恢复树种除常遭受空气低湿胁迫外,土壤干旱胁迫也是主要的影响因素。在人工气候室空气湿度恒定以及高温(30℃和40℃)条件下,供试7个树种幼苗叶片净光合速率(Pn)与叶片含水量的关系见图1。
由图1可知,在30℃和40℃条件下,供试7个树种幼苗叶片的Pn值均随叶片含水量的增加呈逐渐升高的趋势,表明叶片含水量高有利于各树种幼苗叶片Pn的提高。当空气温度为30℃时,叶片含水量达到一定的范围,供试7个树种幼苗的Pn均较高,表明30℃的气温对干热河谷区植被恢复树种干季的光合作用较为适宜;而当空气温度上升至40℃高温时,供试树种幼苗的Pn大幅度下降,表明40℃的高温严重制约了这些树种的光合作用。
2.1.2 非干旱条件下高温对叶片光合参数的影响
为排除盆栽土壤水分胁迫因素,对非干旱条件下(充分浇水,干旱胁迫时间0 d)经高温(30℃和40℃)处理后供试7个树种幼苗叶片的各项光合参数进行测定,结果见表1。
由表1可知:在非干旱条件下,不同树种幼苗的Pn受高温的影响程度不同。其中印楝和赤桉的Pn降幅最小,在40℃条件下Pn分别较30℃条件下降低了22.15%和22.56%,表明这2个树种的光合机构对高温具有较强的抵抗能力,对干热河谷区的高温干旱环境具有很强的潜在适应能力;与30℃条件下相比,在40℃条件下蓝桉及麻风树的Pn降幅最大,分别下降了72.06%和66.09%,表明这2个树种的光合机构受高温的抑制作用最大,对高温干旱环境的适应能力有限;柠檬桉、大叶相思及木豆3个树种幼苗的Pn降幅居中,表明这3个树种具有一定的抵抗高温的能力。
由表1还可见:在非干旱(充分浇水)条件下,当人工气候室内的温度由30℃升至40℃时,供试各树种幼苗叶片的胞间CO2浓度(Ci)均明显上升。根据Farquhar和Sharkey有关光合气孔限制的观点[7],当Pn下降、Ci上升时,不论气孔导度上升或下降,光合限制因子均以非气孔限制因子为主。据此可推断高温是供试树种发生以非气孔限制因子为主导因素的光合限制作用的诱因。在非干旱(充分浇水)条件下,大叶相思、蓝桉和木豆幼苗的气孔导度(Gs)随温度的上升而增大,表明40℃高温导致这3个树种叶片叶肉细胞光合活性的下降。此外,在供试的7个树种中,除印楝外,高温明显加剧了其他6个树种叶片的蒸腾速率(Tr),并导致各树种幼苗叶片水分利用效率的降低。
2.2 低湿处理对麻风树幼苗叶片光合参数的影响
在金沙江干热河谷区,空气相对湿度低、高温、土壤含水量低以及土壤贫瘠共同成为植被恢复工作所面临的环境限制因子。在空气相对湿度20%和50%的条件下,麻风树幼苗叶片光合参数与其叶片含水量的关系见图2。
由图2可见:当人工气候室内的空气相对湿度由20%提高至50%时,麻风树幼苗叶片的Pn、Gs、Ci及Tr均明显提高,但在空气相对湿度20%和50%的条件下,麻风树幼苗叶片的Pn、Gs及Tr均随叶片含水量的下降而降低,表明在其他环境条件一致的情况下,叶片含水量的下降成为麻风树幼苗叶片光合作用减弱的主要限制因素。此外,由图2还可见:随麻风树幼苗叶片含水量的降低,因空气相对湿度不同而导致的叶片Pn的差异逐渐减小。
2.3 高温对供试7个树种幼苗叶片叶绿素荧光参数Fv/Fm的影响
叶绿素荧光参数Fv/Fm指示出PSⅡ反应中心处于关闭时的原初光能转换效率,是叶绿素光化学反应状况的重要描述性指标,该指标具有一定的稳定性,仅在胁迫状态下降低[18]。在人工气候室空气相对湿度恒定以及高温(30℃和40℃)条件下,供试7个树种幼苗叶片Fv/Fm值与叶片含水量的关系见图3。

图1 在30℃和40℃条件下供试7个树种幼苗叶片的净光合速率(Pn)与含水量的关系Fig.1 Relationship between net photosynthesis rate(Pn)and water content of leaf of seedlings of seven tree species tested under 30℃and 40℃conditions
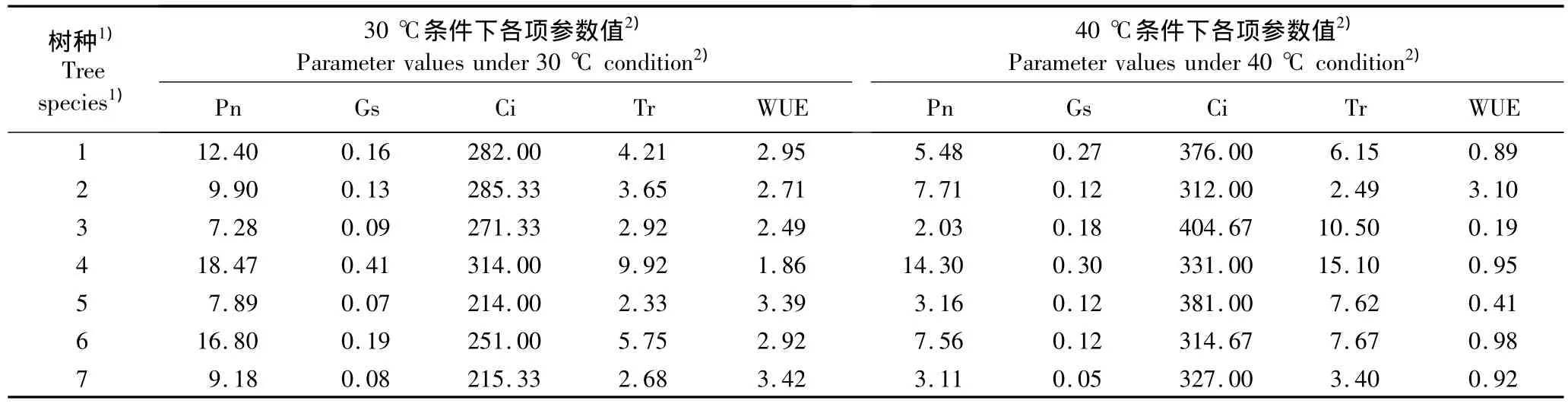
表1 在非干旱条件下不同温度处理(30℃和40℃)对供试7个树种叶片光合参数的影响Table 1 Effect of different temperature treatments(30℃ and 40℃)on photosynthetic parameters of seedling’s leaf of seven tree species tested under non-drought condition

图2 在相对湿度20%和50%的条件下麻风树幼苗叶片的净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)与叶片含水量的关系Fig.2 Relationship between net photosynthesis rate(Pn),stomatal conductance(Gs),intercellular CO2concentration(Ci),transpiration rate(Tr)and water content of leaf of Jatropha curcas Linn.seedling under relative air humidity 20%and 50%conditions

图3 在30℃和40℃条件下供试7个树种幼苗叶片的PSⅡ最大光能转换效率(Fv/Fm)与含水量的关系Fig.3 Relationship between maximal photochemical efficiency of PSⅡ(Fv/Fm)and water content of leaf of seedlings of seven tree species tested under 30℃and 40℃conditions
由图3可见:在温度30℃和40℃的条件下,随叶片含水量的增加,供试7个树种叶片的Fv/Fm值迅速增大,表明叶片含水量高有利于其原初光能转换效率的提高;当温度由30℃升至40℃时,供试7个树种叶片的Fv/Fm值均明显下降,表明40℃高温对各树种的叶绿素光化学效率有明显的抑制作用。相比较而言在供试的7个树种中,在叶片含水量较高的状况下印楝和赤桉的Fv/Fm值受高温的影响较小,而蓝桉、木豆及大叶相思的Fv/Fm值受高温的影响程度相对较大。这与高温对供试树种Pn的影响差异基本一致。
3 讨论和结论
目前,高温对农作物或经济作物光合作用的影响研究较多,而树木光合作用对高温胁迫响应的相关研究尚不多见。以往的研究结果表明:高温往往导致植物净光合速率的下降[13];与28℃条件下相比,在40℃高温条件下2个甘蓝(Brassica oleracea Linn.)品种的净光合速率分别下降7%和24%[19];经40℃高温胁迫处理12 h后,辣椒(Capsicum annuum Linn.)不同品种幼苗的净光合速率均受到明显抑制[20]。本研究结果表明:在光照与湿度条件恒定的条件下,经30℃和40℃高温处理后供试的7个干热河谷植被恢复树种叶片的净光合速率均随叶片含水量的提高呈逐渐增加的趋势,表明叶片含水量高有利于各树种幼苗叶片净光合速率的提高。与30℃气温相比,干热河谷干季常出现的40℃高温严重制约了植株的净光合速率,并加剧了绝大部分树种幼苗的蒸腾作用、导致其水分利用效率降低。
高温胁迫导致植物净光合速率降低可能具有气孔限制和非气孔限制2条途径,但二者没有严格的区分。Berry等[21]认为:高温下气孔导度降低并不是净光合速率降低的主要原因。因为在恒定或较低蒸汽压差下,叶片气孔导度会出现随温度提高而增大的现象,且在光合器官受到破坏时气孔依然开放,这样胞间CO2浓度就不会降低,而气孔限制也不会发生。对于供试的7个干热河谷主要植被恢复树种而言,在土壤水分环境良好的条件下高温显然导致了以非气孔限制为主导因素的光合限制的发生,这是因为供试树种叶片的胞间CO2浓度均随温度的升高表现为增加趋势;经40℃高温处理后供试树种幼苗的Fv/Fm也被不同程度抑制,这一现象表明高温导致了叶片叶肉细胞光合活性的下降,佐证了“光合作用非气孔限制占据主导”的结论。这与霍俊伟等[22]对高温胁迫下黑穗醋栗(Ribes nigrum Linn.)幼苗的相关研究结果一致。而在供试的7个树种中,有3个树种叶片的气孔导度随温度的升高而增大,另有3个树种的气孔导度则随温度的升高而减小,这一现象可能与树种自身的生物学特性及抗热机制差异有关。
干热河谷季节性的气候干旱和土壤水分匮乏是该区域植被恢复的主要限制因子。供试的7个树种幼苗叶片的净光合速率均随叶片含水量的降低明显下降,表明叶片含水量高有利于其净光合速率的提高。张宇等[14]认为:大气湿度越大越有利于植物光合作用和物质积累。本研究结果表明:与空气相对湿度50%的高湿条件相比,空气相对湿度20%的低湿环境明显降低了麻风树幼苗的净光合速率、气孔导度及胞间CO2浓度,且这些光合参数均随其叶片含水量的下降而降低。从树种差异来看,在非干旱(充分浇水)条件下,各树种幼苗的净光合速率受高温的影响程度不同,印楝与赤桉的净光合速率降幅较小、蓝桉及麻风树的净光合速率降幅较大,因而,与蓝桉和麻风树相比,印楝和赤桉适应或忍耐干热河谷高温干旱生境的能力可能更强。这一结论可以从这些树种在干热河谷的实际生长状况得到佐证,蓝桉仅能在干热河谷区温度相对较低的高海拔区域生长良好,而麻风树则以落叶的方式避开了干热季节的大部分时期。
[1]赵俊臣.干热河谷经济学初探[M].香港:香港中国经济文化出版社,1992.
[2]张荣祖.横断山区干旱河谷[M].北京:科学出版社,1992.
[3]李 昆,张春华,崔永忠,等.金沙江干热河谷区退耕还林适宜造林树种筛选研究[J].林业科学研究,2004,17(5):555-563.
[4]马焕成,胥 辉,陈德强,等.元谋干热河谷几种相思和桉树水分消耗量估测[J].林业科技通讯,2000(4):9-12.
[5]高 洁,刘成康,张尚云.元谋干热河谷主要造林植物的耐旱性评估[J].西南林学院学报,1997,17(2):19-24.
[6]金振洲,欧晓昆,区普定,等.金沙江干热河谷种子植物区系特征的初探[J].云南植物研究,1994,16(1):1-16.
[7]许大全.光合作用测定及研究中一些值得注意的问题[J].植物生理学通讯,2006,42(6):1163-1167.
[8]张建国,李吉跃,沈国舫,等.树木耐旱特性及其机理研究[M].北京:中国林业出版社,2000.
[9]SHARP R E,MATTHEWS M A,BOYER J S.Kok effect and the quantum yield ofphotosynthesis:lightpartially inhibitsdark respiration[J].Plant Physiology,1984,75:95-101.
[10]KYEI-BOAHEN S,LADA R,ASTATKIE T,et al.Photosynthetic response of carrots to varying irradiances[J].Photosynthetica,2003,41(2):301-305.
[11]段爱国,张建国,张俊佩,等.金沙江干热河谷主要植被恢复树种叶水势的时空变化规律[J].林业科学研究,2007,20(2):151-159.
[12]战 伟,沙 伟,王 淼,等.降水和温度变化对长白山地区水曲柳幼苗生长和光合参数的影响[J].应用生态学报,2012,23(3):617-624.
[13]刘东焕,赵世伟,高荣孚,等.植物光合作用对高温的响应[J].植物研究,2002,22(2):205-212.
[14]张 宇,宋敏丽,李丽平.亚高温下不同空气湿度对番茄光合作用和物质积累的影响[J].生态学杂志,2012,31(2):342-347.
[15]段爱国,张建国,张守攻,等.干热河谷主要植被恢复树种蒸腾作用[J].生态学报,2009,29(12):6691-6701.
[16]段爱国,杨文忠,张建国,等.苗木离体叶片水分状况的叶绿素荧光参数定量诊断[J].浙江林学院学报,2010,27(4):529-537.
[17]赵会杰,周 琦,于振文.叶绿素荧光分析技术及其在植物光合机理研究中的应用[J].河南农业大学学报,2000,34(3):248-251.
[18]彭方仁,朱振贤,谭鹏鹏,等.NaCl胁迫对5个树种幼苗叶片叶绿素荧光参数的影响[J].植物资源与环境学报,2010,19(3):42-47.
[19]付振书,赵世杰,孟庆伟,等.高温强光下耐热性不同的两个甘蓝品种幼苗光合作用差异的研究[J].园艺学报,2005,32(1):25-29.
[20]潘宝贵,王述彬,刘金兵,等.高温胁迫对不同辣椒品种苗期光合作用的影响[J].江苏农业学报,2006,22(2):137-140.
[21]BERRY J A,BJORKMAN O.Photosynthetic response and adaptation to temperature in higher plants[J].Annual Review of Plant Physiology,1980,31:491-543.
[22]霍俊伟,张壮飞,秦 栋,等.高温胁迫对黑穗醋栗幼苗光合特性的影响[J].东北农业大学学报,2012,43(10):79-84.
