5个猪种CAST基因第二内含子微卫星标记的遗传多态性分析
2013-11-30赵先萍张燕青何永梅高鹏飞郭晓红李步高曹果清
赵先萍,张燕青,梁 琳,何永梅,高鹏飞,郭晓红,李步高,曹果清*
(1.山西省畜牧兽医学校,山西 太原 030024;2.山西农业大学 动物科技学院,山西 太谷 030801)
钙蛋白酶抑制蛋白(Calpastatin,CAST)为内源性专一抑制钙蛋白酶活性的蛋白质,可识别钙蛋白酶与钙结合引起的构象变化与特异性结合,抑制其活性降低蛋白质水解[1],影响肌肉嫩度。肌肉嫩度主要取决于肌纤维直径大小、肌节长度、肌内脂肪含量和结缔组织等因素。研究表明,CAST等位基因A(Lys)和G(Arg)与肌肉的嫩度、屠宰45 min后pH值及滴水损失存在强相关[2];CAST/Rsa I和CAST/Msp I多态位点对背膘厚、大理石纹有明显影响[3];CAST/Hinf I位点对肌内脂肪含量、肉色评分、失水率和贮存损失有显著影响[4]。CAST基因在多个性状上影响着胴体品质和肉质。
对猪CAST基因分析发现,其第2内含子中存在以GT为核心序列的微卫星座位,有关该座位的遗传变异和效应还未见报道。本研究分析了长白猪、大白猪、杜洛克猪、马身猪和山西黑猪的CAST基因第2内含子微卫星座位的遗传多态性,为该基因应用于改善家畜肉质品质等方面奠定了基础。
1 材料与方法
1.1 试验猪群及材料
长白猪60头、大白猪54头、杜洛克猪35头、马身猪42头、山西黑猪36头,均来自山西省大同市种猪场。用耳号钳剪耳组织约0.1 g,放入装有1 mL 70%乙醇的1.5 mL离心管中,冰壶带回实验室,-70 ℃冰箱保存备用。研究中所用的Taq DNA Polymerase、dNTPs、50 bp DNA Ladder Marker等试剂均为宝生物工程(大连)有限公司产品。
1.2 试验方法
1.2.1 猪基因组DNA的提取与检测 采用常规的酚/氯仿抽提法提取基因组DNA,用0.8%琼脂糖凝胶电泳进行检测。
1.2.2 PCR扩增 根据CAST基因第二内含子序列,采用primer 3.0软件在线引物,上游引物为:5'-AGGAACCCAGAAGAGGTGCT-3';下游引物为:5'-TGTCCTTGCCCTCTAAGATGT-3',引物由华大基因(深圳)公司合成,扩增产物约189 bp。 PCR反应体系为20 μL,包括:2.0 μL 10 PCR Buffer (Mg2+)、0.4 μL dNTPs (10 mmol/L)、1.0 μL Mixed Primer、0.2 μL Taq DNA polymerase (5U/μL)、2.0 μL DNA 模板、14.4 μL ddH2O。反应条件为:94 ℃预变性5 min;94 ℃变性30 s,56.6 ℃退火30 s,72 ℃延伸30 s;最后72 ℃保温7 min,最终4 ℃保存。PCR扩增产物用1%琼脂糖凝胶电泳进行检测。
1.2.3 基因型分型 扩增良好的个体用12%的非变性聚丙烯酰胺凝胶电泳进行基因型分型,银染显色。用Bio-Rad公司凝胶成像系统的Quantity One软件计算等位基因大小,判定个体基因型。所用的分子量标记为宝生物工程(大连)有限公司的50 bp DNA Ladder Marker。
1.3 数据统计与分析
利用Popgen32和PIC软件计算等位基因频率、基因型频率、有效等位基因数、遗传纯合度、遗传杂合度和多态信息含量等指标。
2 结果与分析
2.1 微卫星多态性检测及基因频率分布
在所检测的群体中,共检测到4个等位基因,分别是A(171 bp)、B(179 bp)、C(185 bp)和D(191 bp),存在10种基因型。在5个供试猪群中,在该座位上的基因频率和基因型频率分布见表1。由表1可知,在山西黑猪和杜洛克猪群体中,等位基因D的频率较高,分别为0.6389和0.5143;在马身猪群体中,等位基因B的频率较高,为0.6310;在大白猪和长白猪群体中,4种等位基因的频率相近,分别介于0.1389~0.3889和0.1417~0.3500。就基因型频率分布来看,在山西黑猪群体中,检测到6种基因型,DD型频率最高,为0.3889;在马身猪群体中,也检测到6种基因型,BB型和AB型频率较高,分别为0.3810和0.3572;在长白猪和大白猪群体中,分别检测到9种和8种基因型;在杜洛克群体中,检测到9种基因型,DD型和AD型频率较高,分别为0.3143和0.2286。

表1 猪CAST基因第2内含子微卫星座位在不同猪种中的等位基因频率和基因型频率Table 1 Allele frequency and genotypic frequency of the microsatellite locus within intron 2 of CAST gene in different pig populations
2.2 5个猪种CAST基因第2内含子微卫星座位的遗传特性分析
5个猪种CAST基因第2内含子微卫星座位有效等位基因数、遗传纯合度、遗传杂合度、多态信息含量等指标见表2。由表2可知,在该座位上,长白猪和大白猪的有效等位基因数较高,分别为3.6567和3.4570,而山西黑猪和马身猪的有效等位基因数偏低,分别为2.1910和2.0888。长白猪和大白猪的遗传纯合度较高,分别为0.6667和0.7037,山西黑猪和马身猪的遗传杂合度较高,分别为0.6111和0.5476。在该位点上,除马身猪为中度多态外,其余品种均呈高度多态。
3 讨论与小结
钙蛋白酶抑制蛋白(Calpastatin, CAST)是细胞内钙蛋白酶系统的内源性抑制剂。研究表明CAST基因活性提高会增加钙蛋白酶抑制蛋白的表达,抑制肌肉蛋白质水解,促使蛋白质降解减少,影响肌肉嫩度。因此,CAST基因被认为是影响肉质品质的候选基因,基因内的突变位点对肉质品质有显著影响。研究表明,CAST基因cDNA序列上的A317G和G2042C突变对嫩度及背膘厚有显著影响,而对pH值和滴水损失无显著影响[5];内含子24上的A916G 和C1633G突变对肌肉嫩度、屠宰45 min后pH值及滴水损失有显著影响(P<0.05)[6];内含子6上的MspⅠ多态位点对肌肉嫩度、pH 值、系水力有显著影响(P<0.05),HinfⅠ酶切位点对肌内脂肪含量有显著影响(P<0.05 ),对肌肉嫩度有极显著影响(P<0.01)[7]。本研究小组在猪CAST基因外显子9检测到C193T和C256T错义突变,C193T引起苏氨酸向异亮氨酸的转变,C256T引起苏氨酸向蛋氨酸的转变,但这两个突变位点对山西白猪6月龄活体背膘厚无显著影响(未发表)。鸡CAST基因CDS区的G35193T突变为错义突变,可引起氨基酸Tyr(D)-Asp(Y) 的变异,该突变位点对鸡肌纤维密度和直径有着显著的影响(P<0.05),同日龄鸡群AA型个体的肌纤维密度高于AB型、肌纤维直径低于AB型,且均达到显著水平(P<0.05);同体重鸡群BB型个体的肌纤维密度高于AB型、肌纤维直径低于AB型,且均达到显著水平(P<0.05)[8]。牛CAST基因内含子6上检测到XmnⅠ多态位点,对剪切力值有显著影响,BB纯合型与AB杂合型的效应值明显小于AA型,差异达到显著水平,而对大理石纹的影响未达到显著水平[9];延边黄牛CAST基因内含子16上存在g101681A→G、g101745 A→G、g101758 T→C、g101828 T→C、g101888 T→C 及g101912 G→A 等6个突变位点,该位点基因型多态性与蒸煮损失、亚麻酸含量及肉色等多种肉质性状相关[10]。由此可见,CAST基因不同区域的遗传变异都可能影响到畜禽的肉质品质。
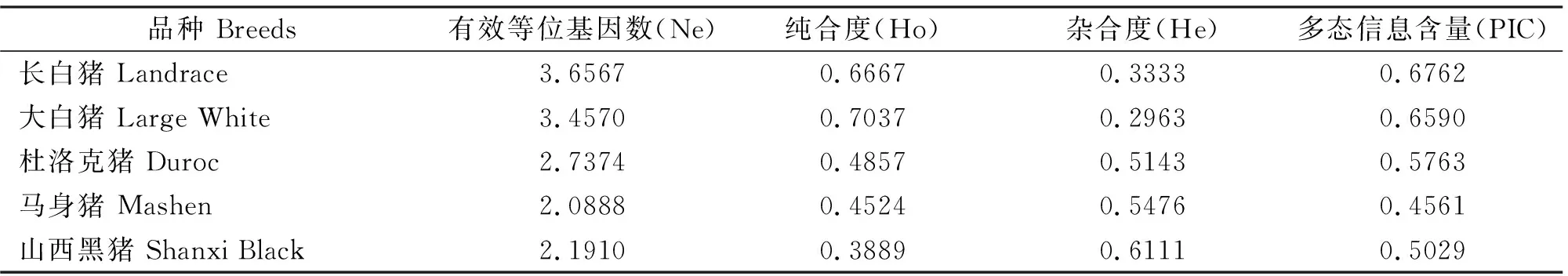
表2 不同猪种CAST基因第2内含子微卫星座位有效等位基因数、纯合度、杂合度和多态信息含量Table 2 Effective allele number (Ne), homozygosity (Ho), heterozygosity (He), and polymorphism informationcontent (PIC) for the microsatellite locus within intron 2 of CAST gene in different pig populations
在真核生物基因表达过程中,由于mRNA的剪接,内含子在成熟的mRNA和蛋白质中不作为编码序列出现,但在基因转录中有重要的调控作用[11]。Joan等[12]研究发现IGF-I基因第1内含子的以CA为重复序列的微卫星座位与160日龄血清IGF-I浓度、背膘厚和瘦肉量有关,该座位与猪的平均日增重也有关联[13]。本研究中,猪CAST基因第2内含子存在以GT为核心序列的微卫星座位,检测到4个等位基因和10种基因型,在不同猪种中,优势等位基因不完全相同,在山西黑猪和杜洛克猪群体中,D为优势等位基因,频率分别为0.6389和0.5143;在马身猪群体中,等位基因B的频率较高为0.6310。在不同类型猪种中,等位基因存在的方式不同,长白猪和大白猪的遗传纯合度较高,而山西黑猪和马身猪的遗传杂合度较高。鉴于CAST基因对猪肉质品质的影响及该微卫星座位基因型分布在不同品种中的差异,我们推测该微卫星座位可能对肉质品质有一定影响,但还有待于进一步研究证实。
参考文献:
[1] 邱莫寒,鲜凌瑾,张 平.钙蛋白酶系统基因的研究进展[J].安徽农业科学,2009,37(28),13 628-13 631.
[2] Xue H L,Xu L X.Calpastatin genotype is associated with meat quality traits in pigs[J]. Chinese Journal of Biochemistry and Molecular Biology,2009,25(7):615-619.
[3] Jolanta K, Wojciech K, Mariusz P, et al. Preliminary observations on the effect of calpastatin gene (CAST) polymorphism on carcass traits in pigs[J]. Animal Science Papers and Reports,2003,21(2):87-95.
[4] 邓正德,连林生.大河乌猪钙蛋白酶抑制蛋白基因、脂肪细胞定向和分化因子1基因多态性及其与肉质性状的关联分析[J].养猪,2012(3): 57-60.
[5] 薛慧良,徐来祥. 猪CAST基因的遗传多态性及遗传效应分析[J].分子细胞生物学报,2007,40(6): 403-409.
[6] Xue H L,Xu L X. Single nucleotide polymorphisms and genetic effects of CAST gene on meat quality traits in pigs[J]. Acta Ecologica Sinica,2008,28(6):2 937-2 944.
[7] 刘 香,虞德兵,邱新深,等.苏太猪CAST基因多态性与肉质性状的相关性研究[J].畜牧与兽医,2010,42(4): 15-18.
[8] 刘安芳,刘益平,蒋小松,等. 鸡CAST基因编码区的单核苷酸多态与肌纤维性状的相关研究[J].畜牧兽医学报, 2008, 39(4): 437-442.
[9] 邓桂馨,张银花,梁鸿斌.肉牛CAST基因分子多态性与肉品质性状相关性的研究[J]. 北京农学院学报, 2007, 22(3): 25-27.
[10] 张立春,李赵志,金 鑫, 等.延边黄牛CAST 基因16内含子基因多态性与肉质性状相关性分析[J].中国畜牧杂志,2011,47(15):12-15.
[11] Foti D M,Reichardt J K. YY1 binding within the human HSD3B2 gene intron 1 is required for maximal basal promoter activity: identification of YY1 as the 3beta1-A factor[J]. Journal of Molecular Endocrinology, 2004, 33(1): 99-119.
[12] Joan E, Marc T, Daniel V, et al. Association of CA repeat polymorphism at intron 1 of insulin-like growth factor (IGF-I) gene with circulating IGF-I concentration,growth,and fatness in swine[J]. Physiological Genomics,2006,31:236-243.
[13] Casas Carrillo E, Kirkpatrick B W, Prill Adams A, et al. Relationship of growth hormone and insulin-like growth factor-1 genotypes with growth and carcass traits in swine[J]. Animal Genetics,1997,28(2): 88-93.
