氮-硫沉降对邓恩桉及杉木人工林凋落物C和N残留率的影响
2013-12-14包和林吴承祯林勇明
黄 婷,包和林,吴承祯,2,①,林勇明,洪 伟,李 键
(1.福建农林大学林学院,福建福州350002;2.武夷学院生态与资源工程系,福建武夷山354300)
杉木〔Cunninghamia lanceolata(Lamb.)Hook.〕为中国自然分布广、人工栽培多的速生商品树种[1],其材质结构均匀、抗虫耐腐性良好,在南方林区广泛种植,为中国亚热带地区人工林生态系统的主要组成树种。邓恩桉(Eucalyptus dunnii Maiden)是桉树中不可多得的纸浆原料树种,在土层厚、水肥好的环境条件下生长快且具较强的耐寒能力[2]。
以氮沉降和硫沉降为主导因子的酸沉降对森林枯落物的分解有较大影响[3]。在全球气候变化及工业化发展的背景下,大气氮、硫干湿沉降已成为森林生态系统中氮、硫的重要输入途径。其中,硫沉降可引起土壤理化性质改变,导致许多森林生态系统衰退[4];而氮沉降则对森林生态系统结构、功能及生物多样性有明显影响,过剩的氮与全球多地的森林衰退有密切关系[5-11]。目前,有关酸沉降对森林凋落物分解影响的研究主要集中于单一的氮沉降方面[12-14],对复合沉降的相关研究报道较少。
为此,本课题组借鉴段雷等[15]对中国土壤氮硫沉降的临界负荷区划结果,在酸沉降中等敏感的福建省开展了氮硫沉降条件下人工林凋落物与植株养分关系、胸径生长状况等方面的研究,获得了相关的研究结果[16-17]。在前期研究基础上,作者以邓恩桉和杉木人工林为研究对象,开展模拟氮-硫沉降条件下森林生态系统尤其是人工林生态系统凋落物元素动态变化的研究,评价酸沉降对人工林养分循环的影响效应,以期为人工林凋落物分解对全球酸沉降响应特征的研究、以及人工林生态系统的可持续经营和酸沉降减缓措施的提出提供相关研究数据。
1 研究区概况和研究方法
1.1 研究区概况
研究区位于福建省建阳市,属闽西北中低山丘陵中部,地理坐标为东经 118°09'19″、北纬 27°19'47″;平均海拔187 m。土壤类型为红壤和黄壤,土层深厚且肥沃。区域内光热资源丰富,适宜动植物生殖繁衍,为中国南方重点林区之一。该区域为中亚热带季风气候,冬短夏长、温暖湿润、静风多;年平均日照时数1 802 h,温差大;年均气温18℃,无霜期230~280 d;雨季集中,年平均降水量约为1 700 mm[1]。
供试样地2005年皆伐后萌发杉木幼龄林,每年进行2次除草追肥;杉木现存11 000株·hm-2,平均胸径3.53 cm、平均树高2.28 m。邓恩桉于2006年2月种植,造林地为杉木和马尾松(Pinus massoniana Lamb.)采伐迹地,土壤类型为红壤,造林密度1 750株·hm-2,每年进行3次除草追肥(500 g·株-1);邓恩桉现存1 650株·hm-2,平均胸径8.57 cm、平均树高9.1 m。邓恩桉和杉木林下植被稀疏,主要种类为芒萁〔Dicranopteris dichotoma(Thunb.)Bernh.〕和两色鳞毛蕨〔Dryopteris setosa(Thunb.)Akasawa〕。
1.2 研究方法
1.2.1 模拟氮-硫沉降实验设计 采用二次正交旋转组合设计[16-17]并参考段雷等[15]和 Gundersen 等[18]对酸沉降临界负荷的研究结果,确定模拟氮-硫沉降梯度和沉降量,其中mc=4、mr=4、m0=8。以Na2SO4为硫源、46%CO(NH2)2为氮源,二者的0编码水平分别为96和150 kg·hm-2·a-1。各处理水平为:A处理Na2SO4164 kg·hm-2·a-1和46%CO(NH2)2256 kg·hm-2·a-1;B 处理164 kg·hm-2·a-1Na2SO4和46%CO(NH2)244 kg·hm-2·a-1;C处理Na2SO428 kg·hm-2·a-1和 46%CO(NH2)2256 kg·hm-2·a-1;D处理 Na2SO428 kg·hm-2·a-1和46%CO(NH2)244 kg·hm-2·a-1;E 处理 Na2SO40 kg·hm-2·a-1和46%CO(NH2)2150 kg·hm-2·a-1;F处理Na2SO4192 kg·hm-2·a-1和 46%CO(NH2)2150 kg·hm-2·a-1;G 处理 Na2SO496 kg·hm-2·a-1和 46%CO(NH2)20 kg·hm-2·a-1;H 处理 Na2SO496 kg·hm-2·a-1和 46%CO(NH2)2300 kg·hm-2·a-1;I处理 Na2SO496 kg·hm-2·a-1和 46%CO(NH2)2150 kg·hm-2·a-1。其中I处理为零水平处理,设置8个重复;此外还设置1个空白处理,合计为17个处理组。
1.2.2 凋落物放置及模拟氮-硫沉降处理方法 于2007年10月在邓恩桉和杉木幼林内分别设置68块面积20 m×20 m的样地,不同处理样地间设3 m缓冲带。
2007年10月采用直接收集法采集样地内长势良好的邓恩桉和杉木新鲜叶片,采样叶位为植株顶部从上至下的第3个分枝。叶片自然风干后于65℃恒温烘干,按每袋10 g装入上下孔径均为0.5 mm的12 cm×12 cm尼龙网袋。次月底,在邓恩桉和杉木各样地中分别放入36个邓恩桉叶片或杉木叶片分解袋,分解袋悬挂在离地面1.3 m处。
根据上述模拟氮-硫沉降水平将46%CO(NH2)2和Na2SO4混合溶解于20 L水中,每月月底在样地用背式喷雾器均匀喷洒在分解袋上,对照则喷洒等量水;每月1次,持续1 a。
1.2.3 凋落物样品采集及C和N含量测定方法 于2007年10月第1次取样测定凋落物中C和N的本底值,其后于每月28日(与喷洒时间一致)从各样地中随机取回1个分解袋(4次重复),在同等条件下将凋落物烘干并称量,粉碎后供测定。参照文献[17],采用低温外加热重铬酸钾氧化-比色法测定凋落物中C含量,采用半微量凯氏定氮法测定N含量。
1.3 数据处理
采用EXCEL 2003统计分析软件对实验数据进行统计分析和作图,其中对零水平(处理I)的8个重复实验点的测定数据取平均值。用DPS 7.05统计分析软件进行方差分析、LSD多重比较及Olson指数模型拟合。
参照文献[19]采用Olson指数模型“y=ekt”预估凋落物中C和N分解速率,式中:y为凋落物中C或N的残留率;t为时间;k为分解系数(a-1)。
2 结果和分析
2.1 模拟氮-硫沉降对邓恩桉凋落物中C和N释放的影响
2.1.1 C和N残留率的动态变化 在模拟氮-硫沉降条件下经过1 a的处理周期邓恩桉凋落物中C和N残留率的变化见表1。结果表明:在不同的模拟氮-硫沉降水平下,随处理时间延长邓恩桉凋落物的C残留率在分解过程中均呈明显的下降趋势,其中前7个月C呈释放状态;至2008年10月各处理组C残留率均达到最低值,从A处理至I处理邓恩桉凋落物中C残留率依次为 39.47%、47.83%、50.28%、44.71%、49.10%、38.99%、51.61%、49.62% 和 45.84%。方差分析结果显示不同处理组及不同取样时间邓恩桉凋落物的C残留率均有极显著差异(P<0.01);LSD多重比较结果显示处理F〔Na2SO4192 kg·hm-2·a-1和46%CO(NH2)2150 kg·hm-2·a-1〕的C残留率与其余处理组有显著差异(P<0.05)。
在不同的模拟氮-硫沉降水平下,随处理时间延长各处理组邓恩桉凋落物的N残留率总体呈下降趋势,但N较难释放,其下降过程总体表现为“释放-富集-释放”的动态模式。各处理组凋落物的N均有明显的富集时间段,具体时间为2008年的1月、3月、4月、6月和7月;至2008年10月各处理组N残留率均达到最低值,从A处理至I处理邓恩桉凋落物中N残留率依次为 60.67%、74.34%、80.52%、69.65%、79.89%、63.45%、76.86%、80.52% 和 71.50%。方差分析结果显示不同处理组及不同取样时间邓恩桉凋落物的N残留率均有极显著差异(P<0.01);LSD多重比较结果显示处理C〔Na2SO428 kg·hm-2·a-1和46%CO(NH2)2256 kg·hm-2·a-1〕中邓恩桉凋落物的N残留率与其余处理组(除处理E外)有显著差异(P<0.05)。
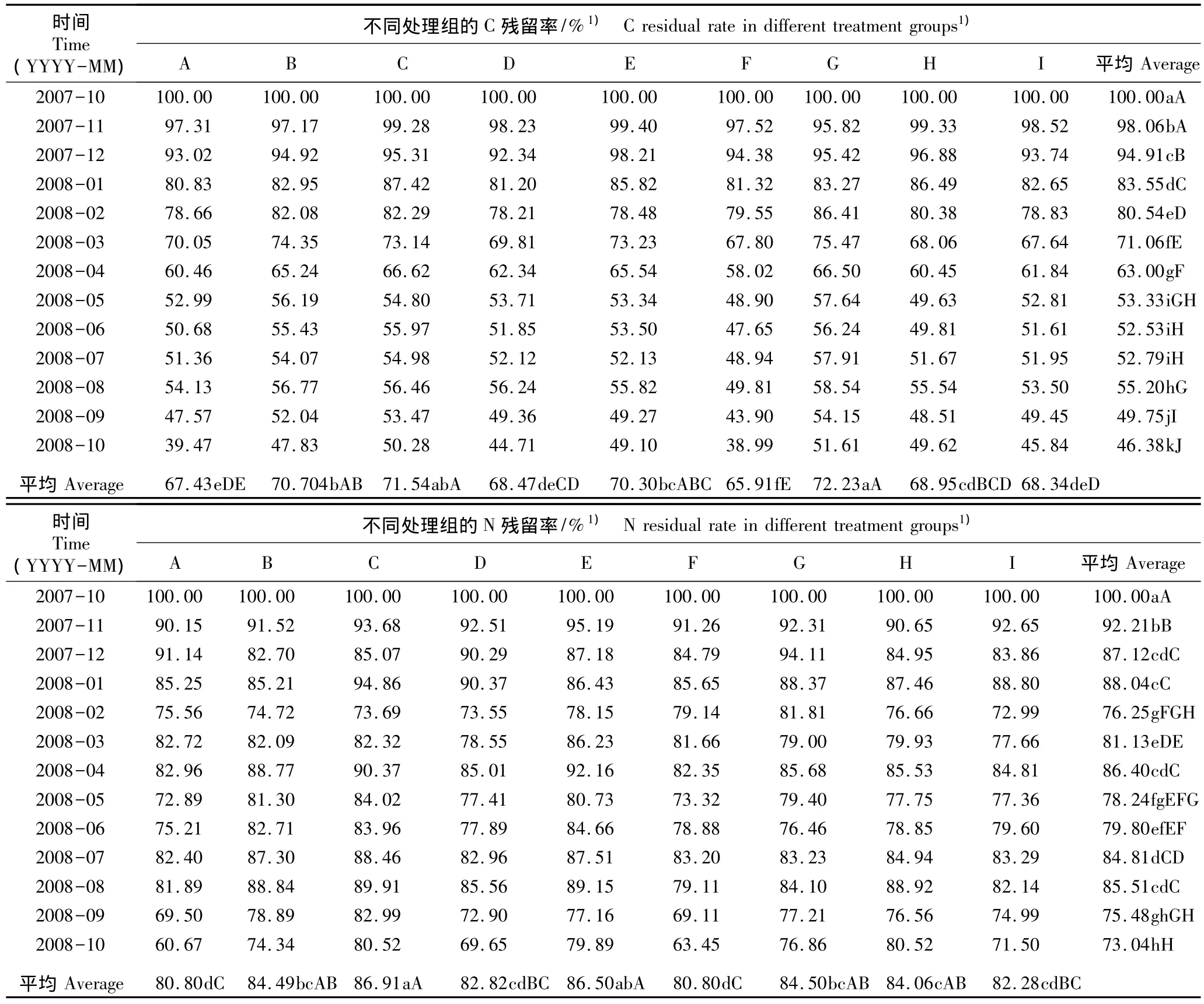
表1 在模拟氮-硫沉降条件下2007年10月至2008年10月邓恩桉凋落物中C和N的残留率Table 1 Residual rates of C and N in litter of Eucalyptus dunnii Maiden from Oct.2007 to Oct.2008 under condition of simulated nitrogensulfur deposition
2.1.2 C和N的分解模型 采用Olson指数模型对不同氮-硫沉降水平下邓恩桉凋落物中C和N的分解模型进行拟合,结果见表2。由表2可见:各处理组邓恩桉凋落物中C分解模型的相关系数均达到极显著水平(P<0.01),相关性优于其N分解模型。各处理组邓恩桉凋落物的C和N的平均分解系数分别为0.877和 0.208,C 和 N 的平均周转期分别为 3.148和15.877 a,表明邓恩桉凋落物中C释放速率大于N释放速率。按照邓恩桉凋落物中C释放速率从大至小各处理组依次排序为 F、A、H、I、D、E、B、C、G;按照N释放速率从大至小各处理组依次排序为A、F、D、I、G、E、H、B、C。
结合不同处理组的模拟氮-硫沉降水平可以看出,在 Na2SO4192 kg·hm-2·a-1和 46%CO(NH2)2150 kg·hm-2·a-1的水平下C释放最快;在Na2SO4164 kg·hm-2·a-1和 46%CO(NH2)2256 kg·hm-2·a-1的水平下N释放最快;在Na2SO428 kg·hm-2·a-1或96 kg·hm-2·a-1的水平下C和N释放速率随氮沉降水平提高而减小;在Na2SO4164 kg·hm-2·a-1的水平下,氮沉降促进 C和 N的释放;在 46%CO(NH2)244 kg·hm-2·a-1的水平下,C和N的释放速率随硫沉降水平的提高而减小;在46%CO(NH2)2150或256 kg·hm-2·a-1的水平下,硫沉降对C和N的释放有促进作用。
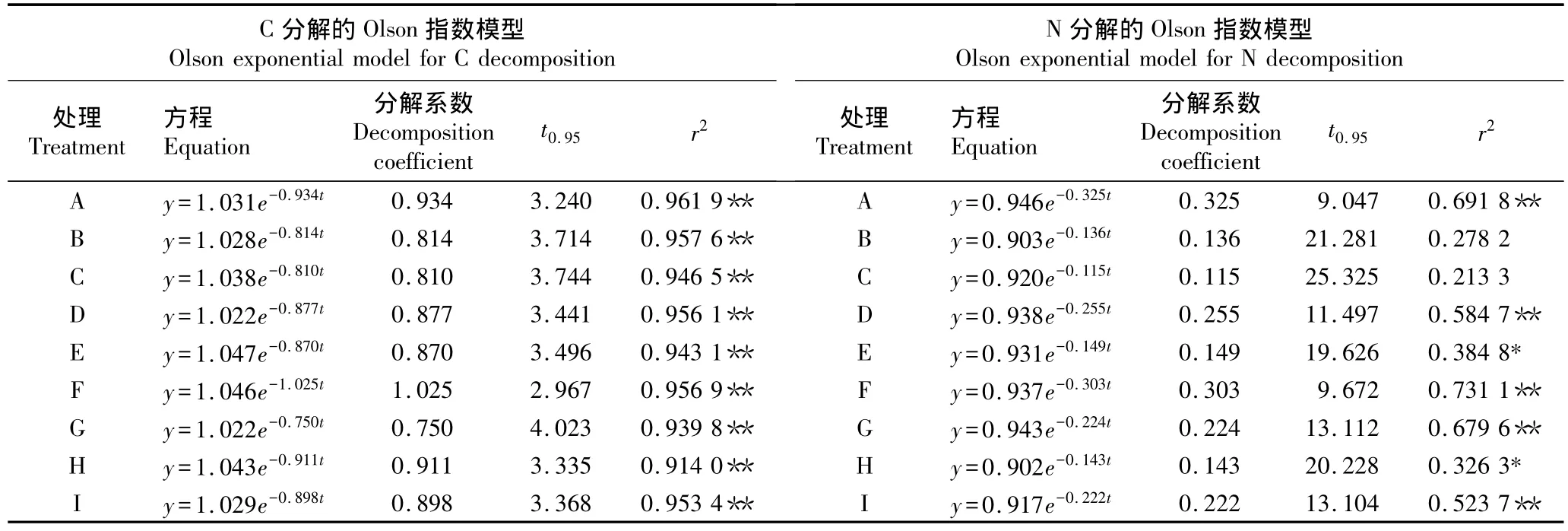
表2 在模拟氮-硫沉降条件下2007年10月至2008年10月邓恩桉凋落物中C和N分解的Olson指数模型1)Table 2 Olson exponential models for C and N decomposition in litter of Eucalyptus dunnii Maiden from Oct.2007 to Oct.2008 under condition of simulated nitrogen-sulfur deposition1)
2.2 模拟氮-硫沉降对杉木凋落物中C和N释放的影响
2.2.1 C和N残留率的动态变化 在模拟氮-硫沉降条件下经过1 a的处理周期杉木凋落物中C和N残留率的变化见表3。结果表明:在1 a的处理过程中,前2个月杉木凋落物中C残留率呈明显的下降趋势,各处理组(除处理G和H外)的C残留率在2008年1月均小幅上升;但在此后的5个月中各处理组的C残留率均逐月下降;至2008年7月,除处理B外各处理组的C残留率均显著增加;至2008年10月,从A处理至I处理凋落物的C残留率依次为48.61%、45.00%、43.15%、46.20%、42.03%、44.43%、51.03%、44.69%和46.68%。方差分析结果显示不同处理组及不同取样时间杉木凋落物的C残留率均有极显著差异(P<0.01);LSD多重比较结果显示处理 G〔Na2SO496 kg·hm-2·a-1和 46%CO(NH2)20 kg·hm-2·a-1〕的C残留率与其余处理组(除处理A 外)有显著差异(P<0.05)。
在不同的模拟氮-硫沉降水平下,随处理时间延长各处理组杉木凋落物的N残留率在分解过程中总体呈下降趋势。具体表现为:前2个月N残留率呈明显的下降趋势,其后2个月基本保持平衡,至2008年3月均小幅上升,2008年4月至10月逐渐下降;至2008年10月从A处理至I处理凋落物的N残留率依次为 46.13%、49.08%、51.77%、45.03%、52.04%、48.78%、44.93%、49.58% 和 48.15%。整体上看:处理 E〔Na2SO40 kg·hm-2·a-1和 46%CO(NH2)2150 kg·hm-2·a-1〕的杉木凋落物N残留率最大,处理G的N残留率最小,其余处理条件下杉木凋落物交替进行N释放与N富集,且释放量大于富集量。方差分析结果显示不同处理组及不同取样时间杉木凋落物的N残留率有极显著差异(P<0.01);LSD多重比较结果显示处理G的N残留率与其余处理组有显著差异(P<0.05)。
2.2.2 C和N的分解模型 采用Olson指数模型对不同氮-硫沉降水平下杉木凋落物中C和N的分解模型进行拟合,结果见表4。由表4可见:各处理组杉木凋落物的C和N分解模型的相关系数均达到显著水平(P<0.05),拟合效果较好。各处理组C和N的平均分解系数分别为0.704和0.600,平均周转期分别为4.090和4.947 a,说明杉木凋落物的C释放速率大于N释放速率。按照杉木凋落物的C释放速率从大至小各处理组依次排序为 H、I、B、F、C、A、E、D、G,按照N释放速率从大至小各处理组依次排序为D、F、I、G、A、H、C、B、E。
结合不同处理组的模拟氮-硫沉降水平可以看出:在 Na2SO496 kg·hm-2·a-1和 46%CO(NH2)2300 kg·hm-2·a-1的水平下杉木凋落物中C释放速率最快;在Na2SO428或96 kg·hm-2·a-1的水平下C释放速率随氮沉降水平的升高而增大;在Na2SO4164 kg·hm-2·a-1的水平下,N沉降对杉木凋落物的C释放有抑制作用;在46%CO(NH2)244 kg·hm-2·a-1的水平下,C释放速率随硫沉降水平的升高而增大;在46%CO(NH2)2256 kg·hm-2·a-1的水平下,硫沉降对杉木凋落物的 C释放有抑制作用。在Na2SO428 kg·hm-2·a-1和 46%CO(NH2)244 kg·hm-2·a-1的水平下,杉木凋落物中N释放速率最快;在Na2SO428 kg·hm-2·a-1的水平下,N释放速率随氮沉降水平的升高而降低;在Na2SO4164 kg·hm-2·a-1的水平下,氮沉降可促进凋落物的 N释放;在 46%CO(NH2)244 kg·hm-2·a-1的水平下,凋落物中N释放速率随硫沉降水平的升高而降低;在46%CO(NH2)2150或256 kg·hm-2·a-1的水平下,硫沉降可促进杉木凋落物中的N释放。
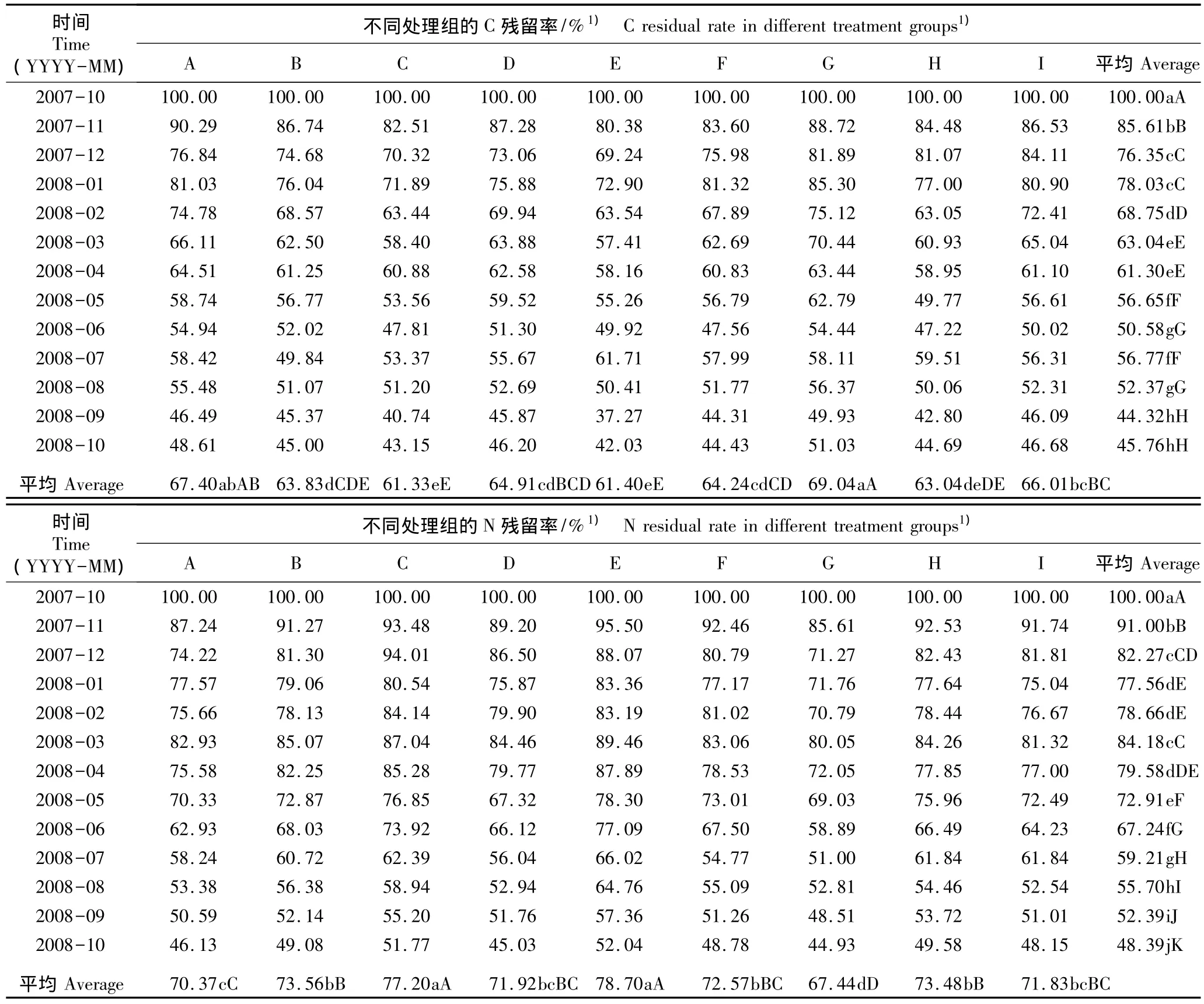
表3 在模拟氮-硫沉降条件下2007年10月至2008年10月杉木凋落物中C和N的残留率Table 3 Residual rates of C and N in litter of Cunninghamia lanceolata(Lamb.)Hook.from Oct.2007 to Oct.2008 under condition of simulated nitrogen-sulfur deposition
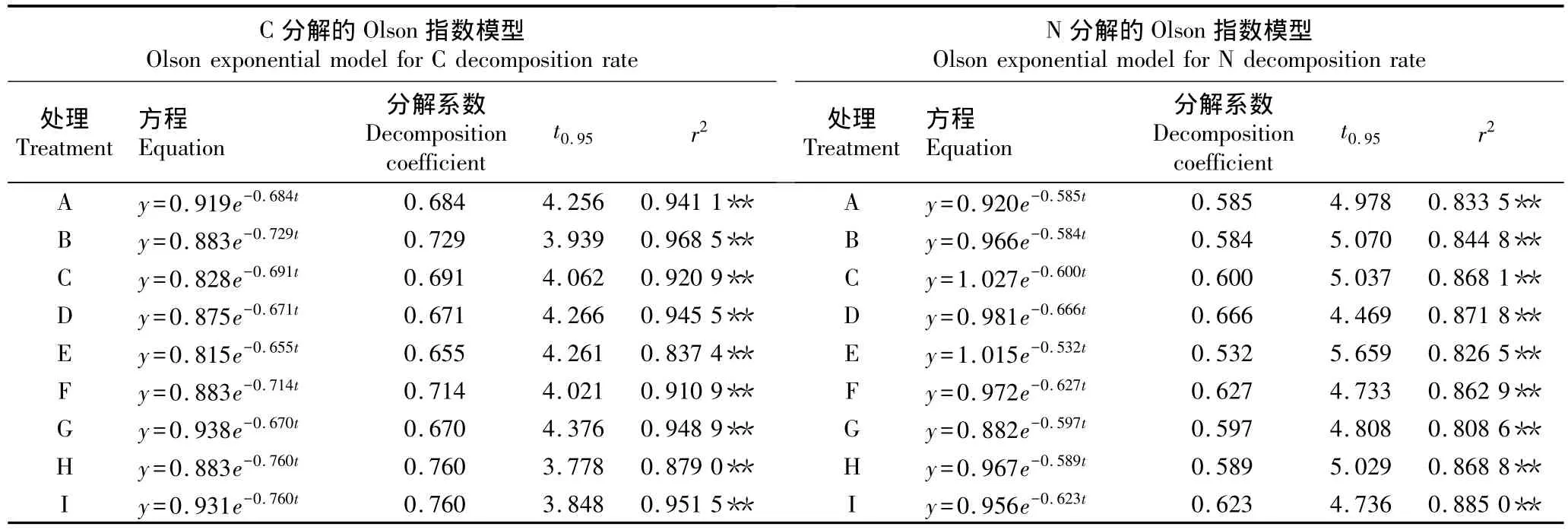
表4 在模拟氮-硫沉降条件下2007年10月至2008年10月杉木凋落物中C和N分解的Olson指数模型1)Table 4 Olson exponential models for C and N decomposition in litter of Cunninghamia lanceolata(Lamb.)Hook.from Oct.2007 to Oct.2008 under condition of simulated nitrogen-sulfur deposition1)
2.3 模拟氮-硫沉降条件下邓恩桉和杉木凋落物的C/N值动态变化
在模拟氮-硫沉降条件下经过1 a的处理周期邓恩桉和杉木凋落物C/N值的变化见表5。在不同氮-硫沉降水平下各处理组凋落物的C/N值呈波动但总体减小的趋势。除处理A、D和G外,各处理条件下其C/N值在前2个月呈明显增大的趋势,2008年1月大幅减小,2008年2月则小幅增大,2008年3月至6月又大幅减小,其后4个月则小幅波动;至2008年10月,从A处理至I处理凋落物中的C/N值分别比初始值(45.21)下降了34.95%、35.67%、37.56%、35.80%、38.54%、38.55%、32.86%、38.37% 和35.91%。方差分析结果显示其C/N值在不同处理间有极显著差异(P<0.01);LSD多重比较结果显示处理G的C/N值与其他处理组(除处理B外)有显著差异(P<0.05)。

表5 在模拟氮-硫沉降条件下2007年10月至2008年10月邓恩桉和杉木凋落物的C/N值Table 5 Value of C/N in litters of Eucalyptus dunnii Maiden and Cunninghamia lanceolata(Lamb.)Hook.from Oct.2007 to Oct.2008 under condition of simulated nitrogen-sulfur deposition
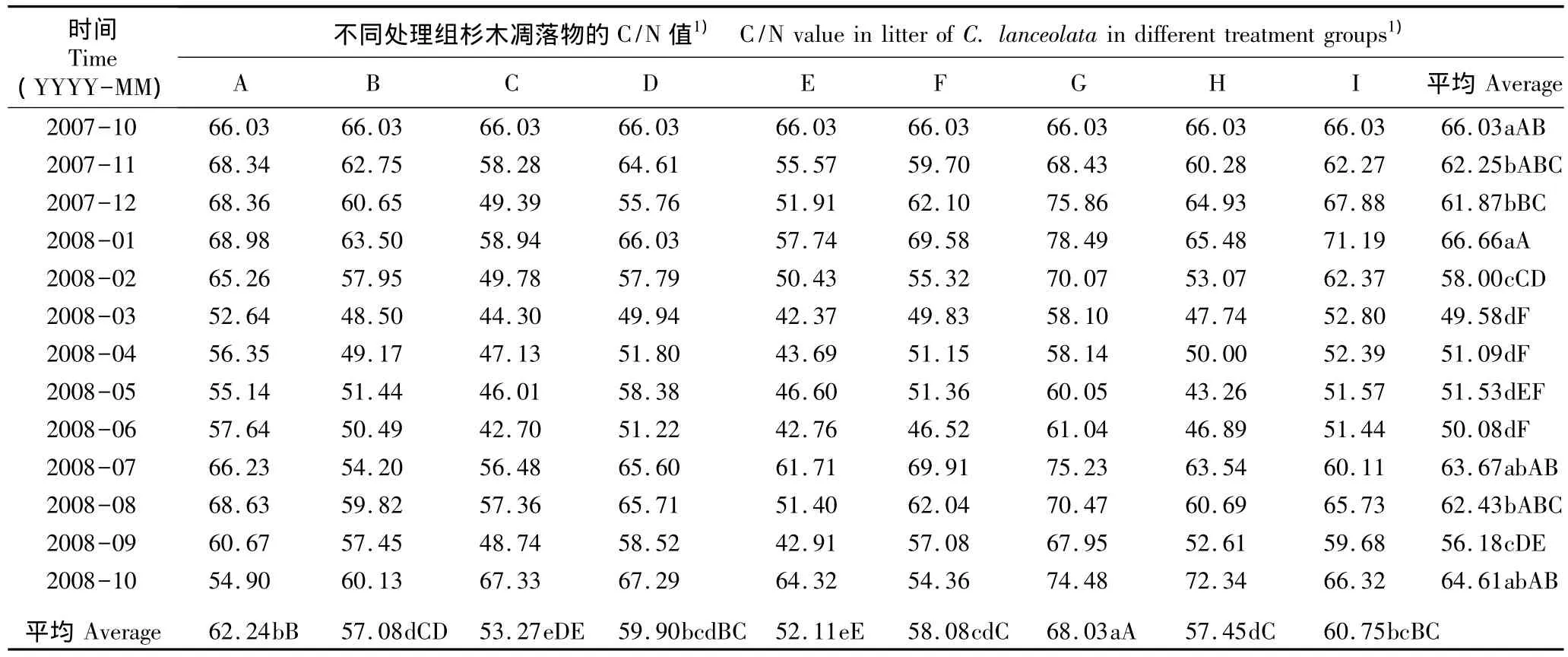
续表5 Table 5(Continued)
在不同氮-硫沉降水平下各处理组杉木凋落物的C/N值呈波动的趋势。其C/N值在前3个月变化规律复杂,2008年1月至3月均呈下降趋势,2008年3月至6月基本不变;至2008年10月,从A处理到I处理杉木凋落物的C/N值分别比其初始值(66.03)下降了16.90%、8.94%、-1.97%、-1.91%、2.59%、17.67%、-12.80%、-9.56% 和-0.44%。方差分析结果显示其C/N值在不同处理间有极显著差异(P<0.01),LSD多重比较结果显示处理E的C/N值与其他处理组(除处理C外)有显著差异(P<0.05)。
3 讨论和结论
上述研究结果表明:在不同的模拟氮-硫沉降条件下,邓恩桉和杉木凋落物分解过程中C和N的残留率均呈下降趋势,主要是因为氮-硫沉降时间的延长会逐渐加快凋落物中有机物的分解。从C和N的释放状态看,N较难释放,总体表现为“释放-富集-释放”的动态过程,前期凋落物无法适应氮-硫沉降过程而释放N,适应外界变化之后凋落物则吸收N,达到饱和之后表现出N释放,形成了动态的变化过程。邓恩桉凋落物C释放的Olson拟合模型相关性较大,而杉木凋落物N释放的Olson拟合模型相关性较大;杉木凋落物的C周转期大于邓恩桉,而其凋落物的N周转期则小于后者。整体上看,在不同的模拟氮-硫沉降条件下邓恩桉凋落物C/N值的变化幅度大于杉木凋落物,可能因为邓恩桉凋落物对氮-硫沉降的响应比杉木强烈。在同样的氮-硫沉降水平下,邓恩桉凋落物与杉木凋落物的C释放速率的变化相异。在N释放速率方面,在Na2SO428 kg·hm-2·a-1水平下2种植物凋落物的N释放速率随氮沉降量的增加而降低,而在Na2SO4164 kg·hm-2·a-1水平下氮沉降可促进2种植物凋落物的N释放;在46%CO(NH2)244 kg·hm-2·a-1水平下邓恩桉和杉木凋落物的N释放速率随硫沉降量增加而降低,而在46%CO(NH2)2150或256 kg·hm-2·a-1水平下,硫沉降可促进邓恩桉和杉木凋落物的N释放。
不同植物凋落物的分解对氮沉降的响应不同。有些研究者认为部分森林、草地和湿地生态系统的氮沉降可促进微生物活性[20];也有研究者[21]认为氮沉降可抑制微生物活性进而影响凋落物分解;但Hobbie等[22]却认为凋落物分解对氮沉降的响应不明显,可能与氮沉降量增加对微生物酶系统的影响及群落组成差异有关[23]。杉木凋落物的C释放速率大于N释放速率,这一结果与樊后保等[23]的研究结果一致。在模拟氮-硫沉降条件下杉木凋落物的C/N值在2008年1月至3月呈显著下降的趋势,可能由于N分解对氮-硫沉降的响应比C大,与樊后保等[23]和项文化等[24]的研究结果相似。杉木凋落物中C/N平均值比邓恩桉凋落物高,也大于樊后保等[25]测得的杉木凋落物C/N平均值(44.01);本研究中,邓恩桉凋落物的N周转期大于杉木凋落物,也大于樊后保等[25]测得的杉木凋落物的N周转期。主要原因可能是凋落物中C/N值有一个临界值,高于临界值则表现为N释放、较低则表现为N富集,且试验过程中氮沉降量的增加可能加剧了邓恩桉凋落物的N富集作用。
植物凋落物的分解与气候、土壤和微生物等环境因素密切相关[25]。随季节的变化本研究区域的气候及降雨量的变化幅度相对较大,可能会增强凋落物对氮-硫沉降的响应,但是否对凋落物的分解有明显影响则有待进一步的实验和研究。
[1]林开敏,章志琴,叶发茂,等.杉木人工林下杉木、楠木和木荷叶凋落物分解特征及营养元素含量变化的动态分析[J].植物资源与环境学报,2010,19(2):34-39.
[2]谭 艳,吴承祯,洪 伟,等.邓恩桉林地土壤pH空间变异分析[J].植物资源与环境学报,2012,21(1):14-19.
[3]HALL S J,MATSON P A.Nitrogen oxide emissions after nitrogen additions in tropical forests[J].Nature,1999,400:152-155.
[4]陈建忠.邓恩桉优树选择标准[J].福建林学院学报,2006,26(4):368-371.
[5]赵 亮,周国逸,张德强,等.CO2浓度升高和氮沉降对南亚热带主要乡土树种及群落生物量的影响[J].应用生态学报,2011,22(8):1949-1954.
[6]李德军,莫江明,方运霆,等.模拟氮沉降对南亚热带两种乔木幼苗生物量及其分配的影响[J].植物生态学报,2005,29(4):543-549.
[7]段洪浪,刘菊秀,邓 琦,等.CO2浓度升高与氮沉降对南亚热带森林生态系统植物生物量积累及分配格局的影响[J].植物生态学报,2009,33(3):570-579.
[8]方运霆,莫江明,GUNDERSEN P,等.森林土壤氮素转换及其对氮沉降的响应[J].生态学报,2004,24(7):1523-1531.
[9]鲁显楷,莫江明,董少峰.氮沉降对森林生物多样性的影响[J].生态学报,2008,28(11):5532-5548.
[10]邢军会,倪红伟,王建波.二氧化碳浓度升高与氮沉降对三江平原小叶章群落生物量累积及其分配格局的影响[J].中国农学通报,2011,27(13):49-54.
[11]涂利华,戴洪忠,胡庭兴,等.模拟氮沉降对华西雨屏区撑绿杂交竹林土壤呼吸的影响[J].应用生态学报,2011,22(4):829-836.
[12]WRIGHT R F,ROELOFS J G M,BREDEMEIER M,et al.NITREX:responses of coniferous forest ecosystems to experimentally changed deposition of nitrogen[J].Forest Ecology and Management,1995,71(1/2):163-169.
[13]樊后保,袁颖红,王 强,等.氮沉降对杉木人工林土壤有机碳和全氮的影响[J].福建林学院学报,2007,27(1):1-6.
[14]莫江明,薛璟花,方运霆.鼎湖山主要森林植物凋落物分解及其对N沉降的响应[J].生态学报,2004,24(7):1413-1420.
[15]段 雷,郝吉明,谢绍东,等.用稳态法确定中国土壤的硫沉降和氮沉降临界负荷[J].环境科学,2002,23(2):7-12.
[16]侯 丹,吕广林,张艳荷,等.邓恩桉胸径生长对模拟硫、氮复合沉降的响应[J].福建林学院学报,2009,29(2):125-130.
[17]吴承祯,洪 伟,陈 灿,等.氮、硫沉降下邓恩桉人工林土壤与植株养分特征关系研究[J].武夷学院学报,2013,32(2):1-7.
[18]GUNDERSEN P,EMMETT B A,KJØNAAS O J,et al.Impact of nitrogen deposition on nitrogen cycling in forest:a synthesis of NITREX data[J]. Forest Ecology and Management, 1998,101(1/3):37-55.
[19]刘增文,潘开文.Olson枯落物分解模型存在的问题与修正[J].西北农林科技大学学报:自然科学版,2005,33(1):69-70.
[20]BERG B,STAAF H.Decomposition rate and chemical changes of Scots pine needle litter.Ⅱ.Influence of chemical composition[J].Ecological Bulletins,1980,32:373-390.
[21]CARREIRO M M,SINSABAUGH R L,REPERT D A,et al.Microbial enzyme shifts explain litter decay responses to simulated nitrogen deposition[J].Ecology,2000,81(9):2359-2365.
[22]HOBBIE S E,VITOUSEK P M.Nutrient limitation of decomposition in Hawaiian forests[J].Ecology,2000,81(7):1867-1877.
[23]樊后保,刘文飞,徐 雷,等.氮沉降下杉木(Cunninghamia lanceolata)人工林凋落叶分解过程中C、N元素动态变化[J].生态学报,2008,28(6):2546-2553.
[24]项文化,闫文德,田大伦,等.外加氮源及与林下植物叶混合对杉木林针叶分解和养分释放的影响[J].林业科学,2005,41(6):1-6.
[25]樊后保,刘文飞,杨跃霖,等.杉木人工林凋落物分解对氮沉降增加的响应[J].北京林业大学学报,2008,30(2):8-13.
