不同光照条件下结缕草全株碳素生理整合格局及其生态效应
2013-12-14何莹莹李德志程立丽
何莹莹,李德志,b,c,①,徐 洁,王 浩,程立丽,刘 微
(华东师范大学:a.资源与环境学院环境科学系,b.上海市城市化生态过程与生态恢复重点实验室,c.浙江天童森林生态系统国家野外科学观测研究站,上海200241)
克隆植物是指在自然条件下能够通过营养繁殖形成多个在遗传上一致、在形态和生理上独立或者潜在独立个体的一类植物[1]。克隆植物的生理整合是指在源-汇关系驱动下物质或资源(如光合同化产物、矿质养分和水分等)通过连接物或间隔子(如根茎及匍匐茎等)在克隆分株间进行运输与分享的过程。生理整合不仅仅是一种能量和物质从高水平向低水平运输直至整个系统达到平衡的被动运输过程,而且是植物对生境高度适应的复杂生理过程[2]。Eriksson等[3]的研究结果表明:如果分株间的资源传递和共享使接受者获得的贡献大于施予者的损失(或对基株适合度造成的损失),那么分株间的资源互享对整个克隆系是有益的,此时的生理整合具有进化上的优势。因此,生理整合作用是克隆植物所具有的重要生理特征之一,同时也是其适应异质性生境的重要生态策略之一。
碳素整合是克隆植物生理整合过程中非常重要的一部分,是克隆植物生存、生长和繁殖的物质基础[1]。克隆植物碳素整合的基本机制具有一定的共同性或相似性,但因克隆植物种类的不同、所处生境以及发育阶段的不同或所受到的干扰和胁迫程度的不同,碳素整合的范围、强度、方向和过程等也有一定的差异[1]。但在均质性生境中生理整合往往不能体现其优势,而只有在异质性生境中生理整合作用才具有一定的优势[1,3-5]。
光合作用是植物从环境中获取物质和能量并实现生长、发育和繁殖的生理基础。对高度分工的高等植物而言,同化产物在植物各器官间的运输和再分配是植物成为协调统一整体的不可缺少的生理过程。对于克隆植物而言,每个分株除了能实现同化产物在源、汇器官间的纵向运输外,在形态上相连的分株间还能发生同化产物的横向运输,这种基于克隆分株构件性生长而形成的生理整合是克隆植物特有的生理过程[6]。光照是影响光合作用的主要因素之一,在野外生境中植物获得的光资源经常是异质性分布的(主要表现为梯度变化或斑块性分布)[7],对植物生长会产生复杂的影响;而对具有双重构件性的克隆植物而言,这种影响则更为特殊。
结缕草(Zoysia japonica Steud.)是一种广泛用于草坪绿化的草种,也是天然放牧场的优良牧草。结缕草为C4多年生匍匐茎型克隆植物,形态结构十分特殊,其基本结构由须根系、匍匐茎、复合节和分蘖叶等部分组成。匍匐茎的基本组成单位称为复合节间,是由1个伸长的节间和1对缩短的节间(其上分别着生被称为A和B的2个分蘖或分株)所组成[8]。在自然环境下,结缕草总是优先发育A分株,继而生长成A分枝;B分株和B分枝只有在养分充足的环境中才能发育,并且总是后于A分株发育。结缕草克隆植株的典型形态结构示意图见图 1[9-10]。
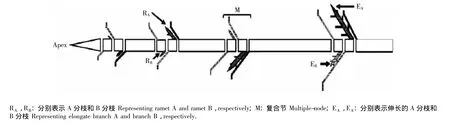
图1 结缕草克隆植株的典型形态结构示意图Fig.1 Diagram of typical morphological structure of clone-plant of Zoysia japonica Steud.
结缕草克隆植株在生长过程中经常处于光照强度不同的生境中,且这种光生境可能具有异质性或斑块性。探究具有生理整合特征的结缕草克隆植株对光照异质性生境的响应不仅具有理论意义,而且对结缕草的草地经营也具有实践指导意义。为此,作者采用13C作为示踪探针,在4种光照处理条件下对13CO2在结缕草克隆植株各构件内的传输和分配特征进行研究,试图揭示克隆分株间以13C指示的光合同化物的生理整合格局特征(如整合的方向、范围和强度等),并分析这些特征对结缕草生态行为可能产生的影响及其适应意义。
1 实验地概况和实验方法
1.1 实验地概况
实验地位于上海市闵行区马桥园艺场内(北纬31°14'、东经 121°29')。该区域属亚热带季风气候,冬冷夏热、四季分明、降水充沛、光温协调、日照较多,全年平均气温16.5℃。实验在玻璃温室内进行;温室内的土壤为黏性砂质土,土壤有效氮、有效钾和有效磷含量分别为158.5、146.1 和149.56 mg·kg-1;温室内的温度和湿度均较室外偏高,光照强度则弱于室外;供试结缕草在温室内的栽培时间由2011年6月份持续至10月份。在栽培过程中,选择植株生长区周围的不同位点定期对温室内的温度、湿度和光照强度进行测定。温度和湿度采用TES-1361型记忆式温湿度计(台湾泰仕电子工业股份有限公司)测定,光照强度采用DRM-FQ型光量子仪(北京澳作生态仪器有限公司)测定。测定时间为每天6:00至18:00,2 h测定1次,结果取平均值。温室内6月份至10月份的平均气温分别为26.8 ℃、34.3 ℃、35.7 ℃、29.2 ℃和27.1℃,平均空气相对湿度分别为64.5%、69.2%、67.3%、70.6%和 68.4%,平均光量子密度分别为 582、590、643、513 和 492 μmol·m-2·s-1。
1.2 材料
实验所用结缕草购自上海市秀谷生物科技有限公司,供试的所有植株均取自同一结缕草基株(由小分株切段经培养后形成的较大植株)。从基株中选取大小基本一致且根系比较完整、形态特征基本相同的分株,分别栽入土地已经过平整的实验小区内。
自然条件下结缕草总是优先发育A分株,每个复合节上必定发育出A分株但不一定发育出B分株,因此,作者在进行13CO2引入时统一选择从复合节上的A分株引入,即第7复合节、第17复合节和第24复合节上的A分株,引入点分别为R7、R17和R24。
1.3 方法
1.3.1 处理方法 共设置4种光照处理:处理Ⅰ—所有分株均进行光照处理,即全光照;处理Ⅱ—所有分株均进行遮光处理,即全遮光;处理Ⅲ—仅对引入13CO2的复合节分株进行遮光处理,即引入点遮光处理;处理Ⅳ—仅对引入13CO2的复合节分株进行光照处理,即引入点光照处理。遮光处理方法均为在需要遮光的植株部位上方设置1层遮光率70%的遮阳网。
用丰度99%的Ba13CO3和过量稀H2SO4制备13CO2待用。在结缕草植株收获前期,在各处理组中分别选取主匍匐茎上第7(近基部)、第17(居中部)和第24(近顶端)复合节的A分株进行13CO2引入。于2011年10月30日上午10:00整,将预先制备好的13CO2用一次性注射器定量注入事先已安置在指定分株上且体积已知的密闭叶室中,使密闭叶室内的13CO2浓度维持在 350 μmol·mol-1;引入13CO21 h后,撤下密闭叶室,24 h后收获所有处理的克隆植株。实验共设12组处理,每组处理3次重复。
1.3.2 测定方法 收获样株后,对复合节及分株进行标记并按照生长顺序进行编号,整体烘干后再按编号分别将分株、匍匐茎及根系分开并研磨成粉;然后用DELTAplusXP型稳定同位素质谱仪(美国菲尼根质谱公司)测定样品的13C丰度。测量时从R1分株开始,每隔1个复合节测量不同分株的各项指标,即测量第 1、3、5、7、9、……、25 复合节上的 A 分株叶片、匍匐茎和根系的各项指标。
1.4 数据处理
按照以下公式计算13C的丰度值:
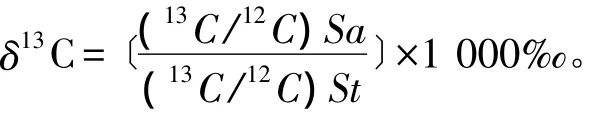
式中:(13C/12C)Sa为样品的C同位素比值;(13C/12C)St为标准物质(即PDB)的C同位素比值。
为分析13C引入后在结缕草植株内的分配格局,采用传输陡度和传输范围2个指标进行度量。其中,对引入复合节各构件器官样品的δ13C值向两侧(基部和顶部)其他复合节的相应构件器官递减的δ13C值作线性回归,其方程斜率的绝对值即为传输陡度;而2条线性回归方程分别与横轴的交点与引入位点之间的复合节数即为向基或向顶的传输范围[11]。
采用SPSS 13.0统计分析软件对数据进行单因素方差分析(P<0.05)。
2 结果和分析
2.1 碳素在结缕草主匍匐茎中的整合格局
2.1.1 从基部分株(即第7分株)叶片引入13CO2后13C的整合格局 结缕草克隆植株基部分株是指在结缕草克隆植株上生长发育较早的分株。通过基部分株叶片引入13CO2,旨在检验从较老分株中同化的碳素资源在整个克隆植株体内的传输与分享的基本格局。本研究所选基部分株为主匍匐茎上靠近基部的第7复合节分株。在4种光照处理条件下由第7复合节A分株引入13CO2后13C在主匍匐茎根、茎和叶片中的分布格局见图2。
由图2可见:对所有复合节分株均进行全光照处理(处理Ⅰ),从第7复合节分株引入13CO2后叶片中的δ13C值向顶或向基均逐渐降低;而对所有复合节分株均进行遮光处理(处理Ⅱ),叶片中的δ13C值向顶或向基均呈先降后升的趋势;仅对引入13CO2的复合节分株进行遮光处理(处理Ⅲ),叶片中的δ13C向基方向逐渐降低、向顶方向则呈先降后升的趋势;仅对引入13CO2的复合节分株进行光照处理(处理Ⅳ),叶片中的δ13C值向基方向逐渐降低、向顶方向则呈先平缓后降低的趋势。但在4种光照处理条件下,引入点分株叶片中的δ13C值均显著高于相邻的其他分株叶片的δ13C值(P<0.05)。在4种光照处理条件下,从第7复合节分株引入13CO2后,根和匍匐茎中的13C传输格局相似,即二者的δ13C值向顶或向基方向均逐渐降低。
通过比较还可见:13C传输到相邻分株或者更远分株后,复合节分株叶片中的δ13C值均小于同一复合节根和匍匐茎中的δ13C值;且同一复合节分株中根的δ13C值均高于匍匐茎中的δ13C值。此外,在处理Ⅳ条件下,所有分株各器官的δ13C值均显著高于另外3组处理中同一复合节分株相同器官的δ13C值(P<0.05)。
2.1.2 从中部分株(即第17分株)叶片引入13CO2后13C的整合格局 结缕草克隆植株的中部分株是指在结缕草克隆植株上生长发育处于中期的分株,选择这类分株引入13CO2,旨在检验从发育程度中等的分株中获取的碳素资源在整个克隆植株体内的传输与分享的格局特征。本研究所选用的中部分株为主匍匐茎上近中部的第17复合节分株。4种光照处理条件下由第17复合节A分株引入13CO2后13C在主匍匐茎根、茎和叶片中的分布格局见图3。
由图3可见:在4种光照处理条件下,从中部复合节分株(位于第17复合节)引入13CO2后,13C在整个克隆植株内的分布格局及传输基本特征与从其基部复合节(第7复合节)分株引入13CO2有相似之处,但各器官中的δ13C值向基或向顶双向均呈逐渐下降的趋势,没有出现先降后升的格局。在分株叶中δ13C值的变化特征与引入点位于基部(第7)复合节时类似,即引入点分株叶中的δ13C值显著高于相邻其他分株叶中的δ13C值(P<0.05);13C传输到相邻分株或更远分株后,同一复合节分株叶中的δ13C值均小于同一复合节根和匍匐茎中的δ13C值。

图2 在不同光照条件下从结缕草主匍匐茎基部(第7)复合节分株引入13CO2后13C在主匍匐茎不同器官中的分布格局Fig.2 The13C distribution pattern in different organs of main stolon of Zoysia japonica Steud.after fed with13CO2at the basal(the seventh)multiple-node ramet under different light treatments
2.1.3 从梢部分株(即第24分株)叶片引入13CO2后13C的整合格局 结缕草克隆植株的梢部分株是指在结缕草克隆植株内生长发育较晚的分株,选择这类分株引入13CO2,旨在检验从发育较晚的分株中获取的碳素资源在整个克隆植株体内的传输与分享的格局特征。本研究所选用的梢部分株为主匍匐茎上近顶部的第24复合节分株。4种光照处理条件下由第24复合节A分株引入13CO2后13C在主匍匐茎根、茎和叶片中的分布格局见图4。
由图4可见:在4种光照处理条件下,从结缕草克隆植株的梢部(第24复合节)分株引入13CO2,主匍匐茎中的13C总体仍呈双向传输趋势且更倾向于向顶传输,各器官中的13C传输格局与引入点位于基部和中部相比有异同。在4种光照处理条件下,在分株叶中δ13C值呈向顶或向基双向逐渐降低,但向顶方向的δ13C值均明显高于向基方向的δ13C值,说明从克隆植株梢部分株引入δ13CO2后,13C更倾向于传输给位于顶端一侧的分株;而在向顶传输过程中,克隆植株的顶端虽然与引入点距离较远,但其δ13C值却未随13C传输距离的增大而减小反而有所升高,其δ13C值甚至明显高于与引入点分株相距最近的分株(图4),总体呈现出明显的先降后升的趋势(由于顶端新叶细小甚至未展开,仅以芽的状态存在,因此本实验中将整个顶端作为匍匐茎测量,不再区分复合节根和分株叶)。此外,从梢部分株引入13CO2后,复合节根中的δ13C值均小于匍匐茎,这与从基部分株和中部分株引入13CO2后13C的传输格局特征恰好相反。
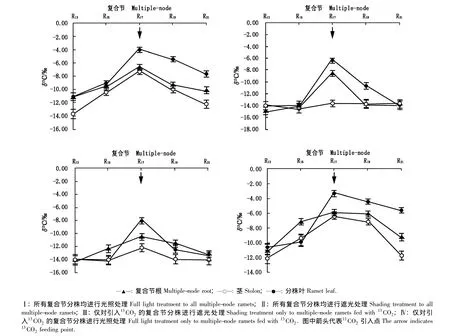
图3 在不同光照条件下从结缕草主匍匐茎中部(第17)复合节分株引入13CO2后13C在主匍匐茎不同器官中的分布格局Fig.3 The13C distribution pattern in different organs of main stolon of Zoysia japonica Steud.after fed with13CO2at the middle(the seventeenth)multiple-node ramet under different light treatments
2.2 碳素在结缕草主匍匐茎中的传输特征
在4种光照处理条件下由结缕草基部分株(第7复合节)、中部分株(第17复合节)和梢部分株(第24复合节)的A分株引入13CO2后13C在结缕草主匍匐茎根、茎和叶片中的传输陡度及传输范围分别见表1和表2。
在4种光照处理条件下,从基部(第7复合节)分株引入13CO2后,13C均呈现出以引入点为起始点沿匍匐茎双向运输的趋势,但向顶传输的陡度值均小于向基传输的陡度值,说明13C的传输更倾向于向顶方向(表1);同时,向顶传输的距离也普遍大于向基传输的距离(表2),进一步佐证了13C传输的向顶趋势;其中,在分株叶中13C传输呈现出脉冲式格局,13C传输陡度值显著大于复合节根和匍匐茎的13C传输陡度值(P<0.001)。在匍匐茎中,13C呈现出平缓和渐进式的传输格局,传输距离也最远,向顶传输能够到达第14复合节,向基传输能够到达第2复合节,比复合节根和分株叶中的传输距离平均多1~2个复合节。此外,在不同光照处理组中,处理Ⅱ的分株叶13C向基或向顶传输的陡度值均最高;而在处理Ⅳ中,根及匍匐茎中13C向基或向顶传输的陡度值均最高,其分株叶中13C的陡度值仅次于处理Ⅱ,且在处理Ⅳ中各部位13C的传输距离均最远。

图4 在不同光照条件下从结缕草主匍匐茎梢部(第24)复合节分株引入13CO2后13C在主匍匐茎不同器官中的分布格局Fig.4 The13C distribution pattern in different organs of main stolon of Zoysia japonica Steud.after fed with13CO2at the distal(the twenty-fourth)multiple-node ramet under different light treatments
在4种光照处理条件下,从中部(第17复合节)分株引入13CO2后,13C向顶或向基传输的陡度值与引入点位于基部时相似,即向顶传输较远且下降趋势平缓,向基传输较近且下降陡度大(表1);在分株叶中13C的传输也表现出脉冲式格局,而且引入点分株叶δ13C值越大,引入点两侧分株叶δ13C值下降越快。在匍匐茎中13C的传输距离相对较远,向顶方向最远能够到达第25复合节、向基方向最远能够到达第11复合节(表2)。从不同部位13C的陡度值看,分株叶中13C的陡度值最大、匍匐茎中13C的陡度值最小,表明在匍匐茎中13C的传输最平缓。此外,在不同光照处理组中,处理Ⅲ中分株叶13C向基或向顶传输的陡度值均最高;而处理Ⅳ中根及匍匐茎13C向基或向顶传输的陡度值均最高,且在处理Ⅳ中各器官13C的传输距离均最远。
在4种光照处理条件下,从梢部(第24复合节)分株引入13CO2后,13C向顶或向基的传输也呈现“向顶传输较远、向基传输较近,且向顶传输的陡度值总体上小于向基传输的陡度值”的规律(表1)。由不同器官的比较可见:总体上13C在匍匐茎中的传输距离相对较远,向顶方向最远能够到达第36复合节,向基方向最远能够到达第19复合节(表2)。从不同器官13C的陡度值看:总体上分株叶13C的陡度值最大,而根和匍匐茎13C的陡度值在不同光照条件下表现出不同的规律性。此外,在不同光照处理组中,处理Ⅰ中分株叶13C向基或向顶传输的陡度值均最高;而处理Ⅳ中根向顶及匍匐茎向基的13C传输陡度值均最高,分株叶13C的陡度值仅次于处理Ⅰ,且在处理Ⅳ中各器官13C的传输距离均较远或最远。
表1 在不同光照条件下从结缕草主匍匐茎的3个复合节分株引入13CO2后13C在主匍匐茎不同器官内的传输陡度值(±SD)Table 1 The translocation gradient value of13C in different organs of main stolon of Zoysia japonica Steud.after fed with13CO2at three multiple-node ramets under different light treatments(±SD)

表1 在不同光照条件下从结缕草主匍匐茎的3个复合节分株引入13CO2后13C在主匍匐茎不同器官内的传输陡度值(±SD)Table 1 The translocation gradient value of13C in different organs of main stolon of Zoysia japonica Steud.after fed with13CO2at three multiple-node ramets under different light treatments(±SD)
1)Ⅰ:所有复合节分株均进行光照处理 Full light treatment to all multiple-node ramets;Ⅱ:所有复合节分株均进行遮光处理Shading treatment to all multiple-node ramets;Ⅲ:仅对引入13CO2的复合节分株进行遮光处理Shading treatment only to multiple-node ramets fed with13CO2;Ⅳ:仅对引入13CO2的复合节分株进行光照处理Full light treatment only to multiple-node ramets fed with13CO2.2)同列中不同的小写字母表示差异显著(P<0.05)Different small letters in the same column mean the significant difference(P<0.05).
?
表2 在不同光照条件下从结缕草主匍匐茎的3个复合节分株引入13CO2后13C在主匍匐茎不同器官内的传输范围(±SD)Table 2 The translocation range of13C in different organs of main stolon of Zoysia japonica Steud.after fed with13CO2at three multiplenode ramets under different light treatments(±SD)

表2 在不同光照条件下从结缕草主匍匐茎的3个复合节分株引入13CO2后13C在主匍匐茎不同器官内的传输范围(±SD)Table 2 The translocation range of13C in different organs of main stolon of Zoysia japonica Steud.after fed with13CO2at three multiplenode ramets under different light treatments(±SD)
1)Ⅰ:所有复合节分株均进行光照处理Full light treatment to all multiple-node ramets;Ⅱ:所有复合节分株均进行遮光处理Shading treatment to all multiple-node ramets;Ⅲ:仅对引入13CO2的复合节分株进行遮光处理Shading treatment only to multiple-node ramets fed with13CO2;Ⅳ:仅对引入13CO2的复合节分株进行光照处理Full light treatment only to multiple-node ramets fed with13CO2.2)用13C传输过程中经过的复合节数表示传输范围,表中的数据为传输到达的复合节,例如向基3或向顶12表示向基传输至第3复合节或向顶传输至第12复合节The translocation range is represented with multiple-node number passed during13C translocation process,the datums in this table mean that13C reaches multiple-nodes,and for example,basal 3 or distal 12 mean that13C reaches the third multiplenode toward basal or reaches the twelfth multiple-node toward distal.
?
3 讨论和结论
克隆植物的碳素整合格局既可以单向(向基或向顶)也可以双向[12-14],既可在同一克隆的分株间发生广泛的整合也可在相对独立的生理整合单位内发生局部的整合[11]。另外,碳素整合格局不仅存在种间差异,同种克隆植物的碳素整合格局会因环境资源条件、植株生长发育阶段等因素的不同发生适应性调整或改变[2,15-17]。但 Touchette 等[18]的研究结果表明:虽然存在碳素的双向传输,但向顶传输更利于整株植株的生长发育,更具有适应性优势。
本研究结果表明:在不同光照处理条件下,处于3个生长发育阶段的结缕草克隆分株同化13CO2后13C都有向基和向顶的双向运输趋势,且向顶传输更加明显、递降趋势更加平缓、传输距离更远。13CO2引入点处的复合节分株叶中的δ13C值总体上高于同一复合节的根和匍匐茎(P<0.001),并且也显著高于相邻的分株叶(P<0.05)。在13C传输到相邻分株后,13C总量减少,分株叶中δ13C值急剧下降并小于同一复合节根和匍匐茎的δ13C值,说明当传输至复合节的13C总量减少到一定程度后,分配给复合节各器官的13C比例关系也会发生变化,而且叶中的δ13C值并不一定最高,这一现象与引入点处各器官δ13C值的变化状况不同。这一结果表明:结缕草克隆植株的13C分配策略有时具有保守性,在复合节总体C资源有限的情况下,将有限的C资源保留在根和匍匐茎中其风险可能比保留在叶片中更低,因为叶片中的C很容易随叶片枯萎或掉落而流失,而复合节根和匍匐茎中的C则可以保存较长时间。
当13CO2从克隆植株的中部和基部分株叶引入时,向基和向顶方向复合节根中的δ13C值均大于匍匐茎中的δ13C值;而当13CO2引入点位于梢部分株叶时,向基和向顶方向复合节根中的δ13C值均小于匍匐茎中的δ13C值,表明发育较早的复合节根需要分享和消耗较多的叶同化C才能维持其吸收功能,因此复合节根的δ13C值大于匍匐茎的δ13C值。而发育较晚的分株分配给复合节根的同化C量则较少,一方面可能由于新生复合节根只需消耗少量C就可维持其吸收功能,另一方面也可能因为这一阶段复合节根的数量不多。但最主要的原因是:与衰老的复合节相比,新生复合节是更强力的“汇”,对营养物质的竞争能力更强,可以从衰老复合节获得一定营养,且不必将更多的生物量投入到根中进行营养吸收。但无论如何,把较多的C资源分配给匍匐茎对于结缕草克隆植株的快速生长以及搜索和占据更多的生境空间是有利的。匍匐茎作为同化产物传输的通道,自身并不截留太多的C,实验中所检测到的13C大多处于流动状态,并非真正被匍匐茎截留。
长期处于胁迫环境条件下,有些克隆植物会中断或减少克隆分株间的资源传输而将资源更多地用于抵御不利环境[19-20]。Allessio 等[21]认为:生长在干旱沙地的Dupontia fischeri R.Br.在产生新分株时很少依赖相邻分株提供的资源,这有利于减少资源转移时的消耗。结缕草在对不同浓度的氮素进行吸收和传输时也表现出了这种特征[5]。本实验结果表明:在全遮光和引入点分株遮光条件下,13C的传输强度均有所降低,因为引入点分株自身光合作用受到了抑制,因此也减少了对其他分株同化产物支持。这一现象与白车轴草(Trifolium repens Linn.)和野牛草〔Buchloe dactyloides(Nutt.)Engelm.〕有相似之处[22-23],野牛草分株间同化物的生理整合也是建立在优先满足分株自身生长代谢需要的基础上。在仅对引入13CO2的复合节分株进行遮光处理的条件下(处理Ⅲ),当13CO2引入点位于结缕草克隆植株中部分株叶时,只有很少部分13C被传输到其他分株中(在引入点位于基部时也有该现象但不十分明显);而处于全光照条件下的其他分株的光合作用较强,甚至可能还会将光合产物传输给引入点分株以支持其生长。在仅对引入13CO2的复合节分株进行光照处理的条件下(处理Ⅳ),其他分株总是具有最高的δ13C值,甚至克隆植株顶端分株的δ13C值也最高,这正是因为引入点以外的分株都处于遮光状态,导致自身光合作用减弱,形成C“汇”拉力,需要其他分株的C供应。
当结缕草克隆植株的梢部分株作为13CO2引入点时,13C虽然还呈双向传输趋势,但向顶的传输强度更大,甚至顶端的δ13C值比紧邻引入点的分株还高,13C向顶传输呈现出先降后升的趋势。这一现象完全符合植物光合产物分配的2个基本原则:一是光合产物总是优先供应给植物的生长中心;二是光合产物总是就近供应。结缕草克隆植株的顶端生长点或幼株因自身的自养能力尚未完全形成,相对于其他分株而言是C同化产物强有力的“汇”,它们能够从成熟分株处获取同化产物,对于其建立分株和顺利渡过生长脆弱期十分重要,对于克隆植株快速搜索生境空间也是十分有利的。
通过以上的实验结果及讨论分析可见:从结缕草克隆植株不同生长发育阶段的分株引入13CO2后,13C有向基和向顶的双向传输趋势。在不同光照条件下,13C生理整合格局不尽相同;当13CO2引入点分株比其他分株受光更充分时,引入点分株的δ13C值高且对其他分株的13C传输能力也强;当引入点分株比其他分株受光少时,其自身的δ13C值低且对其他分株的13C传输能力也弱。在不同光照处理条件下,主匍匐茎上不同器官中13C的传输趋势有一定的差异。引入点分株叶的δ13C值总体上显著高于同一复合节的根和匍匐茎;而13C向顶和向基方向传输后,分株叶的δ13C值急剧下降,甚至在同一复合节内叶的δ13C值低于其分株根和匍匐茎。当13CO2引入点位于主匍匐茎的基部和中部分株叶时,分株根的δ13C值均大于同一复合节的匍匐茎;而当13CO2引入点位于主匍匐茎的梢部分株叶时,分株根的δ13C值均小于同一复合节的匍匐茎。
[1]董 鸣.异质生境中的植物克隆生长:风险分摊[J].植物生态学报,1996,20(6):543-548.
[2]MARSHALL C,SAGAR G R.The distribution of assimilates in Lolium multiflorum Lam.following differential defoliation[J].Annals of Botany,1968,32(4):715-719.
[3]ERIKSSON O,JERLING L.Hierarchical selection and risk spreading in clonal plants[M]∥Van GROENENDEAL J,De KROON H.Clonal Growth in Plants:Regulation and Function.Hague:SPB Academic Publishing,1990:79-94.
[4]盛丽娟,李德志,朱志玲,等.克隆植物的碳素生理整合及其生态学效应[J].应用与环境生物学报,2007,13(6):888-894.
[5]HOSHINO M.Translocation and accumulation of assimilates and phosphorus in Ladino clover[J].Bulletin of the National Grassland Research Institute,1974,5(4):35-84.
[6]LI D Z,TAKAHASHI S.Particularities of clonal plant species induced by physiological integration[J].Journal of Japanese Society of Grassland Science,2003,49(4):395-402.
[7]CALDWELL M M,PEARCY R W.Exploitation of Environmental Heterogeneity by Plants:Ecophysiological Processes Above and Belowground[M].San Diego:Academic Press,1994:98-106.
[8]LI D Z,ITO M,ZHU T C.Impacts of soil nutrient heterogeneity pattern on the growth of clonal plant of Zoysia japonica[J].草业学报,2006,15(4):136-141.
[9]李德志,申芳芳,王长爱,等.克隆植物结缕草在两种环境中的生长发育特征[J].华东师范大学学报:自然科学版,2006(6):117-124.
[10]SHOJI S.Ecological studies on the Zoysia type grassland.4.Development and growth of the stolon of Zoysia japonica Steud.[J].Report of Institute of Agriculture Research of Tohoku University,1976,27(5):49-59.
[11]李立科,李德志,赵鲁青,等.克隆植物结缕草主匍匐茎中氮素的传输特征[J].东北林业大学学报,2011,39(11):40-45.
[12]PRICE E A C,GAMBLE R,WILLIAMS G G,et al.Seasonal patterns of partitioning and remobilization of14C in the invasive rhizomatous perennialJapanese knotweed〔Fallopia japonica(Houtt.)Ronse Decraene〕[J].Evolutionary Ecology,2002,15:347-362.
[13]BENDER M M.Variation in the13C/12C ratios of plants in relation to the pathway of photosynthetic carbon dioxide fixation[J].Phytochemistry,1971,10(6):1239-1244.
[14]SMITH B N,EPSTEIN S.Two categories of13C/12C ratios for higher plants[J].Plant Physiology,1971,47(3):380-384.
[15]YI X F,YANG Y Q,ZHANG X A,et al.No C4plants found at the Haibei Alpine Meadow Ecosystem Research Station in Qinghai,China:evidence from stable carbon isotope studies[J].植物学报,2003,45(11):1291-1296.
[16]XU C Y,SCHOOLER S S,Van KLINKEN R D.Effects of clonal integration and light availability on the growth and physiology of two invasive herbs[J].Journal of Ecology,2010,98(4):833-844.
[17]STUEFER J F,De KROON H,DURING H J.Exploitation of environmental heterogeneity by spatial division of labour in a clonal plant[J].Functional Ecology,1996,10(3):328-334.
[18]TOUCHETTE B W,MOODY J W G,BYRNE C M,et al.Water integration in the clonal emergent hydrophyte,Justicia americana:benefits of acropetal water transfer from mother to daughter ramets[J].Hydrobiologia,2013,702(1):83-94.
[19]李明财,易现峰,李来兴,等.基于稳定碳同位素技术研究青藏高原东部高寒区植被的光合型[J].西北植物学报,2004,24(6):1052-1056.
[20]王昱生,洪锐民,黄大明,等.羊草种群克隆分株之间光合产物的转移[J].生态学报,2004,24(5):900-907.
[21]ALLESSIO M L,TIESZEN L L.Patterns of carbon allocation in an arctic tundra grass,Dupontia fischeri(Gramineae),at Barrow,Alaska[J].American Journal of Botany,1975,62(8):797-807.
[22]CHAPMANA D F,ROBSON M J,SNAYDON R W.Physiological integration in the clonal perennial herb Trifolium repens L.[J].Oecologia,1992,89(3):338-347.
[23]钱永强,孙振元,韩 蕾,等.异质水分环境下野牛草相连分株间光合同化物的生理整合及其调控[J].生态学报,2010,30(15):3966-3973.
