油菜田日本看麦娘对高效氟吡甲禾灵抗性的生理应答
2013-08-02陈宏州杨敬辉张文文庄义庆
陈宏州, 杨敬辉, 张文文, 陈 露, 庄义庆
(江苏丘陵地区镇江农业科学研究所,江苏 句容 212400)
在中国,冬油菜约占油菜总种植面积的90%, 主要分布于长江流域,且多为稻茬油菜田,这就导致了喜湿性禾本科杂草——日本看麦娘(Alopecurus japonicus)危害严重而又难以防除[1]。目前,油菜田日本看麦娘仍以化学防治为主,防除药剂主要为芳氧基苯氧丙酸(Aryloxyphenoxypropionate,AOPP)类和环已烯酮(Cyclohexanedione,CHD)类除草剂,其中AOPP类除草剂——高效氟吡甲禾灵(Haloxyfop-R-methyl)为主导药剂[1]。近年来,油菜田日本看麦娘发生比重不断上升,危害日益严重,农民为了达到防除杂草的目的,不断增加施药量和施药次数,使除草剂对杂草的选择压逐年上升。由于AOPP类和CHD类除草剂作用靶标单一,长时间连续单一使用,导致防效下降,甚至有产生明显抗性的报道。杨彩宏等[2]、潘以楼等[3]报道油菜田日本看麦娘对高效氟吡甲禾灵产生了明显的抗药性,且抗性程度与用药年限的长短有一定的关系。陈宏州等[4]报道在江苏省句容市部分油菜田,10.8%高效氟吡甲禾灵EC按推荐剂量的3倍使用,1次用药后60 d,对日本看麦娘的株防效为负值。
AOPP类和CHD类除草剂作用靶标都是乙酰辅酶A羧化酶(Acetyl coenzyme A carboxylase,ACCase)。研究人员发现杂草体内ACCase被修饰使杂草对多数AOPP类和CHD类除草剂的敏感性降低,从而产生抗药性,而这种抗性多为单个基因控制[5-6]。这种抗药性机制已在澳大利亚的抗性早熟禾、羊茅属杂草和瑞士黑麦草中得到证实[7]。中国在杂草抗药性方面的研究,与国外相比相对滞后,且多数集中于抗性杂草的鉴定方法上。近年来,为了延缓抗性的发展,越来越多的研究人员开始更加注重抗性产生的分子机理以及生理机制研究。黄世霞等[8]采用双向等位基因特异性PCR研究不同抗性生物型看麦娘对ACCase抑制剂类除草剂稀禾啶和高效氟吡甲禾灵产生交互抗性的分子基础,证实了靶标酶ACCase可能存在多个突变位点而产生不同模式的抗药性,同时也表明AOPP类和CHD类除草剂的作用位点是有差异的。韩瑞娟等[9]研究了抗性和敏感日本看麦娘体内细胞色素P450还原酶和谷胱甘肽-S-转移酶的活性差异,结果表明代谢酶活性增强是日本看麦娘对高效氟吡甲禾灵产生抗性的机理之一。
目前,关于油菜田日本看麦娘对高效氟吡甲禾灵产生抗性的生理机制还鲜有报道。本研究以种子生物测定法鉴定了3个油菜田日本看麦娘种群对高效氟吡甲禾灵的抗性水平,然后进行不同抗性生物型日本看麦娘喷施高效氟吡甲禾灵后在生理水平上的应答研究,以期为抗性日本看麦娘的防除与治理提供科学依据。
1 材料与方法
1.1 供试种子
2011年5月,在高效氟吡甲禾灵使用15~20年的江苏省句容市和丹阳市油菜田采集成熟的日本看麦娘种子,获取2个日本看麦娘种群,分别命名为JR种群和DY种群[10]。野生日本看麦娘种群(CZ种群)由南京农业大学植物保护学院董立尧教授提供,该种群采自安徽滁州休闲田[2]。
1.2 供试药剂
94.9%高效氟吡甲禾灵原药(江苏扬农化工有限公司),用适量二甲苯溶解后,加入10%的吐温-80,配制成10 000 μg/ml的母液置于4℃冰箱中备用;10.8%高效氟吡甲禾灵EC为美国陶氏益农公司产品。
1.3 日本看麦娘对高效氟吡甲禾灵的抗性水平检测
在预试验基础上,以种子生物测定法[11],将50粒经预先催芽且芽长约1 mm的日本看麦娘种子放在铺有一层滤纸的9 cm培养皿内,加入5 ml浓度分 别 为 0 μg/ml、80.000 00 μg/ml、40.000 00 μg/ml、 20.000 00 μg/ml、 10.000 00 μg/ml、5.000 00 μg/ml、2.500 00 μg/ml、1.250 00 μg/ml、0.625 00 μg/ml、0.312 50 μg/ml和0.156 25 μg/ml的高效氟吡甲禾灵药液,加盖确保种子处于湿润状态,置于光照培养箱内培养(光照/黑暗为12 h/12 h;温度:白天18℃,晚上15℃),每处理重复4次。药剂处理10 d后,测量芽长,计算药剂对杂草种子芽长的抑制率。
采用DPS数据处理软件,计算药剂对不同日本看麦娘种群的毒力回归方程、相关系数、EC50及其95%置信限,并计算相对抗性倍数。相对抗性倍数=不同地区日本看麦娘种群的EC50/敏感日本看麦娘种群的EC50[3]。
1.4 不同抗性生物型的生理指标测定
1.4.1 材料准备 将壤质菜园土与沙按3∶1混匀后,装入直径为9 cm的塑料盆钵内,然后将适量不同抗性生物型的日本看麦娘种子分别播种于盆钵内,并置于光照培养箱内培养,培养条件同方法1.3,出苗后每盆定苗30株。日本看麦娘长至3~4叶期后,用10.8%高效氟吡甲禾灵EC以400 ml/hm2进行茎叶喷雾处理,以清水处理为对照,各处理重复4次。以敏感种群受药剂胁迫约7 d后开始出现明显的受害症状为依据,分别于处理后0 d、1 d、2 d、3 d、5 d和7 d采集各处理植株的叶片,取样后立即测定各项生理指标。
1.4.2 生理指标测定 分别测定各样品的叶绿素(Chl.)、类胡萝卜素(Car.)、可溶性糖和丙二醛(MDA)的含量以及过氧化物酶(POD)活性。测定方法参照《植物生理生化实验原理和技术》[12],其中可溶性糖含量用苯酚法测定;POD活性用愈创木酚法测定。试验数据采用Excel 2003处理并作图;用DPS数据处理软件进行单因素方差分析,采用Duncan氏新复极差法进行多重比较,最小显著法(LSD)检验差异显著性。
2 结果
2.1 抗性水平检测结果
抗性水平检测结果(表1)表明,高效氟吡甲禾灵对JR、DY和CZ种群日本看麦娘种子胚芽生长抑制的 EC50分别为5.796 3 μg/ml、0.425 2 μg/ml和0.067 9 μg/ml。以采自安徽滁州未用药的日本看麦娘种群(CZ种群)对高效氟吡甲禾灵的敏感性作为敏感基线得出,JR和DY种群的相对抗性倍数分别为85.36和6.26。即JR种群为高抗生物型,DY种群为低抗生物型。
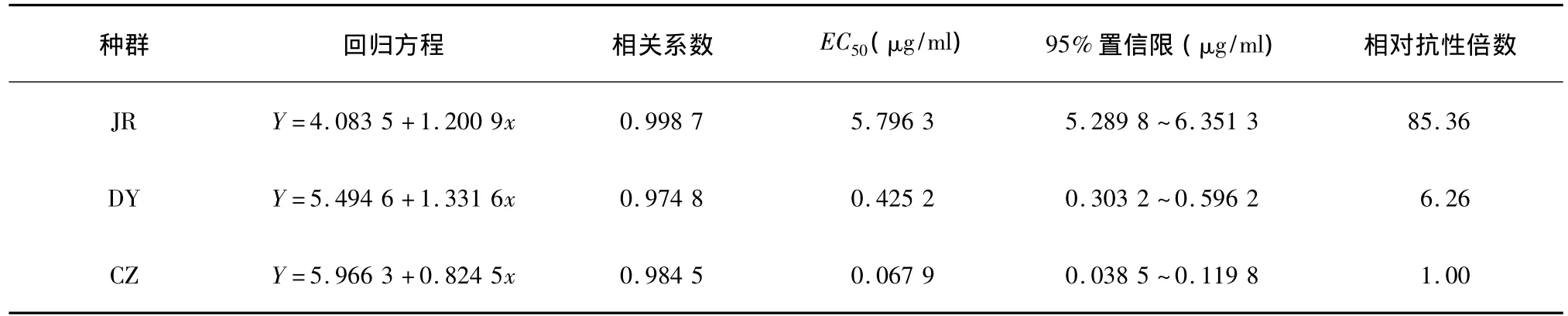
表1 不同日本看麦娘种群对高效氟吡甲禾灵的抗性水平检测Table 1 Resistance levels of different Alopecurus japonicus populations to haloxyfop-R-methyl
2.2 日本看麦娘喷施高效氟吡甲禾灵后叶片光合色素含量
从表2可以看出,JR种群日本看麦娘,在药剂胁迫后的第 0 d、1 d、2 d、3 d、5 d 和 7 d,叶绿素 a(Chl.a)、叶绿素 b(Chl.b)和总叶绿素(Chl.)含量略有下降,而对照均略有上升;类胡萝卜素(Car.)含量和 Chl.a/Chl.b比值与对照一样略有上升;Car./Chl.比值略有上升趋势,对照则基本稳定。DY种群和CZ种群日本看麦娘,在药剂胁迫后,Chl.a、Chl.b、Chl.和 Car.含量均显著下降,而对照略有上升。CZ种群日本看麦娘,在药剂胁迫后第 7 d,Chl.a、Chl.b、Chl.和Car.含量分别为对照的 31.52%、40.83%、34.00%和23.97%,下降最显著。DY种群和CZ种群日本看麦娘,Chl.a/Chl.b比值均有下降,CZ种群下降较明显,而对照略有上升。DY种群日本看麦娘 Car./Chl.比值略有上升,对照基本稳定。CZ种群日本看麦娘 Car./Chl.比值下降显著,对照基本稳定。
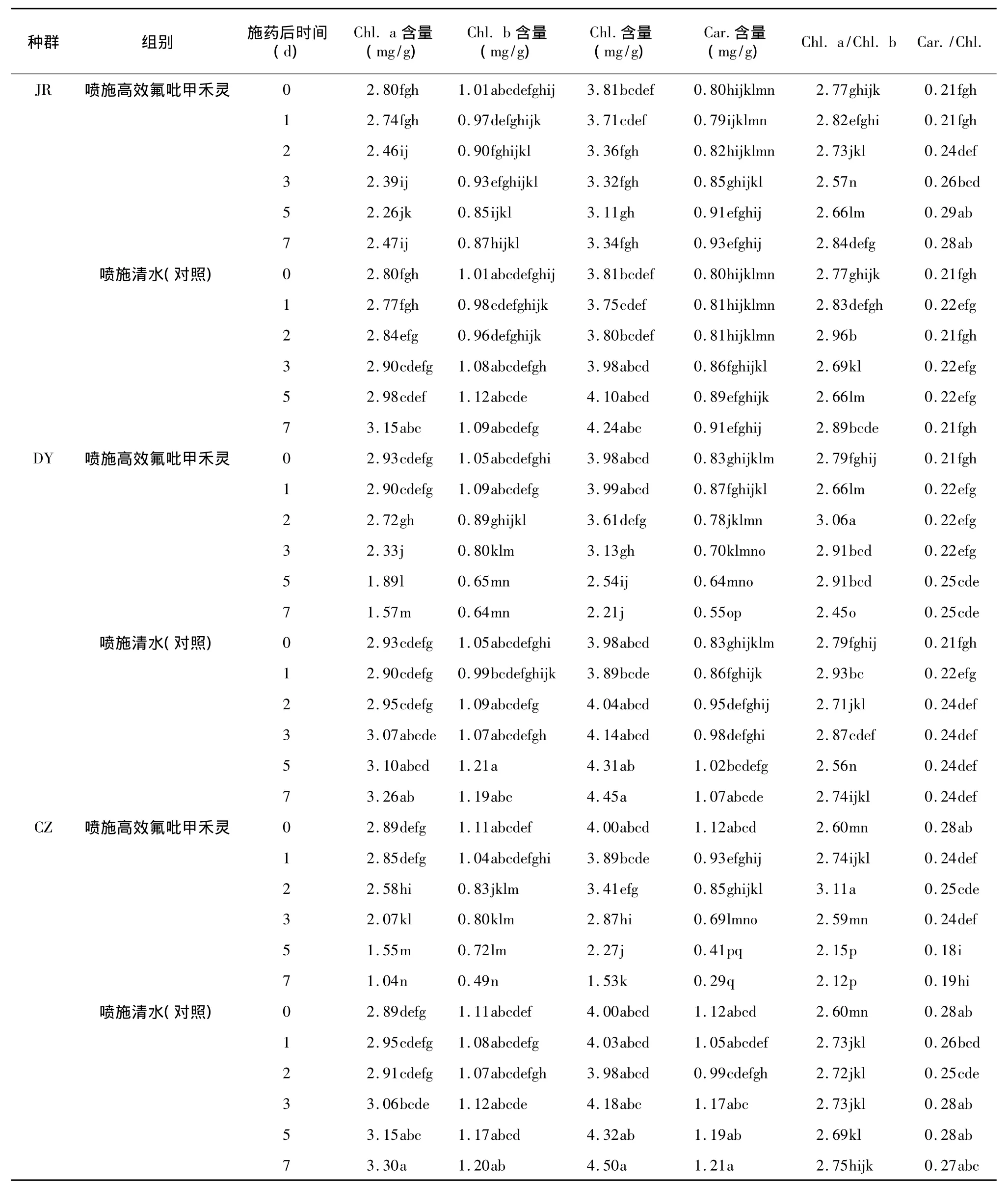
表2 不同抗性生物型日本看麦娘喷施高效氟吡甲禾灵后叶片光合色素的含量Table 2 Photosynthetic pigments contents in the leaves of Alopecurus japonicus with different resistances after haloxyfop-R-methyl spraying
2.3 日本看麦娘喷施高效氟吡甲禾灵后叶片可溶性糖含量
从图1可以看出,不同抗性生物型日本看麦娘在喷施高效氟吡甲禾灵后,叶片可溶性糖含量均有上升趋势,其中高抗生物型略有上升,敏感和低抗生物型上升较为显著。JR种群可溶性糖含量增加相对较少,药剂胁迫后第7 d,可溶性糖含量为其对照的1.5倍;DY种群在药剂胁迫后第7 d,可溶性糖含量为其对照的2.3倍;CZ种群可溶性糖含量增加最显著,药剂胁迫后第7 d,可溶性糖含量为其对照的3倍。
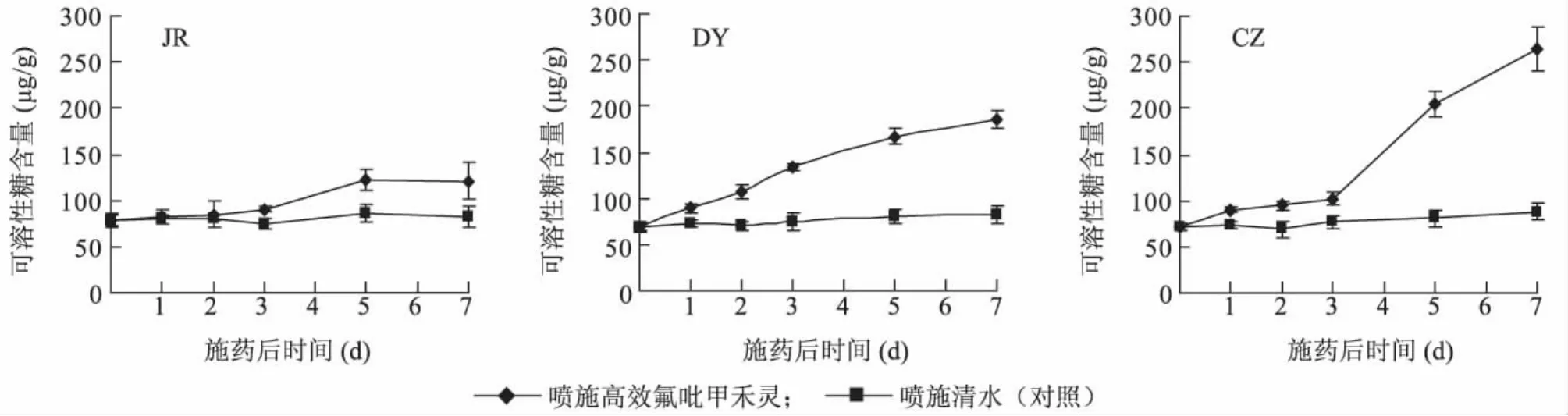
图1 不同抗性生物型日本看麦娘喷施高效氟吡甲禾灵后叶片可溶性糖含量Fig.1 Soluble sugar contents in the leaves of Alopecurus japonicus with different resistances after haloxyfop-R-methy spraying
2.4 日本看麦娘喷施高效氟吡甲禾灵后叶片MDA含量
从图2可以看出,不同抗性生物型日本看麦娘在喷施高效氟吡甲禾灵后,叶片MDA含量变化差异显著。JR种群在药剂胁迫后7 d内,叶片MDA含量与其对照差异不显著。DY种群在药剂胁迫后第3 d,叶片MDA含量开始呈上升趋势,药剂胁迫后第7 d,叶片MDA含量为其对照的2.2倍。CZ种群在药剂胁迫后,叶片MDA含量呈上升趋势,药剂胁迫后第7 d,叶片MDA含量为其对照的4倍。
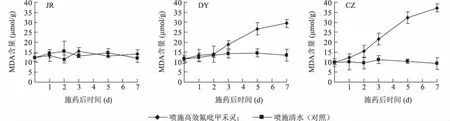
图2 不同抗性生物型日本看麦娘喷施高效氟吡甲禾灵后叶片MDA含量Fig.2 MDA contents in the leaves of Alopecurus japonicus with different resistances after haloxyfop-R-methyl spraying
2.5 日本看麦娘喷施高效氟吡甲禾灵后叶片POD活性
从图3可以看出,不同抗性生物型日本看麦娘在喷施高效氟吡甲禾灵后,叶片POD活性均呈现先上升后下降的趋势,其中高抗生物型变化平缓,而低抗和敏感生物型则反应强烈。JR种群在药剂胁迫后第5 d,叶片POD活性为其对照的1.7倍,第7 d降至其对照的1.4倍。DY种群在药剂胁迫后5 d内,叶片POD活性有明显上升趋势,第5 d时为其对照的3倍,第7 d时叶片又降至对照的1.5倍。CZ种群在药剂胁迫后5 d内,叶片POD活性上升最显著,第5 d时为其对照的3.7倍,第7 d时又降至对照的2.2倍。这表明,高抗生物型日本看麦娘受高效氟吡甲禾灵的影响较小,表现出一定的生理适应性。
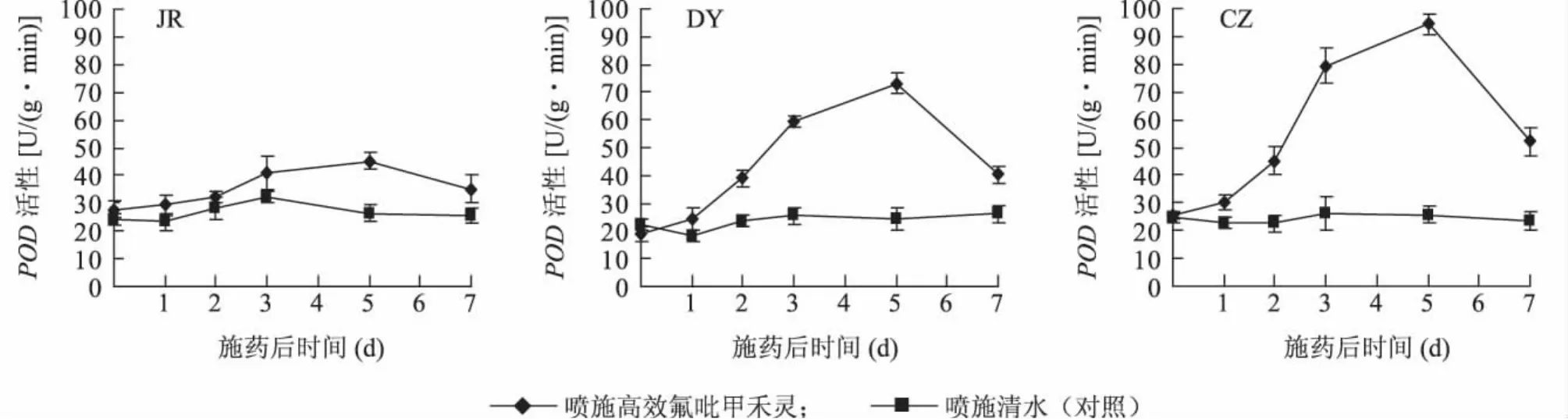
图3 不同抗性生物型日本看麦娘喷施高效氟吡甲禾灵后叶片POD的活性Fig.3 POD activities in the leaves of Alopecurus japonicus with different resistances after haloxyfop-R-methyl spraying
3 讨论
由于过度依赖和长期使用相对有限的化学除草剂,导致了杂草抗药性的发生和发展,且杂草抗药性问题越来越突出,备受全球关注。抗药性检测是抗性杂草治理的重要环节,其研究方法不断发展并日渐重要[13]。目前,杂草抗药性检测的方法主要有种子生物测定法、整株测定法、幼苗实验法、分蘖法、花粉粒检测法、酶联免疫测定(ELISA)法、PCR 检测法等[14-16]。种子生物测定法,只需观察种子发芽趋势就可准确判断是否发生抗药性,具有快速、简便易行、设备简单、成本低、占用空间小、适用于大面积种子的检测等优点,因而在抗药性检测中被广泛应用。本研究利用种子生物测定法检测了不同油菜田日本看麦娘种群对高效氟吡甲禾灵的抗药性,结果表明采自江苏省句容市的JR种群日本看麦娘为高抗生物型,这与前人的研究结果一致[2-3],而采自江苏省丹阳市的 DY种群为低抗生物型。
在喷施高效氟吡甲禾灵后,低抗和敏感生物型油菜田日本看麦娘叶片的 Chl.a、Chl.b、Chl.和 Car.含量有明显下降,光合色素构成发生变化,叶片可溶性糖和MDA含量上升显著,POD活性先上升后下降并且反应强烈。这表明,低抗和敏感生物型日本看麦娘植株体内的保护酶系统受到损害,清除活性氧及过氧化物自由基,能力大幅下降,细胞质膜受损严重,其中敏感生物型比低抗生物型受药剂的影响更为明显。高抗生物型日本看麦娘,通过增加渗透调节剂可溶性糖含量,来保护原生质与环境之间的渗透平衡和结构的完整性,增加POD活性来清除活性氧及过氧化物自由基,从而降低了药剂胁迫下叶片膜脂过氧化产物MDA的生成量。这表明,高抗生物型日本看麦娘对高效氟吡甲禾灵具有一定的生理适应性。
致谢: 感谢南京农业大学植物保护学院董立尧教授为本研究提供了油菜田野生日本看麦娘种子!
[1]强 胜.杂草学[M].北京:中国农业出版社,2001.
[2]杨彩宏,董立尧,李 俊,等.油菜田日本看麦娘对高效氟吡甲禾灵抗药性的研究[J].中国农业科学,2007,40(12):2759-2765.
[3]潘以楼,朱桂梅,杨敬辉.日本看麦娘对芳基苯氧基丙酸类除草剂的耐性[J].杂草科学,2007(3):19-21.
[4]陈宏州,朱桂梅,杨敬辉,等.烯草酮与高效氟吡甲禾灵对日本看麦娘的毒力及田间防效[J].杂草科学,2009(3):25-27.
[5]MARLES M A,DEVINE M D,HALL J C.Herbicide resistance in Setaria viridis conferred by a less sensitive form of acetyl-CoA carboxylase[J].Pesticide Biochemistry and Physiology,1993,46(1):7-14.
[6]DEVINE M D.Mechanisms of resistance to acetyl-coenzyme A carboxylase inhibitors:a review[J].Pesticide Science,1997,51(3):259-264.
[7]PRESTON C,TARDIF F J,CHRISTOPHER J T,et a1.Multiple resistance to dissimilar herbicide chemistries in a biotype of Lolium rigidum due to enhanced activity of several herbicide degrading enzymes[J].Pesticide Biochemistry and Physiology,1996,54(2):123-134.
[8]黄世霞,何金铃,王庆亚,等.看麦娘对稀禾啶和高效氟吡甲禾灵产生抗药性的分子基础[J].激光生物学报,2010(6):832-837.
[9]韩瑞娟,董立尧,李 俊,等.日本看麦娘对高效氟吡甲禾灵代谢抗性的初步研究[J].杂草科学,2010(1):3-7.
[10]陈宏州,杨敬辉,朱桂梅,等.油菜田日本看麦娘对高效氟吡甲禾灵的抗药性检测[J].西南农业学报,2012,25(2):502-506.
[11]TAL A,KOTOUIL-SYKA E,RUBIN B.Seed-bioassay to detect grass weeds resistant to acetyl coenzyme acarboxylase inhibiting herbicides[J].Crop Protection,2000,19:467-472.
[12]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[13]张朝贤,倪汉文,魏守辉,等.杂草抗药性研究进展[J].中国农业科学,2009,42(4):1274-1289.
[14]LETOUZE A,GASQUEZ J.A rapid reliable test for screening aryloxyphenoxypropionic acid resistance within Alopecurus myosuroides and Lolium spp.populations[J].Weed Research,1999,39(1):37-48.
[15]LETOUZE A,GASQUEZ J.A pollen test to detect ACCase targetsite resistance within Alopecurus myosuroides populations[J].Weed Research,2000,40(2):151-162.
[16]LETOUZE A,GASQUEZ J.Inheritance of fenoxaprop-P-ethyl resistance in a blackgrass(Alopecurus myosuroides Huds.)population[J].Theoretical& Applied Genetics,2001,103(2/3):288-296.
