内蒙古白绒山羊erk2基因的克隆及表达模式分析
2013-10-31王彦凤吴曼琳王晓晶王婧李洋连梦瑶王志钢
王彦凤,吴曼琳,王晓晶,王婧,李洋,连梦瑶,王志钢
1 内蒙古大学生命科学学院,内蒙古 呼和浩特 010021
2 赤峰市医院,内蒙古 赤峰 024000
胞外信号调节激酶 (Extracellular regulated protein,Erk)是丝裂原激活蛋白激酶 (Mitogen activated protein kinase,MAPK)家族中的一个亚族,是Ras/Raf/MEK/ERK信号通路的重要成分,该信号通路介导着多种生物学效应,如细胞的增殖、分化,调节细胞周期[1]。该酶被激活后,成为磷酸化的ERK (p-ERK),p-ERK可以介导信号由胞浆向胞核传递,参与调节细胞的生长、发育、分化、分裂等多种生理过程,其异常表达在细胞的恶性转化和肿瘤的发生发展中起重要作用[2]。对erk基因的研究不仅可以区别物种间的遗传差异和进化特征,而且对动物细胞生长及个体发育过程中对外界影响因素的反应机制研究具有重要意义。早在 1990年,Boulton等发现了一种与酵母KSS1和FUS3激酶有着惊人相似的酶,被称为ERK1[3]。1991年,Boulton等继而发现了一类 ERK酶类,定义为ERK1、ERK2和ERK3等。ERK1和ERK2基因有90%的同源性,相对分子量分别为44 kDa和42 kDa[4]。Srinivasan等发现在鼠类血管发生中,Ras介导的信号通路是内皮细胞增殖和迁移的必要条件,缺乏ERK1/2会抑制初期内皮细胞的增殖[5]。Miltenberger等应用反义c-Raf1或抑制性Raf1使NIH3T3细胞的增殖受到干扰,而组成性激活的Raf1能加速增殖过程[6]。在许多肿瘤细胞中,都能检测到ERK信号通路的异常活化,Boucher等研究表明胰腺癌组织中的 ERK活性是癌组织周围正常组织的 2~18倍。Kang等研究表明,白血病细胞K562凋亡特性与ERK基础活性较高有关[7]。ERK通路与细胞周期的调控有着紧密的联系。有资料显示,ERK信号通路是多数生长因子、细胞因子调控靶细胞增殖的重要途径[8]。运用MEK抑制剂U0126阻断ERK通路,会抑制细胞周期蛋白D1 (CyclinD1)表达[9]。使用MEK抑制剂PD98059处理HT1080细胞,24 h后细胞仍然停留在G2/M期,使细胞周期阻滞[10]。内蒙古白绒山羊是经过长期自然选育形成的优良地方性绒肉兼用型品种,对营养的需求、利用与代谢有其特殊性。erk基因及其编码的蛋白已在一些动物中得到深入研究,但尚未有山羊erk基因研究方面的报道。建立有效地克隆内蒙古白绒山羊 erk2基因的 RT-PCR体系及定量RT-PCR体系,获得erk2基因编码区 cDNA序列及该基因的基本表达模式,为进一步研究该基因产物在机体发育中的作用提供条件。
1 材料与方法
1.1 相关载体及试剂
克隆载体 pMD19-T Vector、LA-Taq DNA polymerase、T4 DNA 连接酶以及 10×T4 DNA连接酶缓冲液、常用限制性内切酶及缓冲液、DNA分子量标准 DL2000、氨苄青霉素、卡那霉素、X-gal、IPTG、MiniBEST Plasmid Purification Kit Ver.2.0 (50次量)、RNAiso试剂盒 (TaKaRa,Cat. No. 3124)等均购自宝生物工程 (大连)有限公司 (TaKaRa);琼脂糖凝胶DNA回收试剂盒 (DP209-02离心柱型)购自TIANGEN BIOTECH (BEIJING)CO., LTD.;其余试剂为国产分析纯。
1.2 动物组织及细胞培养
内蒙古白绒山羊40 d的胎儿采自内蒙古白绒山羊种羊场,带回实验室后经原代培养纯化胎儿成纤维细胞,于液氮中保存。继代培养采用单层培养子代细胞,实验使用P1至P4代次之间的胎儿成纤维细胞。所用培养基为含有 10%FBS的 DMEM/F12,培养条件为 37 ℃、5%CO2、饱和湿度条件。
脾、睾丸、脑、肝、肺、乳腺和肾等组织样本取自新屠宰羊,迅速放入-196 ℃的液氮中冷冻后并置-80 ℃冰箱中保存备用。
1.3 总RNA的提取及反转录
按总 RNA提取试剂盒 (RNAiso Reagent)说明书的操作方法提取总RNA,琼脂糖凝胶电泳验证RNA的完整性,经紫外线测定定量后,用Oligod(T)18-Primers按照反转录酶AMV说明书的方法反转录合成第一链cDNA (cDNA1),作为下一步PCR扩增编码区片段的模板。
1.4 内蒙古白绒山羊erk2基因的分子克隆
从GenBank中查找出与绒山羊亲缘关系较近的牛、马、小鼠、大鼠、人等物种的erk2基因的CDS区序列,并进行同源比对。根据erk2基因的CDS区序列设计引物P1 (表1),其中包含的酶切位点为SacⅠ、NdeⅠ (上游),XhoⅠ、PstⅠ (下游)。预计扩增片段长度1 083 bp,以绒山羊胎儿成纤维细胞cDNA为模板进行PCR扩增,反应条件为:94 30 s℃,58 30 s℃,72 ℃16 s,30个循环。
扩增产物经琼脂糖凝胶电泳,回收特异性条带,将其克隆到pMD19-T载体,转化E. coli DH5α。蓝、白菌落筛选阳性克隆,重组质粒经PCR与酶切鉴定正确后送上海生工生物工程技术服务有限公司测序。
1.5 生物信息学方法
基因cDNA序列用NCBI上的BLAST进行序列比对 (http:www.ncbi.nlm.nih.gov/BLAST),开放阅读框 (ORF)预测采用 ORF Finder软件(www.ncbi.nlm.nih.gov/),蛋白质结构域分析采用SMART程序 (http://smart.embl.de/),蛋白质功能位点分析采用 Psite程序 (http://www.softberry.com),蛋白质亚细胞定位采用 PSORT程序 (http://psort.nibb.ac.jp),蛋白质等电点及分子量预测采用在线软件 (http://isoelectric.ovh.org/)分析,用 MEGA4.1软件进行同源性比对及构建进化树,利用瑞士模型工作区(Swiss-Model workspace)预测蛋白质二级结构(http:// swissmodel.expasy.org/)。
1.6 定量RT-PCR检测组织表达特异性
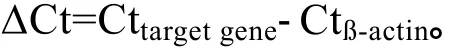
1.7 利用免疫组化方法检测组织表达特异性
收集内蒙古白绒山羊睾丸组织材料,按下列步骤处理:4%的多聚醛固定,酒精脱水,包埋,切片,酒精脱蜡,免疫动物血清封闭,一抗 (Rabbit anti-ERK2)4 ℃孵育过夜,PBS洗3次,生物素标记的二抗室温孵育 10 min,PBS洗 3次,依据试剂盒的说明利用 DAB(3,3'-diaminobenzidine tetrahydrochloride)染色,显微镜观察,照相,每种组织设一个负对照。
2 结果与分析
2.1 erk2基因克隆与序列分析
绒山羊erk2基因PCR扩增产物经电泳检测分析,在预期的1 083 bp处有特异性扩增条带 (图1)。重组质粒经PCR和酶切双重鉴定正确。序列测定结果表明,扩增出的CDS全长1 083 bp,编码360个氨基酸。通过Blast比对,内蒙古白绒山羊erk2基因的核苷酸序列与牛Bos taurus (BC133588.1)、狼 Canis lupus (NM_001110800.1)、人 Homo sapiens (NM_138957.2)、 野 猪 Sus scrofa(NM_001198922.1)、 小 鼠 Mus musculus(NM_011949.3)、 大 鼠 Rattus norvegicus(NM_053842.1)等物种的同源性分别为 96%、96%、92%、92%、89%和 89%,相应的氨基酸序列同源性均达到 96%以上,与牛的同源性更达到了 100%。序列提交 GenBank得到序列登记号为 JX569765。使用 MEGA4.1软件对上述序列进行比对构建进化树 (图 2),从图中可以发现,erk2是一个相对保守的基因,在各物种间的差异非常小。
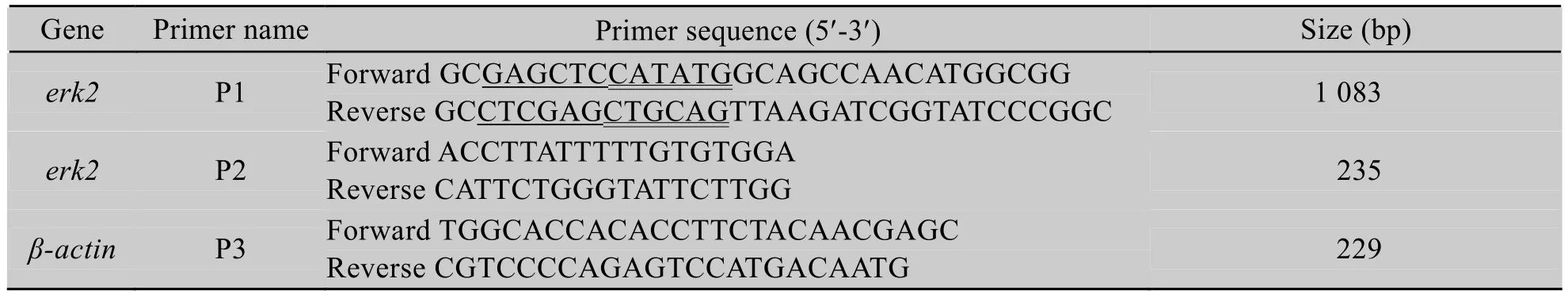
表1 本研究中用到的引物Table 1 Primers used in this study
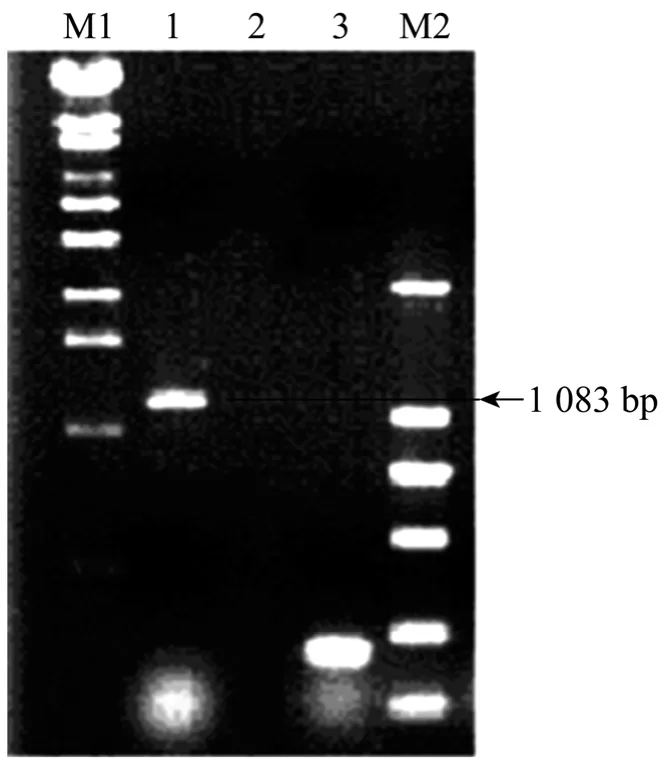
图1 erk2基因cDNA全长的PCR扩增Fig. 1 PCR product of erk2 gene. M1: λ-EcoT14 DNA marker; 1: PCR products; 2: negative control; 3: β-actin positive control; M2: DL 2 000 DNA marker.
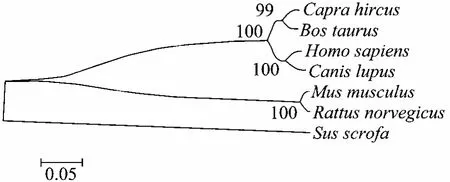
图2 各物种erk2核苷酸聚类分析Fig. 2 Alignment of several species of erk2 nucleotide sequence.
2.2 生物信息学分析
erk2基因编码一个由 360个氨基酸残基组成的蛋白质,该蛋白理论分子质量41.38 kDa,等电点 (pI)为 6.711,氨基酸组成中亮氨酸含量最高,为11.9%。SMART分析表明,在该蛋白质中 25~313位氨基酸是一个 S_TKc结构域(Serine/threonine kinase catalytic domain),即丝氨酸/苏氨酸激酶结构域 (图3)。已知的ERK蛋白是典型的丝/苏氨酸激酶,SMART分析结果进一步表明绒山羊erk2基因被正确克隆。
利用Swiss-Model workspace[12-14]预测蛋白质二级结构 (图 4)。对其编码的氨基酸序列进行 Psite分析表明,含 2个 N-糖基化位点、1个依赖于 cAMP/cGMP的蛋白激酶磷酸化位点、3个蛋白激酶c磷酸化位点、5个酪蛋白激酶Ⅱ磷酸化位点、2个 N-豆蔻酰化位点、2个异戊二烯基结合区 (CAAX box)、7个微体羧基端靶向信号、2个蛋白激酶ATP结合区标记及一个丝/苏氨酸蛋白激酶活性区域标记 (图5)。PSORT (k-NN prediction)程序预测其定位于细胞质中。
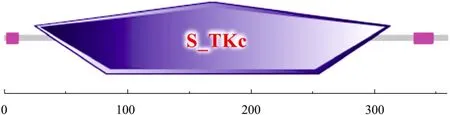
图3 erk2基因编码蛋白质SMART分析 (图中数字为氨基酸编号)Fig. 3 SMART analysis of the protein translated from erk2 gene (The number shows the location of each domain in protein).
图4 ERK2蛋白的3D图(9−358氨基酸序列)Fig. 4 Three-dimensional models of certain regions of ERK2 ( amino acid residues 9 to 358).
2.3 组织表达特异性检测
经定量RT-PCR显示检测,erk2基因在内蒙古绒山羊的心脏、脾、肾、肺、皮肤及乳腺组织中均有不同程度的表达。其中心脏、皮肤以及乳腺组织中mRNA丰度较高,脾、肾中的表达相对较低 (图 6)。利用免疫组化方法检测ERK2蛋白在睾丸中有特异性的表达 (图7)。
3 讨论
ERK属于丝裂原激活蛋白激酶 (Mitogen activated protein kinase,MAPK)家族中的一个亚族,包括ERK1与ERK2,后者研究更加深入。人类的 ERK2蛋白包括360个氨基酸残基,而鼠类包括 358个氨基酸残基[15]。该家族的蛋白一级结构具有两个典型的特征:一是TXY三肽基,被称为“磷酸化唇”或“活化唇”,ERK2的上游信号分子通过识别这一结构来活化ERK2,已报道的ERK2三肽基为“TEY”;另一个是丝氨酸/苏氨酸激酶催化活性的S_TKc结构域。本试验所克隆的内蒙古白绒山羊erk2基因ORF推测编码360个氨基酸残基,蛋白质经SMART程序分析具有这两种保守位点,与其他物种的 ERK2蛋白相符,证明内蒙古白绒山羊erk2基因被正确克隆。
Boulton等[2]通过 Northern blotting方法研究erk基因的组织特异性表达情况,发现其在小鼠的各种组织中均有表达,对于erk2来说,发现其在肌肉、胎盘以及大脑中的表达量最高。而且利用Western blotting的方法研究得到了相同的结果。本研究利用定量RT-PCR检测结果表明,erk2基因在内蒙古白绒山羊的心脏、脾、肾、肺、皮肤及乳腺组织中均有不同程度的表达。表明erk2基因可在多种组织中表达,进一步证明该基因在哺乳动物中具有保守的功能。

图5 内蒙古白绒山羊ERK2氨基酸序列与牛、小鼠及人的氨基酸序列比较及Psite活性位点分析 (1个依赖于cAMP/cGMP的蛋白激酶磷酸化位点 (氨基酸序列为KKLS),以双波浪线标记;3个蛋白激酶c磷酸化位点 (氨基酸序列为TLR、TAR及SNR),以灰色底纹标记;5个酪蛋白激酶Ⅱ磷酸化位点 (氨基酸序列为SAYD、SPFE、TLRE、TTCD 及 SQED),以方框标记;2个蛋白激酶 ATP 结区 (氨基酸序列为IGEGAYGMVCSAydnvnkvrVAIK、IGEGAYGMVCSAydnvnkvrvAIKK),以点下划线标记;1个丝/苏氨酸蛋白激酶活性区域 (氨基酸序列为 VLHRDLKPSNLLL),以灰色下划线标记;2个 N-糖基化位点 (氨基酸序列为NLSY、NTTC),以波浪线标记)Fig. 5 Alignment of protein ERK2 amino acids sequence between Inner Mongolia Cashmere Goat and Bos taurus,Mus musculus and Homo sapiens. The active sites are searched by Psite. A cAMP/cGMP-dependent protein kinase phosphorylation site (KKLS)is marked by double wavy line; Three protein kinase C phosphorylation sites(TLR/TAR/SNR)are marked by gray shadow. Five casein kinase phosphorylation sites(SAYD/SPFE/TLRE/TTCD/SQED)are marked by pane. Two protein kinase ATP-binding regions(IGEGAYGMVCSAydnvnkvrVAIK/IGEGAYGMVCSAydnvnkvrvAIKK)are marked by hidden underline; A Ser/Thr protein kinase activity region (VLHRDLKPSNLLL)is marked by gray underline. Two N-glycosylation site(NLSY/NTTC)are marked by wavy line.
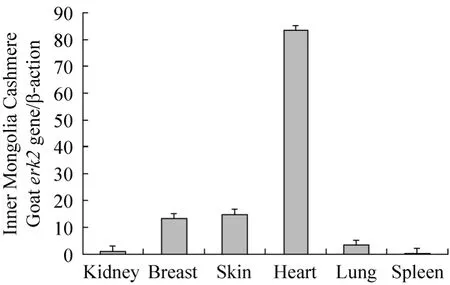
图6 定量RT-PCR检测内蒙古白绒山羊erk2基因组织表达特异性Fig. 6 Tissue-specific expression of erk2 gene of Inner Mongolia Cashmere Goat by quantitative RT-PCR.
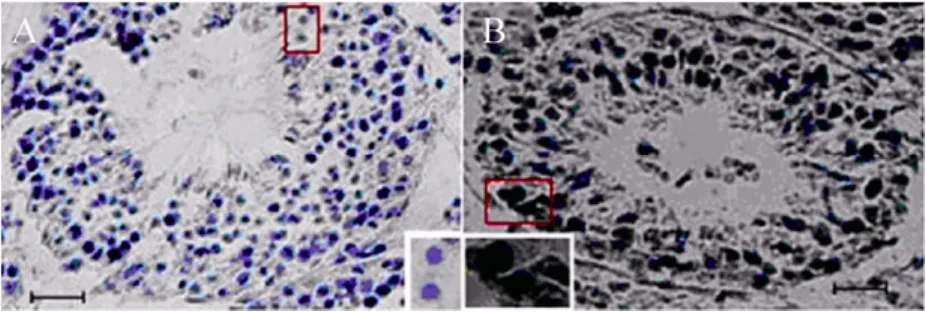
图7 睾丸中ERK2的表达Fig. 7 Expression of ERK2 protein in testis. (A)Negative control. (B)ERK2 is expressed in brown tissue cells. Scale bars=25 μm.
Ras/Raf/MEK/ERK信号通路可以感应多种信号分子,参与多种细胞进程,与多种疾病发生密切相关,如:脑损伤[16]、癌症[17]、肥胖[18]、糖尿病[19]以及炎症[20-21]等。近年来随着ERK的深入研究,一些新的功能也被相继报道。研究发现,长期的压力会降低小鼠海马体以及前额皮质中ERK的表达,而利用氟西汀处理会抵消这种反应[22],这一新功能的发现为抗抑郁症药物的研发带来了新的思路[23]。Zhu等研究发现,MAPK/ERK信号通路与血清饥饿引起的人成骨细胞凋亡相关[24]。
内蒙古白绒山羊 (Inner Mongolia Cashmere Goat)是经过长期的自然选择和人工选育而形成的优良产绒品种,但近年来由于草场退化,种群数量增加,舍饲比例加大,导致其生产性能降低,这其中机体对营养和能量信号刺激的敏感性起着重要的作用,但机制尚不清楚。多项研究显示ERK通路在营养摄入与调节过程中发挥着重要的作用,如ERK通路可以调节氨基酸刺激细胞后的信号传递过程[25];动物食料中缺乏必需氨基酸时,会激发ERK信号通路而不是mTOR信号通路[26];ERK信号通路可以调节氨基酸运输从而调节细胞的增殖[27]。本研究正确克隆了内蒙古白绒山羊erk2基因并对其基本表达模式进行了分析,这些结果将为进一步研究ERK在绒山羊细胞生长和早期胚胎发育中的功能及发挥作用的分子机制提供条件。
[1]SL Abrams, Steelman LS, Shelton JG, et al. The Raf/MEK/ERK pathway can govern drug resistance, apoptosis and sensitivity to targeted therapy. Cell Cycle, 2010, 9(9): 1781−1791.
[2]Roskoski R Jr. ERK1/2 MAP kinases: structure,function, and regulation. Pharmacol Res, 2012,66(2): 105−43.
[3]Boulton TG, Nye SH, Robbins DJ, et al. ERKs: a family of protein-serine/threonine kinases that are activated and tyrosine phosphorylated in response to insulin and NGF. Cell, 1991, 65(4): 663−675.
[4]Boulton TG, Yancopoulos GD, Gregory JS, et al.An insulin-stimulated protein kinase similar to yeast kinases involved in cell cycle control.Science, 1990, 249(4964): 64−67.
[5]Srinivasan R, Zabuawala T, Huang H, et al. Erk1 and Erk2 regulate endothelial cell proliferation and migration during mouse embryonic angiogenesis. PLoS ONE, 2009, 4(12): e8283.
[6]Miltenger RJ, Cortner J, Famham PJ. An inhibitory Raf-1 mutant suppresses expression of a subset of v-raf-activated genes. J Biol Chem,1993, 268(21): 15674−15680.
[7]Kang CD, Yoo SD, Hwang BW, et al. The inhibition of ERK/MAPK not the activation of JAK/SAPK is primarily required to induce apoptosis in chronic myelodenous leukemia K562 cell. Leuk Res, 2000, 24(6): 527−534.
[8]Lv C, Sun W, Sun H, et al. Asperolide A, a marine-derived tetranorditerpenoid, induces G2/M arrest in human NCI-H460 lung carcinoma cells,is mediated by p53-p21 stabilization and modulated by Ras/Raf/MEK/ERK signaling pathway. Mar Drugs, 2013, 11(2): 316−331.
[9]Zhao Y, Lv M, Lin H, et al. Rho-associated protein kinase isoforms stimulate proliferation of vascular smooth muscle cells through ERK and induction of cyclin D1 and PCNA. Biochem Biophys Res Commun, 2013, 432(3): 488−493.
[10]Zhang YX, Kong CZ. The role of mitogenactivated protein kinase cascades in inhibition of proliferation in human prostate carcinoma cells by raloxifene: an in vitro experiment. Zhonghua Yi Xue Za Zhi, 2008, 88(4): 271−275.
[11]Fang MX, Zhang W, Hu YS, et al. Cloning,expression and polymorphism analysis of pig erk2 gene for reproduction traits. Sci Agri Sin, 2011,44(1): 210−217 (in Chinese).方梅霞, 张伟, 胡永胜, 等. 猪繁殖候选基因erk2 的克隆、表达及基因多态分析. 中国农业科学, 2011, 44(1): 210−217.
[12]Arnold K, Bordoli L, Kopp J, et al. The SWISS-MODEL workspace: a web-based environment for protein structure homology modeling. Bioinformatics, 2006, 22(2): 195−201.
[13]Schwede T, Kopp J, Guex N, et al. SWISSMODEL: an automated protein homologymodeling server. Nucleic Acids Res, 2003, 31(13):3381−3385.
[14]Guex N, Peitsch MC. SWISS-MODEL and the Swiss-PdbViewer: an environment for comparative protein modeling. Electrophoresis, 1997, 18(15):2714−2723.
[15]Roskoski Jr R. ERK1/2 MAP kinases: structure,function, and regulation. Pharmacol Res, 2012,66(2): 105−143.
[16]Kim EK, Choi EJ. Pathological roles of MAPK signaling pathways in human diseases. Biochim Biophys Acta, 2010, 1802(4): 396−405.
[17]Tidyman WE, Rauen KA. The RASopathies:developmental syndromes of Ras/MAPK pathway dysregulation. Curr Opin Genet Dev, 2009, 19(3):230−236.
[18]Tanti JF, Jager J. Cellular mechanisms of insulin resistance: role of stress regulated serine kinases and insulin receptor substrates (IRS)serine phosphorylation. Curr Opin Pharmacol, 2009,9(6): 753−762.
[19]Montagut C, Settleman J. Targeting the RAF-MEK-ERK pathway in cancer therapy.Cancer Lett, 2009, 283(2): 125−34.
[20]Chico LK, Van Eldik LJ, Watterson DM. Targeting protein kinases in central nervous system disorders. Nat Rev Drug Discov, 2009, 8:892−909.
[21]Muslin AJ. MAPK signalling in cardiovascular health and disease: molecular mechanisms and therapeutic targets. Clin Sci, 2008(115): 203−218.
[22]First M, Gil-Ad I, Taler M, et al. The effects of fl uoxetine treatment in a chronic mild stress rat model on depression-related behavior, brain neurotrophins and ERK expression. J Mol Neurosci, 2011, 45(2): 246−255.
[23]Chandran A, Iyo AH, Jernigan CS, et al. Reduced phosphorylation of the mTOR signaling pathway components in the amygdala of rats exposed to chronic stress. Prog Neuropsychopharm Biol Psychi, 2013, 40(10): 240−245.
[24]Zhu X, Jiang Y, Shan PF, et al. Vaspin attenuates the apoptosis of human osteoblasts through ERK signaling pathway. Amino Acids, 2013, 44(3):961−968.
[25]Casas-Terradellas E, Tato I, Bartrons R, et al.ERK and p38 pathways regulate amino acid signaling. Biochim Biophy Acta, 2008 1783(12):2241−2254.
[26]Hao S, Ross-Inta CM, Gietzen DW. The sensing of essential amino acid def i ciency in the anterior piriform cortex, that requires the uncharged tRNA/GCN2 pathway, is sensitive to wortmannin but not rapamycin. Pharmacol Biochem Behav,2010, 94(3): 333−340.
[27]Bulus N, Feral C, Pozzi A, et al. CD98 increases renal epithelial cell proliferation by activating MAPKs. PLoS ONE, 2012, 7(6): e40026.
