阿拉伯糖异构酶的重组表达及性质研究
2013-07-22王玥刘秀梅魏晓琨朱玲
王玥,刘秀梅,魏晓琨,朱玲
(1.天津科技大学生物工程学院工业发酵微生物教育部重点实验室,天津 300457;2.上海斯贝生物科技有限公司,上海 200235)
D-塔格糖(D-tagatose)是一种低热量与蔗糖甜度相近的食品甜味剂[1-2]。对其他甜味剂也有较好的协同增效作用[3]。D-塔格糖的生理功效体现在控制高血糖和糖尿病并发症[4-5]、平衡肠道菌群[6]、预防龋齿[7]等方面。大量毒理试验证明塔格糖安全无毒,国际上很多国家包括美国,新西兰和澳大利亚以及中国,已批准其在食品中广泛应用[8-9]。
截止到21 世纪初,D-塔格糖的制备主要有化学和生物两种转化方法。化学转化的问题是产物复杂难于与目标产物分离[10];生物转化法包括发酵法和酶法[11]。发酵法周期长产量低,不适合工业扩大生产。酶法制备则是以D-半乳糖为原料,在L-阿拉伯糖异构酶催化下进行转化[12]。L-阿拉伯糖异构酶由araA 基因编码,参与L-阿拉伯糖的代谢。由于D-半乳糖和L-阿拉伯糖在三维结构上相似,见图1,因此L-阿拉伯糖异构酶还能催化D-半乳糖生成D-塔格糖[13]。该酶在用于生产D-塔格糖中具有很大商业价值[14]。

图1 L-阿拉伯糖异构酶催化的反应Fig.1 In vivo and in vitro reactions catalyzed by AI
本文采用DNA 重组技术构建了一株高表达L-阿拉伯糖异构酶的基因工程菌,将E.coli K12 基因组中的araA 基因插入载体pET-32a,并转入宿主菌BL21(DE3)中高效表达,随后对粗酶的酶活和最适酶活条件进行测定。
1 材料与方法
1.1 材料
1.1.1 菌种与质粒
大肠杆菌K12(Escherichia coli)、大肠杆菌BL21(DE3)、载体pET-38a 为上海斯贝生物科技有限公司实验室保存;pMD-18T 载体购自上海皓嘉科技发展有限公司。
1.1.2 主要试剂
限制性内切酶、T4 DNA 连接酶购自上海皓嘉科技发展有限公司;DNA 回收试剂盒购自上海捷瑞生物工程有限公司并由该公司完成PCR 引物合成;2xTaq PCR MasterMix 购自天根生化科技有限公司;测序由上海英骏生物技术有限公司进行。其它试剂均为市售分析纯。
1.1.3 培养基
LB 培养基:胰蛋白胨10 g/L,酵母粉5 g/L,NaCl 10 g/L,pH 7.2;固体培养基中添加1.5%的琼脂;固体、液体LB 培养基使用时按需要添加氨苄青霉素(Amp,终浓度100 μg/mL)。
1.2 方法
1.2.1 araA 基因的PCR 扩增
根据GenBank 中大肠杆菌L-阿拉伯糖异构酶(ECAI)基因序列设计PCR 引物,分别为araA(+):GCTCTAGAAATAATTTTGTTTAACTT TAAGAAGGAGA TATACCATGACGATTTTTG,Xba I,rbs(点划线);araA(-):GGAAGCTTTTAGCGACGAA ACCCGTAATA CAC TTCGTTC,Hind Ⅲ。
由于选用的酶切位点为Xba I 和Hind III,质粒上的RNA 聚合酶结合位点被切除,故须在引物araA(+)处补入,,确保基因的正常转录。PCR 扩增的模板为大肠杆菌K12 基因组DNA,于94 ℃预变性5 min,变性30 s,65 ℃退火30 s,72 ℃延伸90 s,循环30 次,最后72 ℃延伸5 min。
1.2.2 表达载体的构建以及目的基因的诱导表达
将回收的PCR 产物连接到pMD-18T 载体上并转化到E.coli DH5α,筛选氨苄抗性的阳性克隆,经测序正确,将目标片段克隆到pET-32a 上,并转化入E.coli BL21(DE3)中,筛选氨苄抗性的阳性克隆。将该阳性克隆菌株于37 ℃条件下置于50 μg/mL 氨苄霉素LB 培养基中培养,测得OD600达到0.4~0.6 时加入IPTG 至0.5 mmol/L,15 ℃诱导10 h,4 000 r/min 离心20 min,收集菌体,用50 mmol/L Tris-HC(lpH7.6)重悬菌体。诱导产物用12%的SDS-PAGE 分析。
1.2.3 L-阿拉伯糖异构酶的酶活测定
以重悬菌体为粗酶源、D-半乳糖作为酶活测定的底物,3 mL 酶活测定体系包括0.6 mL 菌悬液,0.4 g/L D -半乳糖,1 mmol/L MnCl2,50 mmol/L Tris -HCl(pH7.6)。60 ℃反应1 h,每隔15 min 取样0.5 mL 到0.5 mL 0.1 mol/L HCl 中,测定D-塔格糖含量。D-塔格糖含量的测定采用半胱氨酸-咔唑法[15]。在上述测定条件下,将每小时产生1 μg D-塔格糖所需的酶量定义为一个酶活单位(U)。
1.2.4 酶活最适条件的测定
根据上述酶活测定方法,分别从温度、pH、二价金属离子以及Mn2+浓度四个方面来研究酶活测定的最适条件。
温度:分别在30、37、40、50、60、70 ℃下测定酶活,其中反应体系pH 为7.6,离子为Mn2+。
pH:采用磷酸氢二钠-磷酸二氢钾缓冲液(pH 4.92~8.18)和Tris-HCl 缓冲液(pH 7.10~9.00),在pH 6、pH 6.6、pH 7、pH 7.6、pH 8、pH 8.6、pH 9 条件下进行酶活测定。反应温度为60 ℃,离子为Mn2+。
二价金属离子:在Mn2+、Co2+、Mg2+、Ca2+、Fe2+、Ni2+、Zn2+、Cu2+存在下测定酶活,各离子浓度均为1 mmol/L,以不加入任何二价金属离子所测定的酶活为对照。
Mn2+浓度:向重悬菌体中加入10 mmol/L EDTA 以去除金属离子,37℃处理2h。离心去上清,再用50mmol/L Tris-HCl(pH7.6)重悬。酶活测定中分别加入0、1、3、5、7、9 mmol/L Mn2+来检测Mn2+浓度对酶活的影响。以未经EDTA 处理的酶的酶活为参照。
2 结果与讨论
2.1 重组质粒pET-38a-araA 的构建与鉴定
以大肠杆菌K12 基因组模板,经PCR 扩增araA得到1 500 bp 左右的特异性条带,其大小与理论预期一致。回收目标条带,直接连接到pMD-T 载体上,转入到大肠杆菌DH5α,筛选氨苄抗性的阳性克隆,并测序。测序结果与GenBank 上公布的大肠杆菌K12 的araA 基因(EG10052)相比,第213 位的由T 变为C,第1395 位由C 变化T。但是对照密码子表,两者都是第三位发生突变,前者由GAT 变成GAC,但其对应的氨基酸未变,仍然为天冬氨酸;后者由TTC 变成TTT,对应的氨基酸仍然为苯丙氨酸。因此,基因序列的突变并不影响目的蛋白的序列。
将T 载体双酶切得到1 500 bp 左右的条带克隆到质粒pET-32a 上,获得重组质粒pET-32a-araA,转入到大肠杆菌BL21(DE3)中,筛选氨苄抗性的阳性克隆菌落,获得重组araA 的工程菌AraA。
2.2 L-阿拉伯糖异构酶的诱导表达
将上述构建的工程菌AraA,按材料与方法中所述进行培养及诱导,所得菌体经超声波破碎,并经SDSPAGE 电泳,结果如图2 所示。

图2 重组菌AraA SDS-PAGE 蛋白电泳图Fig.2 SDS-PAGE result of expression product of araA
经IPTG 诱导后,L-阿拉伯糖异构酶在宿主菌E.coli BL21(DE3)中有显著的表达,见图2 的Line2、3,其大小约为56 kDa,与文献报道的L-AI 的单体分子量(Mr=56 074 Da)大小一致。目的蛋白大部分为胞内表达,离心沉淀中仍有少量包涵体存在,见图2 的Line7。
对相同浓度的重悬菌体、重悬菌体破碎液、破碎上清和破碎沉淀进行酶活测定,结果见图3。

图3 不同类型酶源的活力比较Fig.3 The activity construction of suspension of AI
通过比较,重悬菌体的酶活最高,因此酶活测定时均采用重悬菌体作酶源。与重悬菌体相比,由于超声破碎对酶活产生影响,导致菌体破碎液的酶活有一定下降。破碎液中的酶活完全来自于上清液中的可溶蛋白,这说明包涵体中的蛋白没有活性。
2.3 最适酶活条件
2.3.1 温度
按照酶活测定方法,在不同的温度下考察重组菌的酶的活力,确定最适酶活温度,结果见图4。

图4 温度对酶活测定的影响Fig.4 Effect of temperature on the activity of AI
从图4 中可以看出,随着温度的增加,酶活不断增大,在60 ℃时达到最大活力,最高酶活为746.8 U/mL,并设为100%作为对照。虽然大肠杆菌的最适生长温度为37 ℃,可此时的酶活却仅为最高酶活的43%。明显不同于文献[12-15]中所述的37 ℃为最适酶活温度,即常温菌中AI 的最适酶活温度能够达到嗜热菌中AI 的最适酶活温度。这是前所未有的。
2.3.2 pH
采用磷酸氢二钠-磷酸二氢钾缓冲液(pH 6、pH 6.6、pH 7)和Tris-HCl 缓冲液(pH 7.6、pH 8、pH 8.6、pH 9),在不同pH 条件下对L-AI 进行酶活测定,结果见图5。

图5 pH 对酶活测定的影响Fig.5 Effect of pH on the activity of AI
从图5 中可以看出,在酸性条件下,L-AI 具有极低的活力,因此L-AI 不适合在酸性条件下进行反应。在碱性条件下,随着pH 的逐渐增加,L-AI 活力逐渐下降。在pH7.6 时达到最高活力,最高活力为619.6 U/mL。
2.3.3 二价金属离子
在Mn2+、Co2+、Mg2+、Ca2+、Fe2+、Ni2+、Zn2+、Cu2+存在下测定酶活,各种离子浓度均为1 mmol/L,以不加入任何金属离子所测定的酶活为对照,结果见图6。
从图6 中可以看出,测定中添加Mn2+、Co2+和Fe2+能明显地提高酶活,且Mn2+最高,相对活力高达190%。Mg2+、Ca2+和Ni2+对提高酶活的影响不太明显,而Zn2+和Cu2+却抑制了酶活测定。其中不加入任何金属离子时的酶活为304 U/mL,并设为100%作为对照。
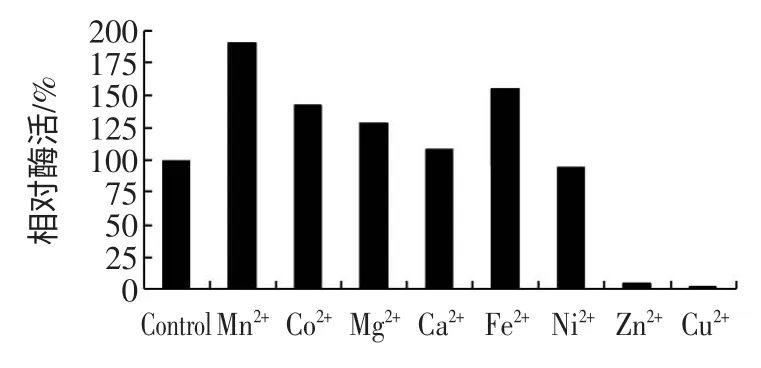
图6 金属离子对酶活测定的影响Fig.6 Effect of divalent metallic ion on the activity of AI
2.3.4 Mn2+浓度
以EDTA 处理后的重悬菌体为酶源,分别加入0、1、3、5、7、9 mmol/L Mn2+来检测Mn2+浓度对酶活的影响,结果见图7。以未经EDTA 处理的酶的酶活为参照(100%)。

图7 Mn2+浓度对酶活测定的影响Fig.7 Effect of concentration of Mn2+on the activity of AI
从图7 可以看出,经过EDTA 处理后,酶完全没有活性。加入Mn2+后,可以使EDTA 处理过的酶重新恢复活力。但活力恢复后,酶活并不随着Mn2+浓度的增加而增加。这表明L-阿拉伯糖异构酶需要Mn2+催化,而且加入Mn2+有助于提高酶活,但提高酶活的量是有限度的。其中未经EDTA 处理的酶液的酶活为660 U/mL,并设为100%作为对照,活力恢复后的酶活均能达到590 U/mL 以上。
3 讨论
本研究中选择E.coli K12 基因组作为模板,用PCR 方法克隆得到L-阿拉伯糖异构酶基因araA,并将其转入表达载体pET-32a 中。基于表达载体上T7 lac 的强启动表达功能,使得目的蛋白在宿主菌BL21(DE3)中得到很高的表达量。经SDS-PAGE 分析,15 ℃诱导10 h 后,目的蛋白主要以可溶性蛋白形式存在于菌体中,而无活性的包涵体表达量极低。以重悬菌液为酶源、D-半乳糖为底物,测定了L-阿拉伯糖异构酶的最适酶活条件。其结果表明:酶的最适温度为60 ℃,最适pH 为7.6,酶活测定中加入Mn2+能显著提高酶活,1 mmol/L Mn2+能使酶活相对提高50%。
[1]Pil Kim.Current studies on biological tagatose production using Larabinose isomerase:a review and future perspective[J].Applied microbiology and biotechnology,2004,65(3):243-249
[2]Levin GV.Tagatose,the new GRAS sweetener and health product[J].Journal of medicinal food,2002,5(1):23-36
[3]Bertorelli AM,Czarnowski-Hill JV.Review of present and future use of nonnutritive sweeteners[J].Diabetes Educ,1990,16(5):415-422
[4]梁敏,翟娅菲,邹洋,等.新型甜味剂塔格糖的应用及生产[J].食品与药品,2011(3):125-128
[5]Ercan-Fang N,Gannon M C,Rath V L.Integrated effects of multiple modulators on human liver glycogen phosphorylase a[J].Am physiol endocrinol metab,2002,283(1):29-37
[6]Bertelsen H,Jesen BB,Buemann B.D-tagatose:a novel lowcalorie bulk sweetener with prebiotic properties[J].World review of nutrition and dietetics,1999,85:98-109
[7]Claire LK,Margaret HW,Vasilios HF.90-day oral toxicity study of D-tagatoseinrats[J].Regulatorytoxicologyandpharmacology,1999,29:1-10
[8]Burdock GA,Garabin I G.Generally recognized as safe (GRAS):history and description[J].Toxico Lett,2004,150(1):3-18
[9]Benjamin B,Soren T,Arne A.Human gastrointestinal tolerance to Dtagatose[J].Regulatory toxicology and pharmacology,1999,29(2):71-77
[10]黄闻霞,江波,沐万孟,等.化学法合成D-塔格糖的研究[J],食品工业科技,2008,1(1):247-249
[11]Oh DK.Tagatose:properties,applications,,and biotechnological processes[J].Appl microbiol biotechnol,2007,76(1):1-8
[12]Granström TB,Takata G,Tokuda M,et al.Izumoring:a novel and complete strategy for bioproduction of rare sugars[J].J Biosci Bioeng,2004,97(2):89-94
[13]Lee SJ,Lee DW,Choe EA,et al.Characterization of a Thermoacidophilic L-Arabinose Isomerase from Alicyclobacillus acidocaldarius:Role of Lys-269 in pH Optimum[J].Appl Envir Microbiol,2005,71(12):7888-7896
[14]Levin GV.Tagatose,The new GRAS sweetener and health product[J].Med Food,2002,5(1):23-36
[15]Dische Z,Borenfreund E.A new spectrophotometric method for the detection and determination of keto sugar and trioses[J].Journal of biological chemistry,1951,192(2):583-587
