太湖日本沼虾与秀丽白虾的食性与食物碳源分析
2013-08-11温周瑞湖北省水产科学研究所湖北武汉430071中国科学院水生生物研究所东湖湖泊生态系统试验站淡水生态与生物技术国家重点实验室湖北武汉430072
温周瑞 (湖北省水产科学研究所,湖北 武汉430071;中国科学院水生生物研究所,东湖湖泊生态系统试验站,淡水生态与生物技术国家重点实验室,湖北 武汉430072)
谢 平 ( 中国科学院水生生物研究所,东湖湖泊生态系统试验站,淡水生态与生物技术国家重点实验室,湖北武汉430072)
日本沼虾 (Macrobrachium nipponense)和秀丽白虾 (Exopalaemon modestus)是湖泊生态系统的重要成员,不仅是食物链的重要环节,也是人类喜爱的水产品。太湖盛产日本沼虾与秀丽白虾,其产量历年平均在50万~75万kg左右,约占全湖渔业产量的5%~10%[1]。有关日本沼虾和秀丽白虾食性的研究报道不多,赖伟等[2]、施炜钢等[3]曾经研究过太湖秀丽白虾的食性,但太湖日本沼虾的食性尚未见有研究报告。刘军等[4]研究了武湖日本沼虾的食性。因此,有必要开展太湖这2种虾食性的研究。
现有的关于日本沼虾、秀丽白虾食性的研究均采用胃含物分析的方法。该法是传统的水生动物食性研究方法,只能了解捕捞前瞬间的食物组成,而且胃含物中易于消化的食物首先被消化,残留下来的多为不易消化的食物,所以会高估难消化食物在胃含物中的比例。天然发生的稳定性同位素被用于研究水生动物的食性,Peterson[5]认为同位素分析技术与胃含物分析相结合能更有效地了解动物的食性。本研究将稳定性碳同位素分析与胃含物分析相结合,对日本沼虾、秀丽白虾的食性与食物碳源进行分析。
1 材料与方法
1.1 研究区概况
研究区位于太湖梅梁湾、贡湖湾。梅梁湾系太湖北部一个较大的湖湾,地处无锡市郊,面积约为123.88km2,平均水深2m左右,属于典型的富营养化湖区[6]。梅梁湾基本没有水草生长[7],仅在人类活动影响较小的东岸三国城至康山渚一带有少量的荇菜、菱等分布[8]。贡湖湾位于太湖东北部,面积147km2,平均水深2.0m左右。采样期间分布有水生植物,如微齿眼子菜 (Potamogeton maackianus)、马来眼子菜 (Potamogeton malaianus)、穗花狐尾藻 (Myriophyllum spicatum)、伊乐藻 (Elodea nuttallii)、苦草 (Vallisneria natans)、金鱼藻 (Ceratophyllum demersum)、黑藻 (Hydrilla verticillata)、菹草 (Potamogeton crispus)及浮叶植物荇菜 (Nymphoides peltatum)和菱 (Trapaquadrispinos)等,优势种类为马来眼子菜、微齿眼子菜[9-10]。
1.2 食性分析
胃含物分析:2005年1月至2005年11月每月在梅梁湾和贡湖湾3个采样点 (图1)用虾拖网拖取活虾,当场冷藏杀死,带回室内,测定体长、体重,取出胃,放入离心管中加入10%甲醛固定保存。胃含物在显微镜下鉴定种类,并统计出现频率。
食物碳源分析:取虾腹部肌肉,用蒸馏水淋洗,60℃烘干至恒重干燥保存。进行同位素分析前,用碾钵碾成粉末,用质谱仪测定虾的δ13C。
采用质量平衡模型[11]计算底栖和浮游食物网对日本沼虾和秀丽白虾的营养贡献:


图1 梅梁湾与贡湖湾采样点分布图
式中,δ13Cshrimp、δ13Cpelagic、和δ13Cbenthic分别代表日本沼虾或秀丽白虾δ13C、浮游食物网基准δ13C和底栖食物网基准δ13C。由于初级生产者,如藻类,δ13C值发生较大的时空变化[12-13],作为基准会影响对高营养级的评价和虾δ13C、δ15N的比较。本研究使用寿命较长的初级消费者螺、蚌分别作为底栖和浮游食物网基准,这些长寿命大型无脊椎动物受初级生产者同位素组成的季节波动影响较小[14-16]。
2 结果与分析
2.1 食物组成

表1 日本沼虾食物出现率 %
梅梁湾和贡湖湾的日本沼虾、秀丽白虾胃含物中共镜检出浮游植物、原生动物、轮虫、枝角类、桡足类、水生寡毛类、昆虫、鱼、植物碎片、有机碎屑等10大类,其中浮游植物有绿藻门、硅藻门、蓝藻门、隐藻门等。
贡湖湾日本沼虾食物中出现率大小顺序依次为:有机碎屑>绿藻门>枝角类>硅藻门>植物碎片>蓝藻门>轮虫 (表1)。出现频率分别高达100.0%、75.2%、67.6%、63.4%、52.4%、44.1%。绿藻门出现频率最高的是盘星藻、栅藻、卵囊藻,蓝藻门中微囊藻出现率占蓝藻门的93.2%;象鼻蚤出现率在枝角类中最高,占枝角类的75.4%;枝角类出现率3#点最高。水生植物出现率1#点达到83.3%,2#点为58.3%,3#点为16.3%。
梅梁湾日本沼虾食物中出现率大小顺序依次为:有机碎屑100.0%>枝角类69.9%>硅藻门57.5%>轮虫49.3%>绿藻门47.3%>蓝藻门41.1%>水生植物11.0% (表1)。绿藻门出现频率最高的是栅藻、卵囊藻、盘星藻;蓝藻门中微囊藻出现率占蓝藻门的96.6%;枝角类中象鼻蚤出现率最高,占枝角类的75.4%;水生植物的出现率为11.0%,1#点水生植物出现率最高,为17.7%。
贡湖湾秀丽白虾食物出现率从大到小的顺序依次是有机碎屑>枝角类>绿藻门>硅藻门>轮虫>蓝藻门>水生植物等。枝角类最高出现率100.0%,平均69.6%,其中象鼻蚤出现率最高达94.7%,平均60.8%。蓝藻门中基本都是微囊藻,绿藻门主要是盘星藻、栅藻等,硅藻门主要是直链藻、桥弯藻、脆杆藻等;水生植物出现率1#点最高 (43.2%),3#点最低 (10.5%),平均为28.4% (表2)。
梅梁湾秀丽白虾食物出现率从大到小的顺序依次是有机碎屑>枝角类>硅藻门>轮虫>蓝藻门>绿藻门>水生植物等。枝角类中象鼻蚤出现率最高平均达63.6%;蓝藻门中基本都是微囊藻;绿藻门主要是盘星藻、栅藻、小球藻等;硅藻门主要是直链藻、小环藻、脆杆藻等;水生植物出现率2#点最高,3#点没有水生植物,平均为12.1% (表2)。
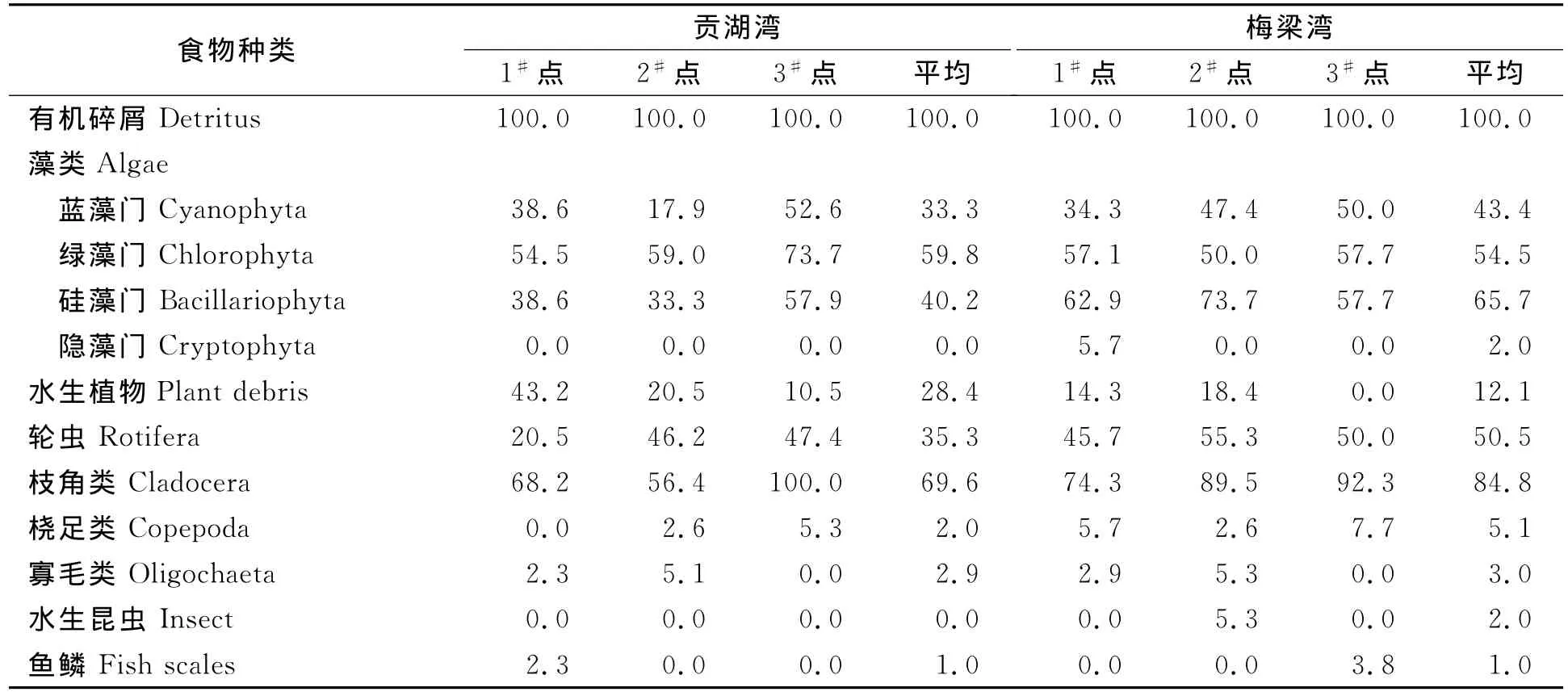
表2 秀丽白虾食物出现率%
日本沼虾的食物组成存在月度变化。贡湖湾日本沼虾食物中有机碎屑各月出现率均为100.00%;水生植物3月至11月出现率较高,1月较低;枝角类出现率除3月稍低,5月、9月、11月高;微囊藻5月至11月出现率均高;其他食物基本在3月~11月较高,1月较低 (表3)。

表3 贡湖湾日本沼虾食物月度出现率 %
梅梁湾日本沼虾食物中有机碎屑各月出现率均为100.00%;水生植物7月、11月出现率较高,1月较低;枝角类出现率1月较低,5月、9月、11月很高;微囊藻7月至11月出现率均高;其他食物基本在3月~11月较高,1月较低 (表4)。

表4 梅梁湾日本沼虾食物月度出现率 %
秀丽白虾的食物组成出现明显月度变化。贡湖湾秀丽白虾食物中有机碎屑各月出现率均为100.00%;水生植物3月至11月出现率较高,1月较低;枝角类出现率1月、3月较低,5月、7月、9月、11月很高;微囊藻除3月较低,其余几个月较高;其他食物出现率基本在3月~11月较高,1月较低 (表5)。
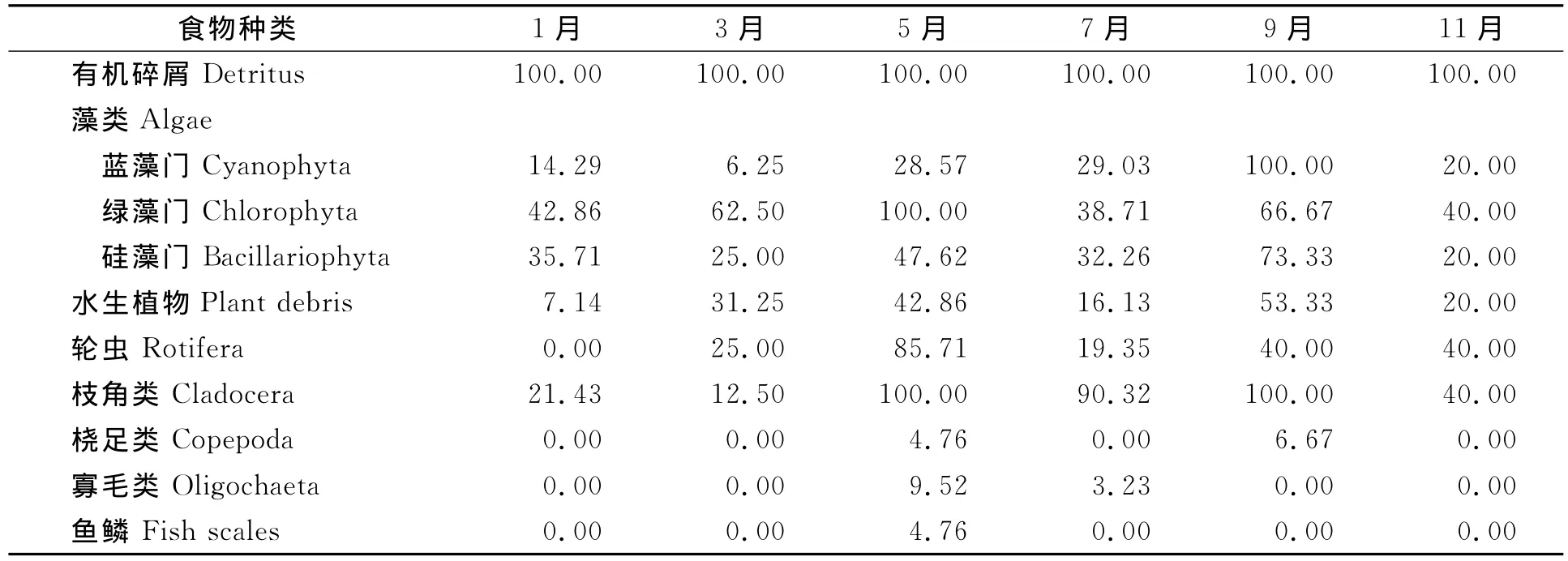
表5 贡湖湾秀丽白虾食物月度出现率 %
梅梁湾秀丽白虾食物中有机碎屑各月出现率均为100.00%;水生植物1月、9月没有;枝角类出现率1月为0,5月、7月、9月、11月很高;微囊藻除1月较低,其余几个月较高;其他食物基本在3月~11月较高,1月基本没有食物 (表6)。
2.2 碳源分析
同位素测定结果见表7。梅梁湾日本沼虾δ13C值1#点与2#点差异不显著 (P>0.05),1#点与3#点、2#点与3#点存在极显著差异 (P<0.01);说明梅梁湾1#点与2#点日本沼虾食物碳源相近,1#点与3#点、2#点与3#点日本沼虾食物来源有差异。3个采样点之间秀丽白虾δ13C均不存在显著差异 (P>0.05),表明3个点秀丽白虾的食物碳源相似,可能与秀丽白虾营游泳生活,活动范围较广有关。贡湖湾3个采样点之间日本沼虾δ13C差异极显著 (P<0.01),秀丽白虾δ13C1#点与2#点、1#点与3#点差异极显著 (P<0.01),2#点与3#差异不显著 (P>0.05)。
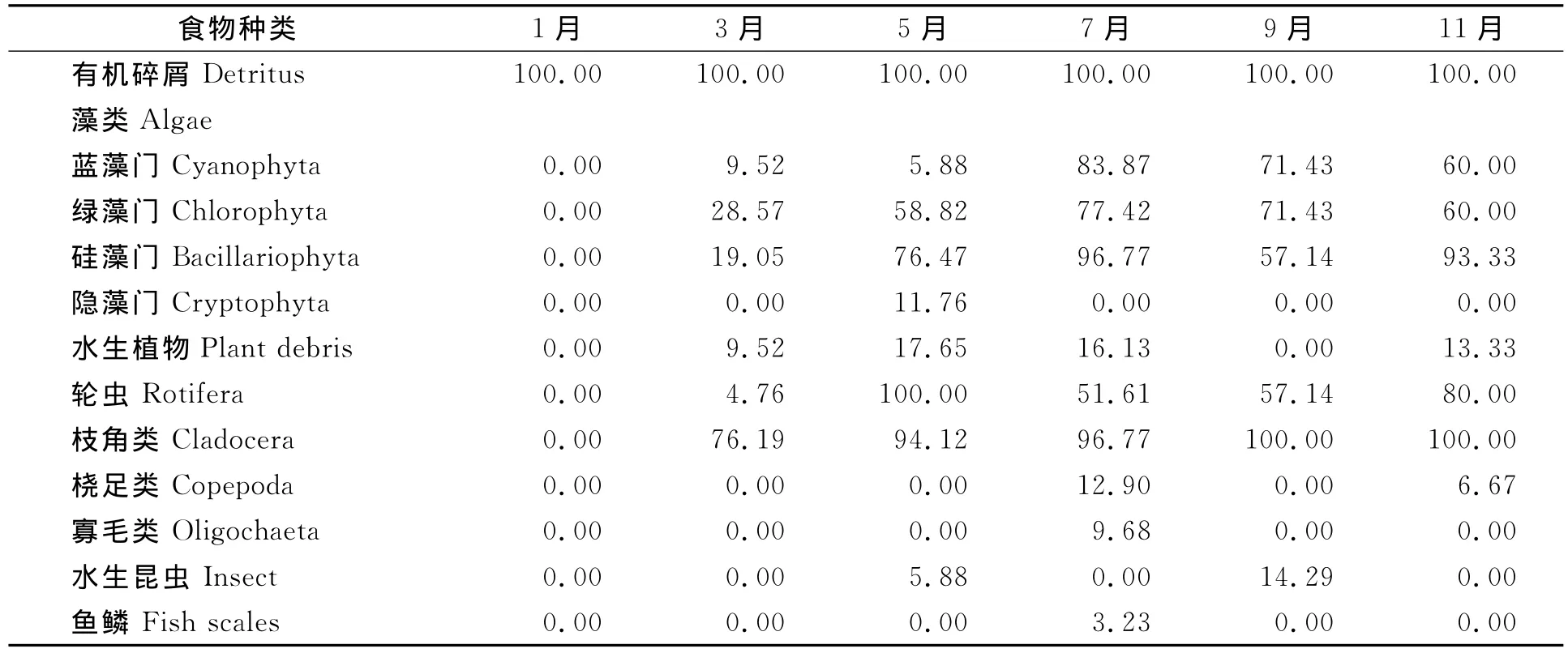
表6 梅梁湾秀丽白虾食物月度出现率 %
梅梁湾日本沼虾和秀丽白虾δ13C分别为(-25.27±1.34)‰ (n=282)、 (-26.64±1.97)‰ (n=236),两者有极显著差异 (P<0.001),说明日本沼虾与秀丽白虾食物碳源有差异。贡湖湾日本沼虾与秀丽白虾δ13C分别 为 (- 24.86 ± 1.69)‰ (n=282)、(-24.75±1.82)‰(n=217),两者没有显著差异(P=0.52)。梅梁湾与贡湖湾日本沼虾δ13C分 别 为 (-25.27±1.34)‰ (n=282)、(-24.86±1.69)‰(n=282),两者有极显著差异 (P<0.001)。梅梁湾与贡湖湾秀丽白虾δ13C分别为 (-26.64±1.97)‰ (n=236)、(-24.75±1.82)‰ (n=217),两者有极显著差异 (P<0.001),说明2个湖区这2种虾食物碳源差异显著。
利用日本沼虾、秀丽白虾、铜锈环棱螺、5种蚌的碳同位素年平均值,通过混合模型计算出底栖碳源贡献率 (表8)。可以看出,贡湖湾底栖碳源贡献率高于梅梁湾,梅梁湾日本沼虾底栖碳源贡献率高于秀丽白虾,且沿岸带的比敞水区的高;贡湖湾几乎全部来自底栖碳源,2种虾基本没有区别。

表7 太湖日本沼虾、秀丽白虾δ13 C值

表8 底栖食物对日本沼虾和秀丽白虾食物的贡献率
3 讨论
3.1 食性
经周年调查发现,梅梁湾、贡湖湾日本沼虾和秀丽白虾食物种类主要包括有机碎屑、浮游植物、水生植物、浮游动物、底栖动物、鱼类等,与严生良等[17]、刘军等[4]、胡廷尖等[18]报道的基本一致。不同食物出现率因虾的种类、季节、采样地点而不同,这种变化与环境中食物丰富度有关。从季节上来看,一般食物的出现率均是春夏秋3季度出现率较高,冬季出现率较低,这与饵料生物的周年生长规律是一致的,就是说食物出现率的高低与食物易得性有关,当环境中食物较丰富时,其在胃含物中出现率也较高。从不同湖区和采样点来看,食物出现率也很好地反映了环境中食物分布的差异。贡湖湾与梅梁湾相比,无论是日本沼虾还是秀丽白虾,水生植物出现率均较高,贡湖湾日本沼虾水生植物出现率51.72%,梅梁湾为10.96%;贡湖湾秀丽白虾水生植物出现率28.43%,梅梁湾为12.12%。贡湖湾日本沼虾浮游植物出现率高于梅梁湾,但秀丽白虾浮游植物出现率则比梅梁湾低。从叶绿素含量来看,贡湖湾浮游植物生物量比梅梁湾要低[19],但日本沼虾浮游植物出现率较高,可能与刘军[4]认为浮游植物仅为日本沼虾摄食时所带入情况有关。贡湖湾秀丽白虾水生植物出现率比日本沼虾要低,所以带入的浮游植物也少,故浮游植物出现率较低。
不同采样点之间食物出现率也有差异。贡湖湾1#点水草覆盖率较高,主要种类有微齿眼子菜、金鱼藻、马来眼子菜、苦草等沉水植物,2#点有零星沉水植物分布,3#点位于沿岸带,岸边有芦苇,少量马来眼子菜分布。1#点水质明显好于其他两个点,2#点次之,3#点水体营养水平最高,浮游动植物生物量最大。1#、2#、3#点日本沼虾食物中水生植物出现率分别是83.3%、58.3%、16.3%,秀丽白虾食物中水生植物出现率分别为43.2%、20.5%、10.5%。可见胃含物中水生植物出现率随其丰度的增大而增大。贡湖湾日本沼虾、秀丽白虾食物中枝角类出现率的大小顺序是3#点>2#点>1#点,与水体中浮游甲壳类密度大小顺序一致,3#点、2#点、1#点浮游甲壳类密度分别为465.8、281.4、198.8ind/L[20]。梅梁湾1#点位于沿岸带,有少量水生植物分布,2#点和3#点位于敞水区,基本没有水生植物生长,但日本沼虾和秀丽白虾胃含物中仍有水生植物出现,日本沼虾1#点、2#点、3#点胃含物中水生植物出现率分别是17.65%、13.04%、2.04%;秀丽白虾1#点、2#点、3#点胃含物中水生植物出现率分别是14.29%、18.42%和0。可能是少量水草漂浮至敞水区被虾类摄食所致。日本沼虾1#点、2#点、3#点胃含物中枝角类出现率分别是60.78%、69.57%、79.59%,秀丽白虾1#点、2#点、3#点胃含物中枝角类出现率分别是74.29%、89.47%、92.31%。梅梁湾1#点、2#点、3#点浮游甲壳类的密度分别为168.4、356.8、438.1ind/L[20],出现率的高低与密度大小顺序是一致的,说明浮游甲壳类的密度越大,虾类对其摄食概率也越高。施炜钢等[3]也报道太湖秀丽白虾胃内浮游动物出现频率基本上与常年太湖浮游动物优势种群相吻合。
值得指出的是,日本沼虾和秀丽白虾胃含物中微囊藻的出现率较高,梅梁湾日本沼虾和秀丽白虾胃含物中微囊藻出现率最高分别为73.3%、83.9%,贡湖湾日本沼虾和秀丽白虾胃含物中微囊藻出现率最高分别为67.7%、100%。微囊藻含有藻毒素,虾类摄入微囊藻后体内会累积微囊藻毒素,人们食用这样的虾类会对身体造成危害[21],因此,对太湖虾类的食用安全必须予以高度关注。
3.2 食物碳源
混合模型计算结果,梅梁湾1#、2#、3#点底栖食物碳源对日本沼虾贡献率分别是100.00%、86.33%、84.29%,对秀丽白虾贡献率分别是69.07%、56.32%、51.50%,说明沿岸带底栖食物碳源贡献率比敞水区高,日本沼虾食物碳源中底栖食物贡献率比秀丽白虾高,故梅梁湾日本沼虾与秀丽白虾δ13C有显著差异。贡湖湾1#、2#、3#点底栖食物碳源对日本沼虾和秀丽白虾贡献率均为100.00%,故贡湖湾日本沼虾与秀丽白虾δ13C差异不显著。梅梁湾与贡湖湾之间日本沼虾δ13C、秀丽白虾δ13C均有显著差异,与梅梁湾与贡湖湾之间2种虾的食物来源有关。
[1]朱清顺,苗玉霞 .江苏淡水虾类及其渔业 [J].动物学杂志,1990,25(3):8-11.
[2]赖 伟,邓雪怀,陈炳良,等.太湖秀丽白虾种群及生殖习性研究 [J].华东师范大学学报 (自然科学版),1983,(3):81-87.
[3]施炜纲,严小梅,邴旭文.太湖秀丽白虾生物学与食性的研究 [J].湖泊科学,1995,7(1):69-76.
[4]刘 军,龚世园,何绪刚,等.武湖日本沼虾食性的研究 [J].淡水渔业,2005,35(1):25-28.
[5]Peterson B.Stable isotopes as tracers of organic matter input and transfer in benthic food webs:a review [J].Acta oecologica,1999,20:479-487.
[6]龚绍琦,黄家柱,李云梅,等.太湖梅梁湾水质参数空间变异及合理取样数目研究 [J].地理与地理信息科学,2006,22(2):50-54.
[7]雷泽湘,徐德兰,黄沛生,等.太湖沉水和浮叶植被及其水环境效应研究 [J].生态环境,2006,15(2):239-243.
[8]杨龙元,梁海棠,胡维平,等.太湖北部滨岸区水生植被自然修复观测研究 [J].湖泊科学,2002,14(1):60-66.
[9]金相灿,颜昌宙,许秋瑾.太湖北岸湖滨带观测场水生植物群落特征及其影响因素分析 [J].湖泊科学,2007,19(2):151-157.
[10]刘伟龙,胡维平,陈永根,等.西太湖水生植物时空变化 [J].生态学报,2007,27(1):159-170.
[11]Xu J,Zhang M,Xie P.Size-related shifts in reliance on benthic and pelagic food webs by lake anchovy [J].Ecoscience,2007,14(2):170-177.
[12]Xu J,Xie P,Zhang M,et al.Variation in stable isotope signatures of seston and a zooplanktivorous fish in a eutrophic Chinese lake[J].Hydrobiologia,2005,541:215-220.
[13]Zhou Q,Xu J,Xie P,et al.Seasonal variations in stable isotope ratios of two biomanipulation fishes and seston in a large pen culture in hypereutrophic Meiliang Bay,Lake Taihu [J].Ecological Engineering,2009,35:1603-1609.
[14]Cabana G,Rasmussen J B.Comparisons of aquatic food chains using nitrogen isotopes[J].Proceedings of the National Academy of Sciences of the United States of America,1996,93:10844-10847.
[15]Vander Zanden M J,Rasmussen J B.Primary consumerδ13C andδ15N and the trophic position of aquatic consumers[J].Ecology,1999,80:1395-1404.
[16]Post D M.Using stable isotopes to estimate trophic position:models,methods and assumptions [J].Ecology,2002,83 (3):703-718.
[17]严生良,金德沂.太湖的青虾 [J].水产科技情报,1981,(6):18-19.
[18]胡廷尖,周志明,黄鲜明,等.秀丽白虾生物学特性及资源开发的初探 [J].水利渔业,2001,21(2):7-8.
[19]Liu Y Q,Xie P,Zhang D W,et al.Seasonal Dynamics of Microcystins with Associated Biotic and Abiotic Parameters in Two Bays of Lake Taihu,the Third Largest Freshwater Lake in China[J].Bull Environ Contam Toxicol,2008,80:24-29.
[20]温周瑞,谢 平.太湖梅梁湾、贡湖湾水质与浮游甲壳动物密度季节变化 [J].长江大学学报 (自然科学版),2013,10(17):36-41.
[21]Chen J,Xie P.Tissue distributions and seasonal dynamics of the hepatotoxic microcystins-LR and-RR in two freshwater shrimps,Palaemon modestus and Macrobrachium nipponense,from a large shallow,eutrophic lake of the subtropical China [J].Toxicon,2005,45:615-625.
