皂荚天然群体间种实表型特性及种子萌发的差异分析
2013-12-14林富荣郑勇奇
李 伟,林富荣,郑勇奇,①,孙 圣
(1.中国林业科学研究院林业研究所国家林业局林木培育重点实验室,北京100091;2.青岛农业大学园林与林学院,山东青岛266109)
种子是植物界演化的最高阶段,是种子植物生活史中的一个重要生育期,对植物群落的建立有决定性作用;而种子萌发过程是建立种子植物群落的关键环节,是实现种群更新和物种延续的重要阶段。种子的大小对于种子的萌发能力及幼苗更新能力起重要作用[1-2],物种之间种子大小有别,而同种植物在不同环境下种子大小也有所不同[3]。群体间种子性状的差异是由地理因子、气候条件、土壤条件、生境以及可利用资源量等多种因素决定的[4-5]。而种子在大小、质量和萌发能力等方面的变异使得种子能逃避不利的环境条件,提高物种的适应能力,进而提高其生存能力[1],对植物幼苗的存活、种群的更新和植被的发展动态具有重要作用[6-7]。
休眠是种子内在的固有特性之一,而萌发是种子对环境条件和萌发需求二者叠合时的响应[8],环境的异质性导致同一物种发生形态变异和遗传差异,产生了不同的生态型,使种子萌发特性存在明显差异,在同种植物的群体间主要反映为种子休眠程度的不同。物种有其独特的种子萌发和休眠策略,以增强其对环境的适应能力。同种植物群体间种子萌发特性的差异受分布区气候条件、经纬度、海拔高度和受干扰程度等外界环境因素的影响,因此,研究生境对群体间种实表型特征和种子萌发特性的影响有助于理解物种对环境的适应机制。
皂荚(Gleditsia sinensis Lam.)为豆科(Fabaceae)皂荚属(Gleditsia Linn.)植物,为中国特有种,自中国北部至南部及西南均有分布,多生长于平原、山谷及丘陵地区。皂荚有很强的抗逆性[9],是优良的中药材和工业原料[10],也是经济林、用材林、防护林及园林绿化的理想树种。皂荚虽分布广泛,但由于人为过度采伐及其自然群体种子萌发困难,目前仅保留残次疏林、个体(丛、簇)或散生木,群体处于濒危状态。迄今为止,对皂荚的研究主要集中在引种栽培、种实和皂刺特性与成分[11-12]、抗逆性[9]和种源研究[13]等方面,对于皂荚不同群体内和群体间表型变异及不同群体的种子萌发差异的研究鲜有报道[14]。
目前,研究者对不同皂荚群体在长期适应各类生境的过程中表型特征是否会发生变化、在不同的生境中其种子的萌发行为是否产生变异等问题尚不清楚。为此,作者拟通过分析不同皂荚群体的种实表型特征及种子萌发行为,以期探讨影响皂荚种实表型特征及种子萌发的因素,阐明不同生境下皂荚种子和果实的生态适应机制,为该种类的资源保护和合理利用提供实验依据。
1 材料和方法
1.1 材料
供试的8个皂荚自然群体分别位于湖北京山(HB)、贵州兴义(GZ)、重庆秀山(CQ)、四川成都(SC)、广西桂林(GX)、湖南城步(HN)、甘肃天水(GS)和山东费县(SD),各群体的地理位置及主要气候条件见表1。每个群体选择20株样株,共160株样株,株间距50 m以上;在每个样株树冠中上部南向位置采集50~80个荚果,于2011年11月完成所有样品的采集。
1.2 方法
1.2.1 种实表型性状测定 测定指标包括荚果长度、宽度和厚度以及种子的长度、宽度、厚度和百粒质量。在每个群体的每一样株上随机选择30个荚果,用游标卡尺分别测定荚果的最长、最宽及最厚处,即为荚果的长度、宽度和厚度,测量精度0.1 cm。在每个群体的每一样株上随机选择30粒种子,采用Epson 4990型平面扫描仪(爱普生中国有限公司)以及WinSeedle图像分析软件(加拿大Regent Instrument公司)测定种子的长度、宽度和厚度;每一样株随机抽取100粒种子,用1/100电子天平称取种子百粒质量,重复8次。
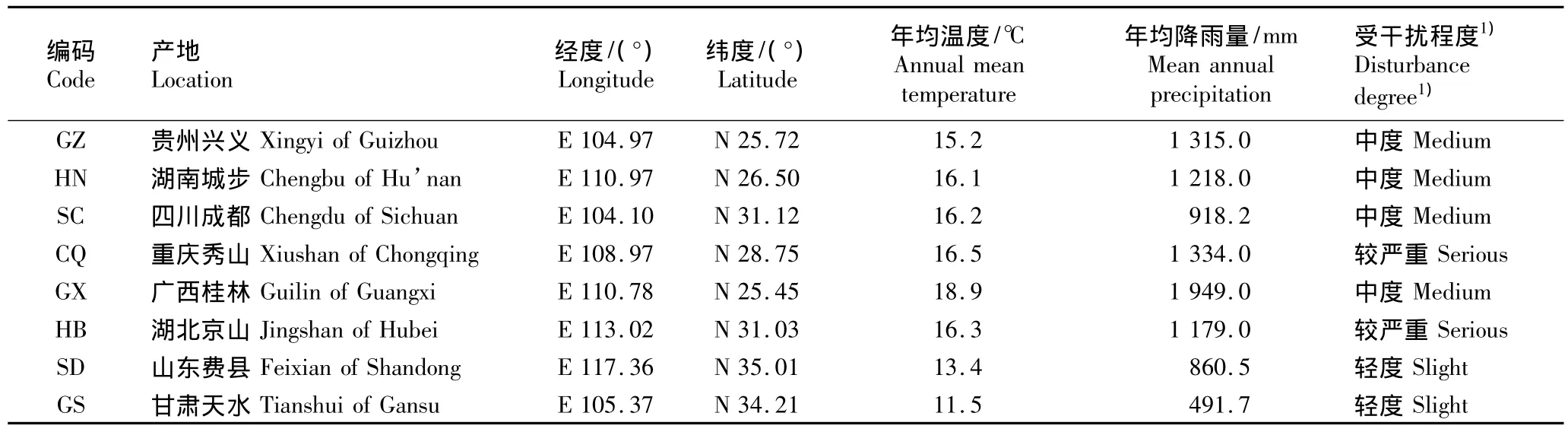
表1 8个皂荚天然群体的地理位置及主要气候条件Table 1 Geographical location and main climatic condition of eight natural populations of Gleditsia sinensis Lam.
1.2.2 种子萌发率测定 用锋利刀片切去种子远离胚根一端的种皮直至露白,作为种皮损伤处理,以未经处理的种子为对照;将种子分别置于光照培养箱中,培养箱设置4个昼/夜温度梯度:35℃/20℃、30℃/15℃、25℃/10℃和15℃/5℃,每天光照12 h;每隔2 d统计萌发种子的数量并将已萌发的种子移除,持续观察30 d。每个群体每一处理30粒饱满种子,各重复3次。
1.3 数据分析
采用SPSS统计分析软件进行实验数据的分析,采用EXCEL 2010数据处理软件作图。用Turkey多重比较方法对皂荚群体间果实和种子性状及种子萌发率等变量的差异进行比较,并对这些指标进行方差分析和相关性分析。
2 结果和分析
2.1 皂荚天然群体间荚果和种子特性的差异分析
8个皂荚天然群体间果实特性和种子特性的比较结果见表2。由表2可知:8个皂荚群体间的荚果长度(F=41.32,P<0.01)、宽度(F=17.54,P<0.01)、厚度(F=32.88,P<0.01)和质量(F=18.12,P<0.01)均有极显著差异,各群体间种子的长度(F=12.99,P<0.01)、宽度(F=21.04,P<0.01)、厚度(F=17.00,P<0.01)和百粒质量(F=151.07,P<0.01)也均存在极显著差异。
表2 8个皂荚天然群体的荚果和种子表型性状的比较(±SD)Table 2 Comparison of phenotypic characteristics of pods and seeds of eight natural populations of Gleditsia sinensis Lam.(±SD)
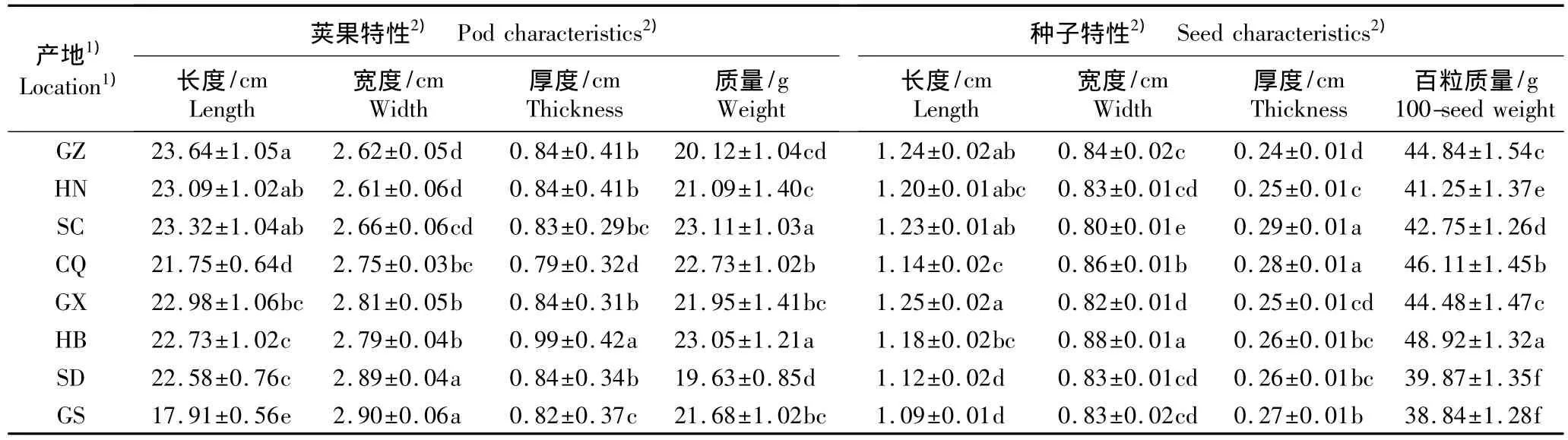
表2 8个皂荚天然群体的荚果和种子表型性状的比较(±SD)Table 2 Comparison of phenotypic characteristics of pods and seeds of eight natural populations of Gleditsia sinensis Lam.(±SD)
1)GZ:贵州兴义Xingyi of Guizhou;HN:湖南城步Chengbu of Hu’nan;SC:四川成都Chengdu of Sichuan;CQ:重庆秀山Xiushan of Chongqing;GX:广西桂林Guilin of Guangxi;HB:湖北京山Jingshan of Hubei;SD:山东费县Feixian of Shandong;GS:甘肃天水Tianshui of Gansu.2)同列中不同的小写字母表示差异显著(P<0.05)Different small letters in the same column indicate the significant difference(P<0.05).
?
贵州群体的荚果最长,达(23.64±1.05)cm;甘肃群体的荚果最短,仅(17.91±0.56)cm。甘肃群体的荚果最宽,为(2.90±0.06)cm;湖南群体的荚果宽度最小,为(2.61±0.06)cm。湖北群体的荚果厚度最大,为(0.99±0.02)cm;重庆群体的荚果厚度最小,为(0.79±0.03)cm。四川群体的荚果质量最大,为(23.11±1.03)g;山东群体的荚果质量最小,为(19.63±0.85)g。
广西群体的种子最长,为(1.25±0.02)cm;而甘肃群体的种子最短,为(1.09±0.01)cm。湖北群体的种子最宽,为(0.88±0.01)cm;四川群体的种子宽度最小,为(0.80±0.01)cm。四川群体的种子厚度最大,为(0.29±0.10)cm;贵州群体的种子厚度最小,为(0.26±0.01)cm。湖北群体的种子百粒质量最大,为(48.92±1.32)g;甘肃群体的种子百粒质量最小,为(38.84±1.28)g。
实验结果表明:供试的8个皂荚天然群体间荚果特性和种子特性均存在显著差异,而且位于纬度高和降雨量低的生境中,群体的荚果和种子的多项指标均低于位于纬度低和降雨量高的生境中的群体。由于采样地受干扰程度不同,皂荚的果实和种子特性也表现出一定的差异,干扰程度相对严重的群体产生的种子相对较大。
2.2 皂荚天然群体间种子萌发特性的差异分析
在不同温度条件下8个皂荚天然群体种子萌发率的比较结果见表3。表3的数据显示:在4个温度处理条件下种皮未处理种子的萌发率在8个群体间无显著差异,平均萌发率为7.70%。而种皮经过损伤处理后种子的萌发率总体上有所提高,表明皂荚种子的种皮渗透性较差,对种皮进行破坏能够打破种子的物理性休眠、促进种子萌发。
在不同温度条件下种皮经过损伤处理后种子萌发率存在显著差异〔35 ℃ /20 ℃(F=2.81,P<0.05)、30℃ /15℃ (F=3.82,P<0.05)、25℃/10 ℃ (F=15.42,P<0.01)、15 ℃ /5 ℃(F=15.21,P<0.01)〕;其中,在昼/夜温度15℃/5℃的条件下种子萌发率低于其他处理温度,说明低温对皂荚种子的萌发有一定的抑制作用。此外,位于纬度高、降雨量低的生境中的山东和甘肃群体的种子萌发率均最低,但与其他群体间的差异并不显著。
表3 不同温度条件下8个皂荚自然群体种皮未损伤与种皮损伤种子萌发率的比较(±SD)Table 3 Comparison of germination rate of seeds with coats undamaged or damaged of eight natural populations of Gleditsia sinensis Lam.under different temperatures(±SD)
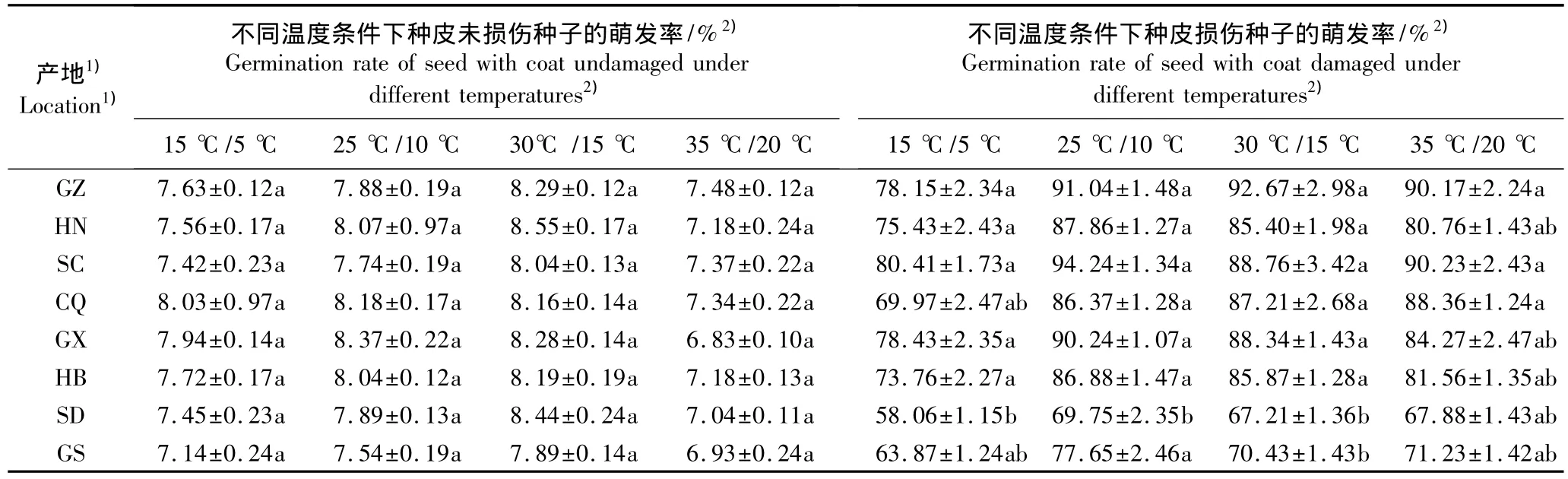
表3 不同温度条件下8个皂荚自然群体种皮未损伤与种皮损伤种子萌发率的比较(±SD)Table 3 Comparison of germination rate of seeds with coats undamaged or damaged of eight natural populations of Gleditsia sinensis Lam.under different temperatures(±SD)
1)GZ:贵州兴义Xingyi of Guizhou;HN:湖南城步Chengbu of Hu’nan;SC:四川成都Chengdu of Sichuan;CQ:重庆秀山Xiushan of Chongqing;GX:广西桂林Guilin of Guangxi;HB:湖北京山Jingshan of Hubei;SD:山东费县Feixian of Shandong;GS:甘肃天水Tianshui of Gansu.2)为昼/夜处理温度 Representing day/night treatment temperatures.同列中不同的小写字母表示差异显著(P<0.05)Different small letters in the same column indicate the significant difference(P<0.05).
?
2.3 皂荚种子特性与萌发率的相关性分析
对皂荚种子的各性状指标和不同温度条件下的萌发率进行相关性分析,结果见表4。由相关系数可见:仅在昼/夜温度30℃/15℃的条件下种子萌发率与其百粒质量有显著相关性(R=0.749,P<0.05),而在其他温度条件下种子百粒质量与种子萌发率均无显著相关性;且在不同温度条件下种子的长度、宽度和厚度与其萌发率均无显著相关性。但种子各项指标与种子萌发率均呈正相关,表明种子萌发率不仅受遗传特性限制,也与其生存环境的综合作用有关。
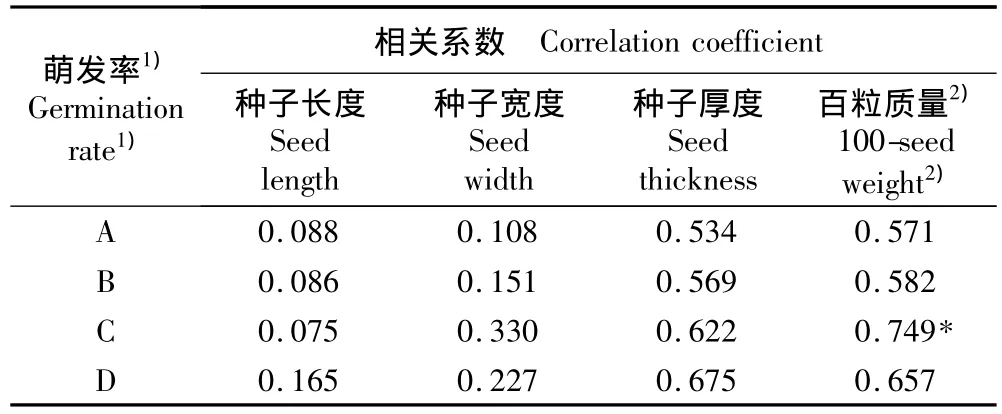
表4 不同温度条件下皂荚种子特性与种子萌发率的相关系数Table 4 Correlation coefficient between seed characteristics and seed germination rate of Gleditsia sinensis Lam.under different temperatures
3 讨论和结论
3.1 皂荚果实和种子特性与环境的关系分析
群体间种子的差异与气候、环境、经纬度、海拔高度以及植株大小等因素有关[3]。种子质量是衡量一个物种适应环境的重要指标之一,但对不同的物种影响因素有一定差异。Mazer[15]的研究结果显示:在印第安纳地区干旱对野萝卜(Raphanus raphanistrum Linn.)种子质量无影响;而 Baker[16]的研究结果表明:在加利福尼亚地区,极度干旱条件导致栎属(Quercus Linn.)和七叶树属(Aesculus Linn.)种类产生大种子。本研究结果显示:在降雨量高的地区,皂荚倾向于产生较大的果实和种子。这一结果可能与南方地区年平均降雨量和光热条件充足、植物个体获得的资源和水分较多、生殖生长较旺盛有关。此外,受人为干扰较严重的重庆和湖北皂荚群体的种子较大且质量也较高,这一结果与 Salisbury[17]的研究结果一致,因为大种子能储存更多的能量和物质,有效适应环境变化。
3.2 皂荚种子萌发特性与环境的关系分析
环境的异质性导致不同种子具有不同的萌发特性,群体间种子的萌发特性受多种因素的综合影响。Gray 等[18]和 Probert等[19]的研究结果表明:生长在高温环境中的植物种子的萌发率高于生长在低温环境中的植物种子萌发率;而 Espigares等[20]发现:昼/夜温度25℃/10℃对种子萌发有抑制作用。本研究中,在不同温度条件下,皂荚群体内种皮损伤种子的萌发率有显著差异,表现为在昼/夜温度15℃/5℃的低温条件下抑制种子萌发。这种在低温条件下种子萌发率较低的特性对皂荚适应当地气候条件有重要意义。
在种子成熟过程中,降雨量的高低会导致一些物种的种子萌发率降低或者增加。Hacker等[21]发现:分布在水分条件良好区域的群体种子萌发率显著高于分布于水分条件较差区域的群体;而Temel等[22]的研究结果表明:沿海群体的种子萌发速率高于水分条件差的内陆群体,但是内陆群体的种子萌发率则高于沿海群体。本研究结果显示:分布于低降雨量区域的山东和甘肃皂荚群体的种子萌发率较低,但与其他群体无显著差异。进一步说明在自然生境中水分条件对不同物种种子萌发的影响有所不同。
3.3 皂荚种子萌发率与种子形态的关系分析
植物种子萌发特性与其形态特征,如种子大小、形状和百粒质量等性状均有关。Grime等[23]认为:种子越小越容易萌发;而Leishman等[24]认为:对同一种类而言,大种子比小种子具有更高的萌发率,在资源受到限制的条件下大种子的萌发和幼苗更新比小种子有更大的优势。而对皂荚种子特性与萌发率的相关性分析结果也表明:相同条件下皂荚的大种子比小种子具有更高的萌发率。表明群体间种子萌发率的差异可能是由种子大小决定的,也即环境的差异使种子产生大小差异,形成了不同的萌发特性。
3.4 皂荚种子休眠与环境的关系分析
群体间种子休眠特性受经纬度、海拔、生境条件、光照、温度、水分以及受干扰程度等因素的影响。在18℃、21℃和24℃条件下Syringa reflexa Schneid种子的休眠率比生长在15℃条件下更高[25];水分胁迫降低了 Avena fatua Linn.种子的生理休眠[26];但水分胁迫可增加无翅猪毛菜(Salsola komarovii Iljin)的生理休眠[27]。种子的物理休眠能保证种子在极端环境条件下存活和产生持久种子库[28]。皂荚种子具有物理性休眠特性,能够在胁迫环境条件下保存种子,对适应不同地区的生长环境有重要意义。通过人为损伤种皮,皂荚种子萌发率可显著提高。但种皮损伤只能打破物理休眠,而在4℃密封条件下保存2 a的皂荚种子仍然具有95%的活力,因此,皂荚种子可能具有复合型休眠类型,种皮损伤只能打破其物理休眠,但不能解除其生理休眠。这也可能是皂荚种子对环境的适应策略之一。
综上所述,为适应不同环境条件,不同皂荚群体的荚果和种子特性广泛变异,且在降雨量高的地区荚果和种子均较大,但群体间种子萌发率无显著差异;种皮损伤能打破皂荚种子的物理休眠,且低温可抑制其种子萌发;皂荚种子还可能存在生理性休眠。可见,不同程度的种子复合休眠是皂荚适应不同生境的生存策略之一,可保证其在不同生境中更新、存活和繁衍。
[1]LORD J,WESTOBY M,LEISHMAN M.Seed size and phylogeny in six temperate floras:constraints,niche conservatism,and adaptation[J].The American Naturalist,1995,146(3):349-364.
[2]KIDSON R,WESTOBY M.Seed mass and seedling dimensions in relation to seedling establishment[J].Oecologia,2000,125(1):11-17.
[3]MICHAELS H J,BENNER B,HARTGERINK A P,et al.Seed size variation:magnitude,distribution,and ecological correlates[J].Evolutionary Ecology,1988,2(2):157-166.
[4]SUSKO D J,LOVETT-DOUST L.Patterns of seed mass variation and their effects on seedling traits in Alliaria petiolata[J].American Journal of Botany,2000,87(1):56-66.
[5]MURRAY B R,BROWN A H D,DICKMAN C R,et al.Geographical gradients in seed mass in relation to climate[J].Journal of Biogeography,2004,31(3):379-388.
[6]MEYER S E,KITCHEN S G,CARLSON S L.Seed germination timing patterns in intermountain Penstemon(Scrophulariaceae)[J].American Journal of Botany,1995,82(3):377-389.
[7]ANDERSSON L,MILBERG P.Variation in seed dormancy among mother plants,populations and years of seed collection[J].Seed Science Research,1998,8(1):29-38.
[8]THOMPSON K,OOI M K J.To germinate or not to germinate:more than just a question of dormancy[J].Seed Science Research,2010,20(4):209-211.
[9]冯 蕾,白志英,路丙社,等.氯化钠胁迫对枳椇和皂荚生长,叶绿素荧光及活性氧代谢的影响[J].应用生态学报,2008,19(11):2503-2508.
[10]WU J,LI J,ZHU Z,et al.Protective effects of echinocystic acid isolated from Gleditsia sinensis Lam.against acute myocardial ischemia[J].Fitoterapia,2010,81(1):8-10.
[11]王蓟花,唐 静,李 瑞,等.皂荚化学成分和生物活性的研究进展[J].中国野生植物资源,2008,27(6):1-3.
[12]LAI P,DU J R,ZHANG M X,et al.Aqueous extract of Gleditsia sinensis Lam.fruits improves serum and liver lipid profiles and attenuates atherosclerosis in rabbits fed a high-fat diet[J].Journal of Ethnopharmacology,2011,137(3):1061-1066.
[13]顾万春,李 斌,孙翠玲.皂荚优良产地和优良种质荐[J].林业科技通讯,2001(4):10-13.
[14]兰彦平,顾万春.北方地区皂荚种子及荚果形态特征的地理变异[J].林业科学,2006,42(7):47-51.
[15]MAZER S J.Ecological,taxonomic,and life history correlates of seed mass among Indiana dune angiosperms[J].Ecological Monographs,1989,59(2):153-175.
[16]BAKER H G.Seed weight in relation to environmental conditions in California[J].Ecology,1972,53(6):997-1010.
[17]SALISBURY E.Seed size and mass in relation to environment[J].Proceedings of the Royal Society of London(Series B):Biological Sciences,1974,186:83-88.
[18]GRAY D,STECKEL J R A.Viability of onion(Allium cega)seed as influenced by temperature during seed growth[J].Annals of Applied Biology,1984,104(2):375-382.
[19]PROBERT R J,SMITH R D,BIRCH P.Germination responses to light and alternating temperatures in European populations of Dactylis glomerata[J].New Phytologist,1985,99(2):317-322.
[20]ESPIGARES T,PECO B.Mediterranean pasture dynamics:the role of germination[J].Journal of Vegetation Science,1993,4(2):189-194.
[21]HACKER J B,RATCLIFF D.Seed dormancy and factors controllingdormancybreakdown in buffelgrassaccession from contrasting provenances[J].Journal of Applied Ecology,1989,26(1):201-212.
[22]TEMEL F,GÜLCÜ S,ÖLMEZ Z,et al.Germination of Anatolian Black Pine(Pinus nigre subsp.pallasiana)seeds from the Lakes Region of Turkey:geographic variation and effect of storage[J].Notulae Botanicae Horti Agrobotanici Cluj-Napoca,2011,39(1):267-274.
[23]GRIME J,MASON G,CURTIS A V,et al.A comparative study of germination characteristics in a local flora[J].Journal of Ecology,1981,69(3):1017-1059.
[24]LEISHMAN M R,WESTOBY M.Hypotheses on seed size:tests using the semiarid flora of western New South Wales,Australia[J].The American Naturalist,1994,143(5):890-906.
[25]JUNTTILA O.Effects of mother plant temperature on seed development and germination in Syringa reflexa Schneid[J].Meldinger fra Norges Landbrukshoeqskole,1971,50(10):1-16.
[26]SAWHNEY R,NAYLOR J M.Dormancy studies in seed of Avena fatua.13.Influence of drought stress during seed development on duration of seed dormancy[J].Canadian Journal of Botany,1982,60(6):1016-1020.
[27]YAMAGUCHI H,ICHIHARA K,TAKENO K,et al.Diversities in morphological characteristics and seed germination behavior in fruits of Salsola komarovii Iljin[J].The Botanical Magazine,1990,103(2):177-190.
[28]TRAN V N,CAVANAGH A K.Structural aspects of dormancy[M]∥ MURRAY D R.Seed Physiology:Vol.2.Sydney:Academic Press Australia,1984:1-44.
