西藏高原高寒草原生态系统植被C/N值的分布特征及其影响因素
2013-12-14王建林钟志明王忠红张宪洲沈振西胡兴祥
王建林,钟志明,王忠红,张宪洲,沈振西,胡兴祥
(1.西藏农牧学院植物科学学院,西藏林芝860000;2.中国科学院地理科学与资源研究所,北京100101)
作为植物的基本营养元素,碳(C)和氮(N)在植物生长和生理过程中发挥着重要作用,且彼此关系密切[1-4]。其中,C是构成植物体内干物质的最主要元素,而N则与植物的光合作用和细胞生长分裂等重要生理活动有关[4]。作为重要的生理指标,C/N比反映了植物的生长速率[5-6],并与植物对N的利用效率有关[7]。近年来,生态化学计量学的发展为研究上述问题提供了有力工具,并强调活有机体主要组成元素(特别是C、N)的关系。应用化学计量学方法研究C/N的区域分布规律已成为当前的研究热点之一[8]。
西藏高原是地球上面积最大、海拔最高的高原,平均海拔在4 000 m以上,被誉为地球“第三极”。西藏高原地势高亢、空气稀薄、紫外线辐射强度大,对亚洲甚至北半球的大气环流和碳平衡等均有重要影响,使得高原植被和土壤对气候变化极为敏感,因此西藏高原也被称为全球气候变化的敏感区,也使西藏高原高寒草原生态系统在全球气候变化研究中占有特殊地位,为研究不同地理气候条件下生态系统结构和功能提供了天然“实验室”。所以该区域一直是全球相关研究者关注的热点地区。
目前,围绕西藏高原土壤的碳储量[9]、CO2排放[10]、有机质周转[11]以及氮循环[12]等方面开展了大量的研究工作,但目前涉及整个高原跨不同植被带(不同自然地带)高寒草原生态系统植被C/N分布的研究报道很少。为此,作者通过对西藏高原高寒草原生态系统植被C/N值的研究,揭示该区域植被C/N值的空间分布特征及其影响因素,以期为研究西藏高原对气候变化响应的区域差异提供科学依据。
1 研究区概况和研究方法
1.1 研究区概况
研究区域为西藏黑阿(那曲—阿里)、青藏(拉萨—青海)、新藏(拉萨—新疆)公路沿线没有受人类活动干扰的高寒草原区。研究区年均温-4℃~6℃,年均降水量50~500 mm,跨越高山灌丛草甸、山地灌丛草原、山地半荒漠与荒漠以及高山草原4个自然地带。植被包含昆仑针茅(Stipa roborowskyi Roshev.)-窄叶薹草(Carex montis-everestii Kǖkenth.)、紫花针茅(Stipa purpurea Griseb.)+ 藏 沙 蒿 (Artemisia wellbyi Hemsl.et Pears.)、紫花针茅+矮火绒草〔Leontopodium nanum(Hook.f.et Thoms.ex C.B.Clarke)Hand.-Mazz.〕、青藏薹草(Carex moorcroftii Falc.ex Boott)、紫花针茅+干生薹草(Carex aridula V.Krecz.)、固沙草〔Orinus thoroldii(Stapf ex Hemsl.)Bor〕+劲直黃蓍(Astragalus strictus Graham ex Bentham)、紫花针茅+杂类草、青藏薹草+紫花针茅、紫花针茅、固沙草-藏薹草(Carex thibetica Franch.)-紫花针茅、藏东蒿(Artemisia vexans Pamp.)、紫花针茅+藏北蒿(Kobresia littledalei C.B.Clarke)、拉 萨 小 檗 (Berberishemsleyana Ahrendt)-紫花针茅+白草(Pennisetum centrasiaticum Tzvel.)、羽柱针茅〔Stipa subsessiliflora var.basiplumosa(Munro ex Hook.f.)P.C.Kuo et Y.H.Sun〕-紫花针茅-禾草、紫花针茅+沙生针茅(Stipa glareosa P.Smirn.)、紫花针茅-矮生嵩草〔Kobresia humilis(C.A.Mey.ex Trautv.)Sergiev〕-藏薹草、金露梅(Potentilla fruticosa Linn.)-青藏薹草+紫花针茅、紫花针茅+羽柱针茅和紫花针茅+固沙草等19个高寒草原草地型[13],其中:紫花针茅+干生薹草、固沙草-藏薹草-紫花针茅、羽柱针茅-紫花针茅-禾草、固沙草+劲直黃芪、藏东蒿、昆仑针茅-窄叶薹草、金露梅-青藏薹草+紫花针茅、紫花针茅-矮生嵩草-藏薹草以及拉萨小檗-紫花针茅+白草9个草地型属于山地灌丛草原带,紫花针茅+杂类草以及青藏薹草+紫花针茅2个草地型属于高山灌丛草甸带,紫花针茅+羽柱针茅、紫花针茅+藏北蒿、紫花针茅+藏沙蒿、紫花针茅+沙生针茅以及紫花针茅+固沙草5个草地型属于山地半荒漠与荒漠带,青藏薹草、紫花针茅以及紫花针茅+矮火绒草3个草地型属于高山草原带。
1.2 研究方法
1.2.1 样点设置与样品采集 于2011年7至8月间,沿黑阿、青藏和新藏公路沿线设置54个采样点,所有采样点的地理位置如下:1)隆子县扎西康(N28.23°,E91.95°);2)康 马 县 嘎 拉 (N28.32°,E89.48°);3)聂拉木县吉隆(N28.47°,E86.15°);4)聂拉木县乃隆 (N28.64°,E86.44°);5)定日县白坝(N28.60°,E87.08°);6)曲松县邛多江(N28.77°,E90.12°);7)浪卡子县白地(N29.01°,E90.40°);8)羊卓雍湖畔(N29.18°,E90.55°);9)羊卓雍湖畔(N29.18°,E90.58°);10)羊卓雍湖畔(N29.18°,E90.60°);11)羊卓雍湖畔 (N29.18°,E90.60°);12)羊卓雍湖畔 (N29.18°,E90.80°);13)萨嘎县麦岗(N29.32°,E85.27°);14)墨竹工县日多 (N29.76°,E92.37°);15)昂仁县桑桑 (N29.43°,E86.70°);16)昂仁县尼日勒(N29.80°,E85.75°);17)林周县松盘(N29.98°,E91.27°);18)仲巴县帕羊(N30.10°,E83.47°);19)当雄县羊八井(N30.18°,E90.60°);20)措勤县塔丝 (N30.28°,E85.30°);21)措勤县郊(N31.10°,E84.98°);22)当 雄 县 城 (N30.50°,E91.01°);23)当雄县城(N30.50°,E91.06°);24)当雄县城(N30.51°,E91.07°);25)普兰马攸木拉山(N30.60°,E80.53°);26)普 兰 拉 昂 错 (N30.68°,E81.30°);27)普兰马攸木拉山(N30.95°,E82.25°);28)纳 木 措 (N30.70°,E90.88°);29)纳 木 措(N30.75°,E91.07°):30)噶 尔 县 门 士 (N31.37°,E80.53°);31)噶尔县莫落江(N31.45°,E80.52°);32)噶尔县城(N32.56°,E80.03°);33)班戈县舍库(N31.12°,E90.57°);34)班 戈 县 城 (N31.40°,E90.05°);35)班戈县城(N31.40°,E90.05°);36)班戈县城(N31.40°,E91.05°);37)班戈县城(N31.42°,E90.00°);38)班戈县城(N31.42°,E90.00°);39)那曲县错鄂湖畔(N31.51°,E91.36°);40)安多县帮爱乡 (N31.67°,E91.83°);41)札达县阿依木拉山(N31.87°,E80.08°);42)札达县阿依木拉山(N31.88°,E80.08°);43)改则县洞措(N32.08°,E84.80°);44)改则县洞措(N32.10°,E84.78°);45)革吉县小盐湖(N32.23°,E82.23°);46)革吉县帮巴(N32.28°,E81.23°);47)革吉县盐湖(N32.52°,E82.45°);48)日土县班公措(N33.43°,E79.77°);49)日土县班公措(N33.43°,E79.77°);50)班戈县屋刻骇 (N31.47°,E89.82°);51)那 曲县那莫 切(N34.57°,E91.57°);52)岗巴县东鸡山(N28.73°,E88.75°);53)浪卡子县工布学(N28.63°,E91.07°);54)措美县哲古乡(N28.53°,E91.63°)。其中,1 ~18、52、53和54 号采样点属于山地灌丛草原,19、22、23、24、39和40 号采样点属于高山灌丛草甸,20、21、28、29、33 ~38、43 ~45、47、50 和 51 号采样点属于高山草原,25 ~27、30 ~32、41、42、46、48 和 49 号采样点属于山地半荒漠与荒漠。
在每个采样点设置面积1 m×1 m的样方6个,先在每个样方中用收获法收集植物的地上部分,再在该样方中随机设1个面积25 cm×25 cm的小样方挖取土柱,土柱深度40 cm,并按10 cm间隔分层,将分层土柱装入80目尼龙袋中,将泥沙冲洗干净后用镊子拣取所有根系装入布袋中;同时,在该样方中再设置另1个面积25 cm×25 cm的小样方,挖取土壤剖面,用机械采样法分别采集0~10、10~20、20~30和30~40 cm土层的土壤样品;同时调查每个样方中的植物种数、优势种、平均高度和平均盖度,并用海拔仪测定海拔。另外,收集各样点相邻气象站的年均气温与年均降水量等气象资料。
1.2.2 测定方法 将采集的植物地上部分和根系样品置于80℃的恒温箱中干燥至质量恒定,分别采用半微 量 凯 氏 法[14]375-376、重 铬 酸 钾 氧 化 - 外 加 热法[14]376-377和双指示剂滴定法[14]399测定植物地上部分(叶和茎)以及0~10、10~20、20~30和30~40 cm土层中植物根系的全N、有机C和无机C含量。
采用酒精燃烧法测定0~10、10~20、20~30和30~40 cm土层的土壤含水量,用环刀法同步测定土壤容重。所有土壤样品风干后磨细并过1 mm筛,用于相关土壤化学指标测定。其中,土壤全N含量采用半微量凯氏法[14]375-376测定,速效 N 含量采用蒸馏法[14]78-82测定,全 P 含量采用钼锑抗比色法[14]97-101测定,速效 P 含量采用碳酸氢钠法[14]105-108测定,全 K和速效K含量采用原子吸收分光光度法[14]113-115测定,土壤有机质、总有机C、总碳水化合物和活性C含量均采用重铬酸钾氧化-外加热法[14]376-377测定,土壤pH值采用电位法[14]146-156测定,水解性C和水溶性C含量采用蒽酮法[14]132-136测定,总无机 C含量用双指示剂滴定法[14]399测定。
1.3 数据处理
采用逆距离权重法(inverse distance weighting,IDW)法对所有样点的植物地上部分和根系C/N值(为全C和全N的含量比值)的平均数据作空间分布图。作图时,设平面上分布一系列离散点,已知其坐标和值为 Xi,Yi,Zi(i=1,2,……,n),通过距离加权值求 Z 点值公式[15]:
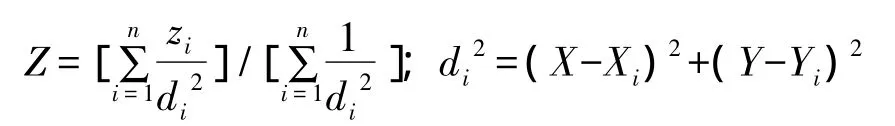
式中,Z为待估计的植被C/N栅格值,zi为第i(i=1,2,……,n)个采样点的植被C/N值,n为用于植被C/N插值的采样点个数,di为插值点到第i个采样点的距离。
同时,采用单因素方差分析和最小显著差异法比较所有样点植物地上部分和根系C/N平均值在不同数据组间的差异。此外,因土壤理化性质、气象因子、植被因子和地理属性对高寒草原土壤及植被C和N含量均有明显影响,因而,用逐步回归分析法分别建立所有样点植物地上部分和根系C/N值与地理、气候、植被、土壤物理和土壤化学因子之间的相关方程。
2 结果和分析
2.1 植被C/N值的分布特征
2.1.1 植被C/N值的水平分布格局 西藏高原高寒草原植被C/N值的水平分布格局见图1。西藏高原高寒草原植被C/N值的水平分布有2个特点:一是整体呈现出中间低而东西部高的态势;二是呈现出斑块状交错分布的格局。植被C/N值的高值区主要集中在以扎日南木错和洞措为核心的藏北高原腹地,而在马攸木拉山以西则为植被C/N值的低值区。
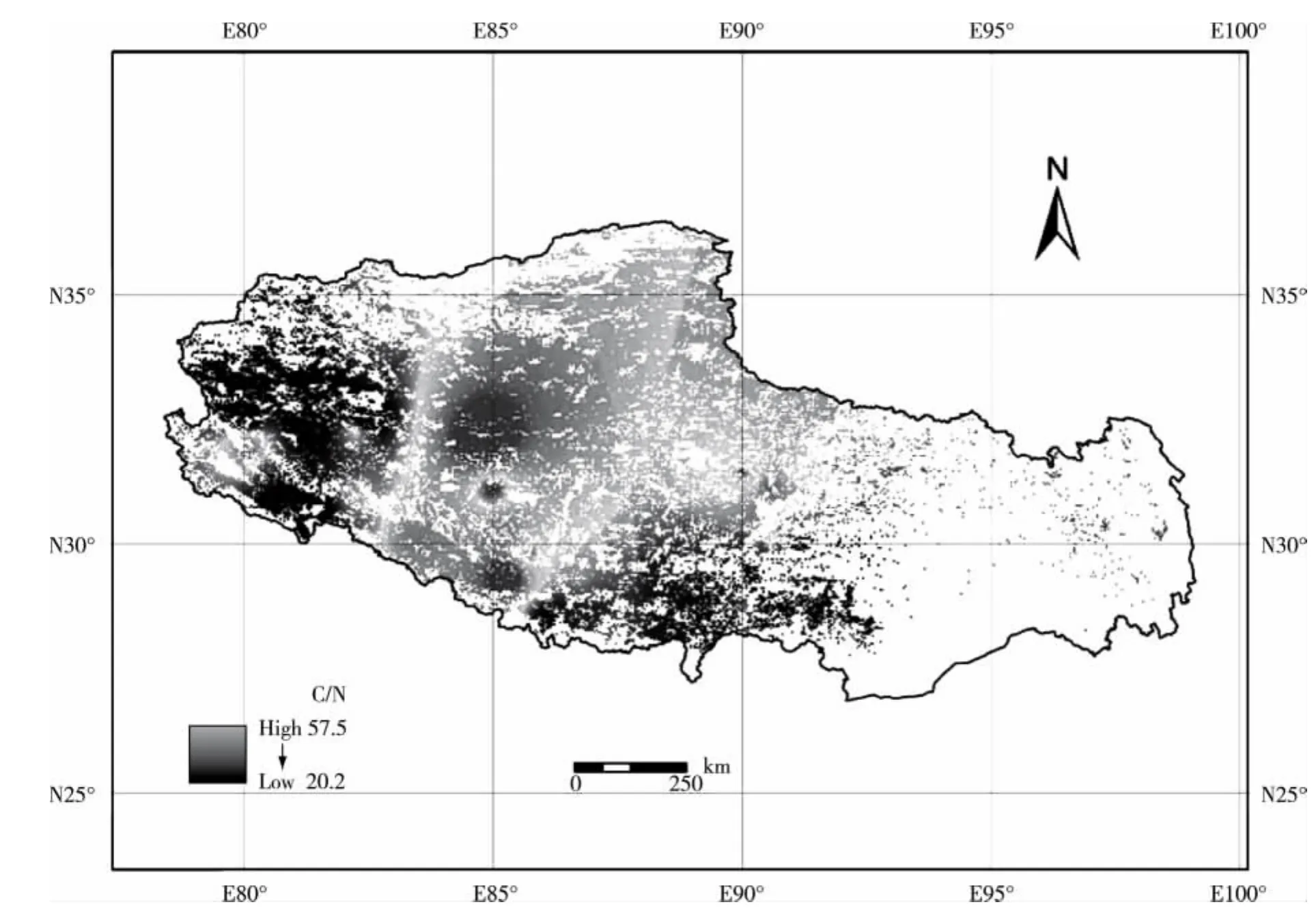
图1 西藏高原高寒草原不同草地型植被C/N值的水平分布格局Fig.1 Horizontal distribution pattern of C/N value in vegetation of different grassland types of alpine steppe in Tibetan Plateau
表1 西藏高原高寒草原不同草地型植被地上部分和根系的C/N值比较(±SE)Table 1 Comparison of C/N value in above-ground part and root of vegetation among different grassland types of alpine steppe in Tibetan Plateau(±SE)
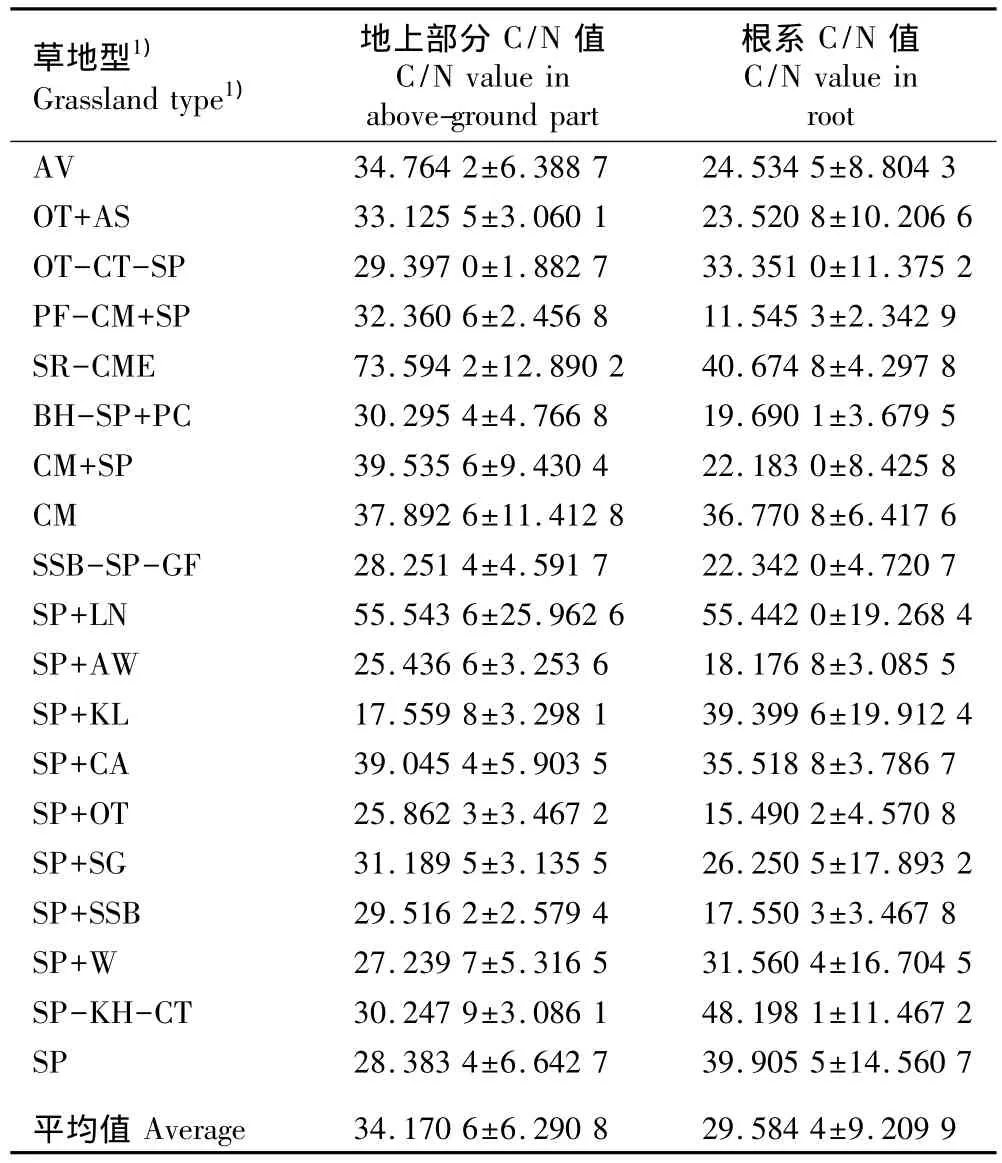
表1 西藏高原高寒草原不同草地型植被地上部分和根系的C/N值比较(±SE)Table 1 Comparison of C/N value in above-ground part and root of vegetation among different grassland types of alpine steppe in Tibetan Plateau(±SE)
1)AV:藏东蒿 Artemisia vexans Pamp.;OT:固沙草 Orinus thoroldii(Stapf ex Hemsl.)Bor;AS:劲直黃蓍 Astragalus strictus Graham ex Bentham;CT:藏薹草 Carex thibetica Franch.;SP:紫花针茅 Stipa purpurea Griseb.;PF:金露梅 Potentilla fruticosa Linn.;CM:青藏薹草 Carex moorcroftii Falc.ex Boott;SR:昆仑针茅 Stipa roborowskyi Roshev.;CME:窄叶薹草 Carex montis-everestii Kǖkenth.;BH:拉萨小 檗 BerberishemsleyanaAhrendt;PC:白 草 Pennisetum centrasiaticum Tzvel.;SSB: 羽 柱 针 茅 Stipa subsessiliflora var.basiplumosa(Munro ex Hook.f.)P.C.Kuo et Y.H.Sun;GF:禾草Gramineous forage grasses;LN:矮火绒草 Leontopodium nanum(Hook.f.et Thoms.ex C.B.Clarke)Hand.-Mazz.;AW:藏沙蒿Artemisia wellbyi Hemsl.et Pears.;KL:藏北蒿 Kobresia littledalei C.B.Clarke;CA:干生薹草 Carex aridula V.Krecz.;SG:沙生针茅Stipa glareosa P.Smirn.;W:杂类草 Weeds;KH:矮生嵩草Kobresia humilis(C.A.Mey.ex Trautv.)Sergiev.
?
2.1.2 不同草地型植被C/N值的分布特征 西藏高原高寒草原不同草地型植被地上部分和根系的C/N值见表1。由表1可知:植物地上部分C/N平均值为34.17,变异系数达到35.87%。从昆仑针茅-窄叶薹草、紫花针茅+矮火绒草、青藏薹草+紫花针茅、紫花针茅+干生薹草、青藏薹草以及藏东蒿草地型,到固沙草+劲直黃芪、金露梅-青藏薹草+紫花针茅、紫花针茅+沙生针茅、拉萨小檗-紫花针茅+白草、紫花针茅-矮生嵩草-藏薹草以及紫花针茅+羽柱针茅草地型,再到固沙草-藏薹草-紫花针茅、紫花针茅、羽柱针茅-紫花针茅-禾草、紫花针茅+杂类草、紫花针茅+固沙草、紫花针茅+藏沙蒿草以及紫花针茅+藏北蒿草地型,植物叶片C/N值逐渐下降,最大者(昆仑针茅-窄叶薹草草地型)几乎是最小者(紫花针茅+藏北蒿草地型)的4倍以上。
由表1还可以看出:19个草地型植被根系的C/N平均值为29.58,变异系数达到40.02%。从紫花针茅+矮火绒草、紫花针茅-矮生嵩草-藏薹草、昆仑针茅-窄叶薹草、紫花针茅、紫花针茅+藏北蒿、青藏薹草草地型,到紫花针茅+干生薹草、固沙草-藏薹草-紫花针茅、紫花针茅+杂类草、紫花针茅+沙生针茅、藏东蒿、固沙草+劲直黃芪、羽柱针茅-紫花针茅-禾草草地型,再到青藏薹草+紫花针茅、拉萨小檗-紫花针茅+白草、紫花针茅+藏沙蒿、紫花针茅+羽柱针茅、紫花针茅+固沙草和金露梅-青藏薹草+紫花针茅草地型,根系的C/N值逐渐下降,最大者(紫花针茅+矮火绒草草地型)也几乎是最小者(金露梅-青藏薹草+紫花针茅草地型)的4倍以上。结果表明:西藏高原高寒草原植被无论是植物地上部分C/N值还是植物根系C/N值,不同草地型之间均有不同程度的差异;此外,植物地上部分C/N值均大于根系的C/N值,但其变异幅度小于后者。
2.1.3 不同自然地带植被C/N值的分布特征 西藏高原高寒草原不同自然地带植被地上部分和根系的C/N值见表2。由表2可见:在西藏高原不同自然地带中植被C/N值的水平分布也存在差异。不同自然地带植物地上部分的C/N平均值为31.98,变异系数仅为13.82%;4种自然地带按植物地上部分C/N值由大至小依次排序为山地灌丛草原带、高山灌丛草甸带、高山草原带、山地半荒漠与荒漠带;其中,山地灌丛草原带植物地上部分 C/N值最大,为34.65;而山地半荒漠与荒漠带植物地上部分C/N值最小,为25.37,仅相当于前者的73.23%。由表2还可见:不同自然地带植物根系的C/N平均值为31.86,变异系数为16.92%;4种自然地带按植物根系C/N值由大至小依次排序为高山草原带、山地半荒漠与荒漠带、山地灌丛草原带、高山灌丛草甸带;其中,高山草原带植物根系C/N值最大,为39.42;而高山灌丛草甸带植物根系C/N值最小,为27.83,仅相当于前者的70.61%。结果表明:无论是植物地上部分还是根系的C/N值,不同自然地带间均存在显著差异;此外,不同自然地带植物地上部分C/N值均大于根系C/N值,但其变异幅度却小于后者。
2.2 植被C/N值与环境因子的关系
2.2.1 植被C/N值与地理因子的关系 西藏高原高寒草原植被地上部分C/N值与地理因子的回归方程为 Y=73.302 0- 1.047 8X1+0.138 1X2-0.004 5X3(N=54,R=0.184 5,R1=-0.114 7,R2=0.049 7,R3=-0.095 2);植物根系C/N值与地理因子的回归方程为 Y=-66.408 4+1.302 9X1- 0.269 0X2+0.017 7X3(N=54,R=0.321 7,R1=0.109 1,R2=-0.073 9,R3=0.277 1)。从回归方程可以看出:西藏高原高寒草原植被植物地上部分C/N值与纬度(X1)和海拔(X3)呈负相关关系、与经度(X2)呈正相关关系;植物根系C/N值与纬度(X1)和海拔(X3)呈正相关关系、与经度(X2)呈负相关关系。根据标准误差检验,这2个回归方程均未通过0.05水平的显著性检验,但植物根系C/N值与海拔(X3)的偏相关系数(R3=0.277 1)达到显著水平(P<0.05)。表明对西藏高原高寒草原植被植物地上部分C/N值影响最大的地理因子是纬度,而对植物根系C/N值影响最大的因子是海拔。此外,地理因子对西藏高原高寒草原生态系统植被C/N值的影响整体不显著。
表2 西藏高原高寒草原不同自然地带植被地上部分和根系的C/N值比较(±SE)Table 2 Comparison of C/N value in above-ground part and root of vegetation among different natural transects of alpine steppe in Tibetan Plateau(±SE)

表2 西藏高原高寒草原不同自然地带植被地上部分和根系的C/N值比较(±SE)Table 2 Comparison of C/N value in above-ground part and root of vegetation among different natural transects of alpine steppe in Tibetan Plateau(±SE)
?
2.2.2 植被C/N值与气候因子的关系 西藏高原高寒草原植被地上部分C/N值与气候因子的回归方程为 Y=16.139 6+0.005 5X7+0.416 1X9-0.010 5X10(N=54,R=0.354 1,R7=0.179 3,R9=0.239 2,R10=-0.260 7)。从回归方程可看出:影响西藏高原高寒草原植被植物地上部分C/N值的主要气候因子是≥10℃积温(X7)、6月至9月降水率(X9)和年均蒸发量(X10),而年均日照时数(X1)、年均气温(X2)、最冷月均气温(X3)、最暖月均气温(X4)、≥0℃积温(X5)、≥5℃积温(X6)、年均降水量(X8)、年均相对湿度(X11)的影响则很小。此外,植物地上部分C/N值与≥10℃积温和6月至9月降水率呈正相关关系、与年均蒸发量呈负相关关系。根据标准误差检验,植物地上部分C/N值与气候因子的相关方程未通过0.05水平的显著性检验。表明西藏高原高寒草原植被地上部分C/N值随≥10℃积温和6月至9月降水率的增加以及年均蒸发量的减少而增大,也表明气候因子对植物地上部分C/N值的影响未达到显著水平。
西藏高原高寒草原植被根系C/N值与气候因子的回归方程为 Y=76.161 1+7.425 4X4+0.016 3X5-0.020 1X7-0.171 7X8+0.7694 8X9-0.072 2X10(N=54,R=0.470 9,R4=0.248 7,R5=0.280 4,R7=-0.219 0,R8=-0.377 0,R9=0.232 9,R10=-0.295 1)。从回归方程可看出:影响西藏高原高寒草原植被植物根系C/N值的主要气候因子是最暖月均气温(X4)、≥0℃积温(X5)、≥10℃积温(X7)、年均降水量(X8)、6月至9月降水率(X9)和年均蒸发量(X10),而年均日照时数(X1)、年均气温(X2)、最冷月均气温(X3)、≥5℃积温(X6)和年均相对湿度(X11)的影响则很小。植物根系C/N值与年均降水量和年均蒸发量的偏相关系数(R8=-0.377 0,R10=-0.295 1)达到极显著(P<0.01)或显著(P<0.05)水平,而与最暖月均气温、≥0℃积温、≥10℃积温和6月至9月降水率的偏相关系数未达到显著水平。根据标准误差检验,植物根系C/N值与气候因子的回归方程未通过0.01水平的显著性检验。表明影响西藏高原高寒草原植被植物根系C/N值的主要气候因子是最暖月均气温、≥0℃积温、≥10℃积温、年均降水量、6月至9月降水率和年均蒸发量;其中,年均降水量和年均蒸发量的影响尤为显著,植物根系C/N值随年均降水量和年均蒸发量的减少而显著增大。此外,气候因子对植物根系C/N值的影响未达到极显著水平,而且影响植物地上部分与根系C/N值的气候因子各不相同。
2.2.3 植被C/N值与植被因子的关系 西藏高原高寒草原植被植物地上部分C/N值与植被因子的回归方程为 Y=33.299 3+0.034 0X5-0.232 1X7(N=54,R=0.650 5,R5=0.312 4,R7=-0.325 2)。从方程可以看出:影响西藏高原高寒草原植被植物地上部分C/N值的主要植被因子是地上部生物量(X5)和植被高度(X7),而0~10 cm地下部生物量(X1)、10~20 cm地下部生物量(X2)、20~30 cm地下部生物量(X3)、30~40 cm地下部生物量(X4)和植被盖度(X6)等植被因子则对地上部分C/N值无明显影响。此外,植物地上部分C/N值与地上部生物量和植被高度的偏相关系数(R5=0.312 4,R7=-0.325 2)均达到显著水平(P<0.05)。根据标准误差检验,西藏高原高寒草原植被植物地上部分C/N值与植被因子的回归方程通过了0.01水平的显著性检验。表明影响西藏高原高寒草原植被植物地上部分C/N值的主要植被因子是地上部生物量和植被高度,地上部分C/N值随地上部生物量的增加和植被高度的降低而显著增大;此外,植被因子对植物地上部分C/N值的影响达到极显著水平。
西藏高原高寒草原植被根系C/N值与植被因子的回归方程为 Y=33.762 0-0.003 1X1+0.029 7X6(N=54,R=0.339 8,R1=-0.277 8,R6=0.199 1)。从方程中可看出:影响西藏高原高寒草原植被植物根系C/N值的主要植被因子是0~10 cm地下部生物量(X1)和植被盖度(X6),而10~20 cm地下部生物量(X2)、20~30 cm地下部生物量(X3)、30~40 cm地下部生物量(X4)、地上部生物量(X5)和植被高度(X7)等植被因子则对植物根系C/N值无明显影响。此外,植物根系C/N值与0~10 cm地下部生物量的偏相关系数(R1=-0.277 8)达到显著水平(P<0.05)。根据标准误差检验,西藏高原高寒草原植被植物根系C/N值与植被因子的回归方程通过了0.01水平的显著性检验。表明影响西藏高原高寒草原植被植物根系C/N值的主要植被因子是0~10 cm地下部生物量和植被盖度,植物根系C/N值随0~10 cm地下部生物量的减少而显著增大;此外,植被因子对植物根系C/N值的影响达到极显著水平,但影响植物地上部分与根系C/N值的植被因子各不相同。
2.2.4 植被C/N值与土壤物理因子的关系 西藏高原高寒草原植被地上部分C/N值与土壤物理因子的回归方程为 Y=45.919 0+28.780 0X1-16.661 7X2-21.964 7X3+0.839 9X5-0.877 4X6- 0.075 5X8(N=54,R=0.290 7,R1=0.112 8,R2= -0.060 0 ,R3=-0.239 4,R5=0.208 6,R6=-0.200 0,R8=-0.035 0)。从方程中可看出:影响西藏高原高寒草原植被植物地上部分C/N值的主要土壤物理因子是0~10 cm土壤容重(X1)、10~20 cm土壤容重(X2)、20~30 cm土壤容重(X3)、0~10 cm 土壤含水量(X5)、10~20 cm土壤含水量(X6)和30~40 cm土壤含水量(X8),而30~40 cm土壤容重(X4)和20~30 cm土壤含水量(X7)则对植物地上部分C/N值无明显影响。根据标准误差检验,植物地上部分C/N与土壤物理因子的回归方程未通过0.05水平的显著性检验。表明影响西藏高原高寒草原植被植物地上部分C/N值的主要土壤物理因子是0~10 cm土壤容重、10~20 cm土壤容重、20~30 cm土壤容重、0~10 cm土壤含水量、10~20 cm土壤含水量和30~40 cm土壤含水量,随0~10 cm土壤容重和0~10 cm土壤含水量的增加以及10~20 cm土壤容重、20~30 cm土壤容重、10~20 cm土壤含水量和30~40 cm土壤含水量的降低,植物地上部分C/N值增大;此外,土壤物理因子对植物地上部分C/N值的影响未达到显著水平。
西藏高原高寒草原植被根系C/N值与土壤物理因子的回归方程为 Y=-3.856 0+28.507 0X3(N=54,R=0.320 6)。从方程中可看出:影响西藏高原高寒草原植被植物根系C/N值的主要土壤物理因子是20~30 cm土壤容重(X3),而0~10 cm土壤容重(X1)、10~20 cm土壤容重(X2)、30~40 cm土壤容重(X4)、0~10 cm土壤含水量(X5)、10~20 cm 土壤含水量(X6)、20~30 cm土壤含水量(X7)和30~40 cm土壤含水量(X8)等土壤物理因子则对植物根系C/N值无明显影响。根据标准误差检验,植物根系C/N值与土壤物理因子的回归方程通过了0.05的显著性检验。表明影响西藏高原高寒草原植被植物根系C/N值的主要土壤物理因子是20~30 cm土壤容重,随20~30 cm土壤容重的增加植物根系C/N值显著增大;此外,土壤物理因子对植物根系C/N值的影响达到显著水平,但影响植物地上部分与植物根系C/N值的土壤物理因子各不相同。
2.2.5 植被C/N值与土壤化学因子的关系 西藏高原高寒草原植被地上部分C/N值与土壤化学因子的回归方程为 Y=31.677 0-0.013 4X2-76.019 9X5-10.770 5X7-2 981 583.002 2X8+5 140 260.734 8X9-1.504 6X13(N=54,R=0.567 1,R2=-0.170 7,R5=-0.424 3,R7=-0.345 4,R8=-0.169 4,R9=0.169 4,R13=-0.142 5)。从方程中可看出:影响西藏高原高寒草原植被植物地上部分C/N值的主要土壤化学因子是土壤速效N含量(X2)、全N含量(X5)、全K含量(X7)、有机质含量(X8)、总有机C含量(X9)和活性C含量(X13),而土壤pH值(X1)、速效P含量(X3)、速效K含量(X4)、全P含量(X6)、总碳水化合物含量(X10)、水解性C含量(X11)、水溶性 C含量(X12)和含量(X14)等土壤化学因子则对植物地上部分C/N值无明显影响。此外,植物地上部分C/N值与全N含量和全K含量的偏相关系数(R5=-0.424 3,R7=-0.345 4)达 到 极 显 著(P<0.01)或 显 著(P<0.05)水平,而与土壤速效N含量、有机质含量、总有机C含量和活性C含量的偏相关系数未达到显著水平(P>0.05)。根据标准误差检验,西藏高原高寒草原植被植物地上部分C/N值与土壤化学因子的回归方程通过了0.01水平的显著性检验。表明影响西藏高原高寒草原植被植物地上部分C/N值的主要土壤化学因子是土壤速效N、全N、全K、有机质、总有机C和活性C含量,其中土壤全N和全K含量的影响尤为显著,随土壤全N和全K含量的降低植物地上部分C/N值显著增大;此外,土壤化学因子对植物地上部分C/N值的影响达到极显著水平。
根系C/N值与土壤化学因子的回归方程为Y=46.299 9-0.019 8X2-0.025 2X4-4 559 013.597 6X8+7 859 749.973 6X9- 458.408 7X10+416.806 2X11-3.976 5X13-49 864.949 7X14(N=54,R=0.600 2,R2=-0.179 1,R4=-0.189 8,R8=-0.189 3,R9=0.189 3,R10=0.238 6,R11=0.207 6,R13= -0.257 1,R14=-0.165 8)。从方程中可看出:影响西藏高原高寒草原植被植物根系C/N值的主要土壤化学因子是土壤速效N含量(X2)、速效 K含量(X4)、有机质含量(X8)、总有机C含量(X9)、总碳水化合物含量(X10)、水解性C含量(X11)、活性C含量(X13)和HCO3-含量(X14),而土壤pH值(X1)、速效P含量(X3)、全N含量(X5)、全P含量(X6)、全K含量(X7)和水溶性C含量(X12)等土壤化学因子则对植物根系C/N值无明显影响。根据标准误差检验,植物根系C/N值与土壤化学因子的回归方程通过了0.01水平的显著性检验。表明影响西藏高原高寒草原植被植物根系C/N值的主要土壤化学因子是土壤速效N、速效K、有机质、总有机 C、总碳水化合物、水解性 C、活性 C和含量,随总有机C、总碳水化合物和水解性C含量的增加以及土壤速效N、速效K、有机质、活性C和含量的降低,植物根系C/N值增大;此外,土壤化学因子对植物根系C/N值的影响达到极显著水平,但影响植物地上部分与根系C/N值的土壤化学因子各不相同。
3 讨论和结论
由于植物C/N值存在相当大的变异性,利用碳储量与固定的C/N值来计算氮储量会产生较大的不确定性,因此,应构建不同植被类型的C/N值数据库,加强不同类型生态系统中植物C/N值的空间变异性研究,深入探讨植物C/N值分布的空间格局和调控因素,才能更加准确地估算植物碳氮储量和模拟生态系统碳氮循环过程[16]。从本研究结果看:西藏高原高寒草原植被C/N值在水平方向上呈现出东西部高中间低的态势和斑块状交错分布的格局;植被C/N高值区主要集中在以扎日南木错和洞措为核心的藏北高原腹地,而在马攸木拉山以西则为植被C/N低值区。无论是植物地上部分还是植物根系的C/N值,在不同草地型间和不同自然地带间均存在显著差异。西藏高原高寒草原生态系统植物地上部分的C/N平均值均高于内蒙古大针茅 (Stipa grandis P.Smirn.)草原[17]、中国东部南北样带森林[18]、青海高寒草甸[19]、川西北亚高山高寒草甸[20]、内蒙古天然白杄(Picea meyeri Rehd.et Wils.)林[21]、黄土高原草木植物[22]、全球草木植物[4]、北京及其周边草木植物[4]、闽江河口湿地植物[23]、秦岭亚高山草甸[24]、亚热带常绿阔叶林以及温带针阔混交林和热带季雨林[25];其植物根系的C/N平均值高于川西北亚高山高寒草甸[20]、秦岭亚高山草甸[24]、内蒙古天然白杄林和内蒙古天然白桦(Betula platyphylla Suk.)林[21],但低于黄土高原典型草原植物[22]、闽江河口湿地植物[23]和内蒙古科尔沁沙地植物[26]。由此可以看出:与其他区域相比,西藏高原高寒草原生态系统植被地上部分C/N值处于较高水平,但其植物根系C/N值处于中等水平。造成这一现象可能有2个原因:1)西藏高原地势高亢、气候较为寒冷,较低的气温抑制了有机物分解和矿化,降低了土壤N素的有效性,抑制了土壤养分的移动和根的吸收;2)西藏高原是世界上最年轻的高原,土壤发育较晚且质地较为粗糙,土壤N的淋溶作用较为强烈。
研究结果显示:西藏高原高寒草原植被地上部分C/N值与地上部生物量以及土壤全N和全K含量呈显著正相关、与植被高度呈显著负相关;其植物根系C/N值与海拔和20~30 cm土壤容重呈显著正相关、与年均降水量和年均蒸发量呈显著负相关。植物地上部分C/N值与纬度和海拔呈负相关、与经度呈正相关;而植物根系C/N值却与纬度和海拔呈正相关、与经度呈负相关。丁小慧等[26]认为:呼伦贝尔草地植物群落叶片C/N值随经度梯度升高而显著增加;王绍强等[16]认为:随着向赤道接近,森林叶C/N值会增加;刘超等[27]认为:黄土高原植物地上部分C/N值与年均降水量和年均温度不相关;作者针对西藏高原高寒草原植被植物地上部分C/N值与环境因子相关性的研究结果也得出了一致的结论。但韩文轩等[4]的结果表明:北京及周边地区植物地上部分C/N值与海拔呈正相关、与气候因子(年均温度和降水量)呈负相关,这一研究结果与作者的结论有异。其原因尚不清楚,有待进一步研究。
[1]MARSCHNER H.Mineral Nutrition of Higher Plants[M].New York:Academic Press,1995.
[2]NIKLAS K J,OWENS T,REICH P B,et al.Nitrogen/phosphorus leaf stoichiometry and the scaling of plant growth[J].Ecology Letters,2005,8(6):636-642.
[3]REICH P B,TJOELKER M G,MACHADO J L,et al.Universal scaling of respiratory metabolism,size and nitrogen in plants[J].Nature,2006,439:457-461.
[4]韩文轩,吴 漪,汤璐瑛,等.北京及周边地区植物叶的碳氮磷元素计量特征[J].北京大学学报:自然科学版,2009,45(5):855-860.
[5]STERNER R W,ELSER J J.Ecological Stoichiometry:the Biology of Elements from Molecules to the Biosphere[M].Princeton:Princeton University Press,2002.
[6]AGREN G I.The C ∶N ∶P stoichiometry of autotrophs-theory and observations[J].Ecology Letters,2004,7(3):185-191.
[7]VITOUSEK P.Nutrient cycling and nutrient use efficiency[J].American Naturalist,1982,119(4):553-572.
[8]ZHANG L X,BAI Y F,HAN X G.Application of N ∶P stoichiometry to ecology studies[J].植物学报,2003,45(9):1009-1018.
[9]王建林,欧阳华,王忠红,等.青藏高原高寒草原土壤活性有机碳的分布特征[J].地理学报,2009,64(7):771-781.
[10]张宪洲,石培礼,刘允芬,等.青藏高原高寒草原生态系统土壤CO2排放及其碳平衡[J].中国科学D辑:地球科学,2004,34(增刊Ⅱ):193-199.
[11]WANG L,OUYANG H,ZHOU C P,et al.Soil organic matter dynamics along a vertical vegetation gradient in the Gongga Mountain on the Tibetan Plateau[J].Journal of Integrative Plant Biology,2005,47(4):411-420.
[12]张金霞,曹广民.高寒草甸生态系统氮素循环[J].生态学报,1999,19(4):509-512.
[13]西藏自治区土地管理局编.西藏草地资源[M].北京:科学出版社,2001.
[14]中国科学院南京土壤研究所.土壤理化分析[M].上海:上海科学技术出版社,1978.
[15]唐明义,冯明光.实用统计分析及其DPS数据处理系统[M].北京:科学出版社,2000:286-297.
[16]王绍强,于贵瑞.生态系统碳氮磷元素的生态化学计量学特征[J].生态学报,2008,28(8):3937-3947.
[17]银晓瑞,梁存柱,王立新,等.内蒙古典型草原不同恢复演替阶段植物养分化学计量学[J].植物生态学报,2010,34(1):39-47.
[18]任书杰,于贵瑞,姜春明,等.中国东部南北样带森林生态系统102个优势种叶片碳氮磷化学计量学统计特征[J].应用生态学报,2012,23(3):581-586.
[19]王文颖,王启基,王 刚,等.高寒草甸土地退化及其恢复重建对植被碳、氮含量的影响[J].植物生态学报,2007,31(6):1073-1078.
[20]石福孙,陈华峰,吴 宁.增温对川西北亚高山高寒草甸植物群落碳、氮含量的影响[J].植物研究,2008,28(6):730-736.
[21]印婧婧,郭大立,何思源,等.内蒙古半干旱区树木非结构性碳、氮、磷的分配格局[J].北京大学学报:自然科学版,2009,45(3):519-527.
[22]董晓玉,傅 华,李旭东,等.放牧与围封对黄土高原典型草原植物生物量及其碳氮磷贮量的影响[J].草业学报,2008,19(2):175-182.
[23]章文龙,曾从盛,张林海,等.闽江河口湿地植物氮磷吸收效率的季节变化[J].应用生态学报,2009,20(6):1317-1322.
[24]何亚婷,刘文治,党高弟,等.秦岭亚高山草甸30种草本植物的碳、氮分布研究[J].草业科学,2008,25(10):1-5.
[25]王晶苑,王绍强,李纫兰,等.中国四种森林类型主要优势植物的C∶N∶P化学计量学特征[J].植物生态报,2011,35(6):587-595.
[26]丁小慧,罗淑政,刘金巍,等.呼伦贝尔草地植物群落与土壤化学计量学特征沿经度梯度变化[J].生态学报,2012,32(11):3467-3476.
[27]刘 超,王 洋,王 楠,等.陆地生态系统植被氮磷化学计量研究进展[J].植物生态学报,2012,36(11):1205-1216.
