三种豆科植物光合特征初步研究
2013-02-24瞿思民吴夏华瞿巧文尹瑞安
瞿思民,吴夏华,瞿巧文,尹瑞安
(渐江省庆元县林业局,浙江庆元 323800)
山核桃(CaryacathayensisSarg.为我国特有的高档干果和木本油料植物,20世纪80年代随着人们生活水平的提高及对保健休闲产品需求的增加,农民种植山核桃积极性高涨,每年新增面积约3万亩。山核桃栽培中,农民为采收方便,大量使用除草剂去除林地杂草,从而造成山核桃林生物多样性下降、病虫害加剧、水土流失等生态问题。
随着人们生态意识的增强和山核桃产业的的可持续发展,如何在保持山核桃经济效益的基础上,实现生态效益和社会效益的最大化,是摆在当地政府和林农面前的一个重大问题。林草复合栽培是农业结构调整、改变单一种植结构、提高单位面积产出量及生态经济效益的有效途径[7-9]。近几年山核桃产区林业技术部门对林下生草栽培进行了一系列的探索,如不同生草的选择、栽培模式及技术等,选择的草种包括白三叶(TrifoliumrepensLinn.)、紫云英(AstragalussinicusL.)和紫花苜蓿(MedicagosativaL.),黑麦草(Loliumperenne)、油菜(Brassica chinensisL.)等。本文对栽植于山核桃林下的三种豆科植物的光合生理进行初步研究,以期为山核桃林下生草栽培提供理论基础。
1 材料与方法
1.1 研究地概况
实验地设在浙江省临安市浙江农林大学东湖校区苗圃,地理位置为E 118°51′,N 29°56′,该区属亚热带季风气候,年平均气温16℃,极端最高气温41.7℃,极端最低气温-13.3℃,年有效积温5774℃,年降水量1350~1500 mm,年日照时数1774 h,无霜期235 d。海拔高度在90 m,土壤为发育于凝灰岩的红壤土类。
1.2 试验材料
3种豆科植物分别是白三叶、紫云英和紫花苜蓿,均为种子繁殖,于2010年9月种植于山核桃林下,常规管理。
1.3 气体交换参数测定
于2011年5月25日上午9:30~11:00时,对3种豆科植物进行光合特性测定。测为生长完好、无病虫害、叶面积相近的完全展开叶。光响应曲线利用Li-6400(美国,LI-Cor公司)红蓝光源6400-02测定,测量时的叶室温度设置为25℃,CO2浓度370 μmol/L,光照强度梯度设置1500、1000、800、500、300、100、50、0μmol/m2·s,测定的的指标有光合速率(Pn 蒸腾速率(Tr 胞间CO2浓度(Gs 气孔导度(Gs)等。每种植物测定6片叶片。
1.4 数据分析
采用Excel、DPS等数据分析软件进行数据分析。
2 结果与分析
2.1 三种豆科植物光合速率的差异
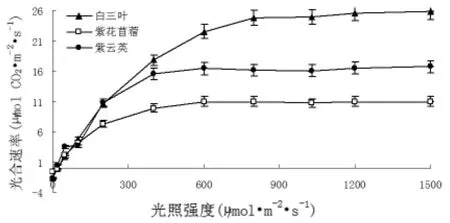
图1 不同豆科植物的光饱和曲线
光响应曲线反映了植物光合速率随光照强度的变化而变化的规律[8]。从图l可以看出,随着光量子通量的增大,净光合速率几乎呈直线上升;当光量通量达到一定值后,净光合速率增加的幅度就逐渐减慢,最后达到一定限度,不再随光量子通量的增加而增加,即达到光饱和。图1表明,在相同条件下,白三叶的光合速率最大。通过光响应曲线可以计算出光补偿点、光饱和点及最大净光合速率,计算结果见表1。由于植物在光补偿点时不能积累干物质,因此,光补偿点的高低可以作为判断植物在低光照强度条件下能否生长的标志。光补偿点越小表明植物利用弱光的能力越强,紫云英、紫花苜蓿、白三叶光补偿点分别为12.20、15.14、24.16μmol/m2·s。植物在光饱和点时光合速率最大,因此,光饱和点反映了植物利用强光的能力,越高说明植物在受到强光时不易发生抑制,植物的耐阳性越强,紫云英、紫花苜蓿、白三叶树光饱和点分别为510.67、512.35、758.00μmol/m2·s,最大净光合速率为15.55、10.70、23.58μmol CO2/m2·s。
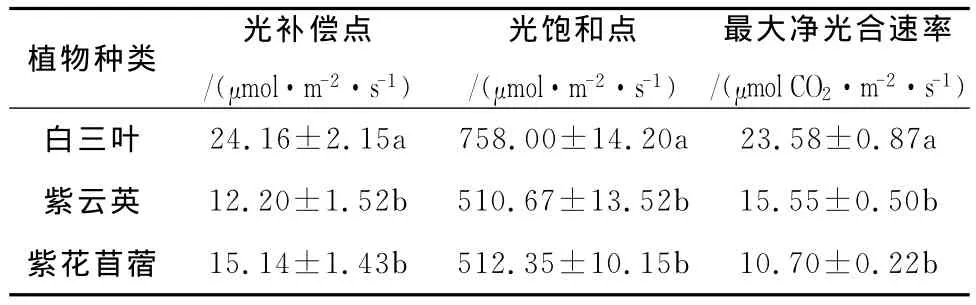
表1 三种豆科植物光补偿点、光饱和点和光饱和点时净光合速率
2.2 三种豆科植物气孔导度的差异
从图2中可知,在光合过程初期,气孔导度随光强的增大而增大,光强为200μmol/m2·s时,三种植物的气孔导度均达最大值,随着光强的继续增大,气孔导度则逐渐减小。从图中还可看出。在整个光合过程中白三叶的气孔导度明显大于紫云英和紫花苜蓿。
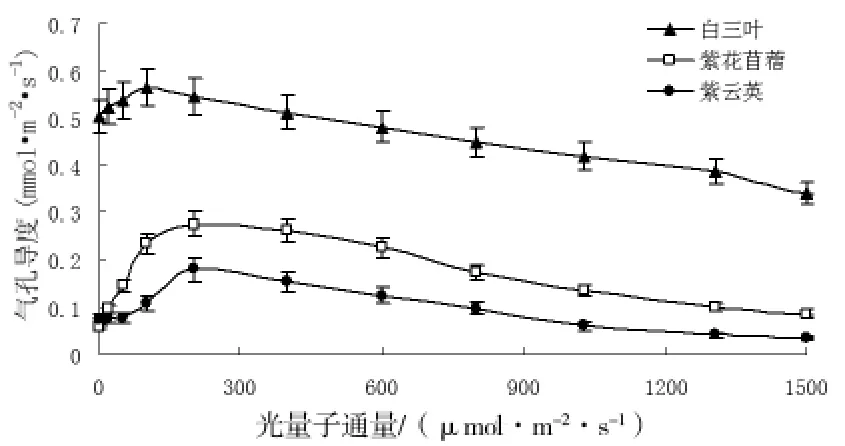
图2 不同豆科植物的气孔导度对光强的响应
2.3 三种豆科植物胞间二氧化碳浓度的差异
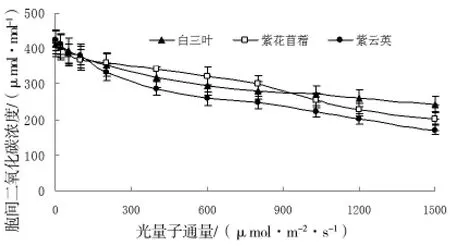
图3 胞间二氧化碳浓度对光强的响应
图3结果显示,三种植物胞间二氧化碳浓度均着随光强的增大而减小,植物间的差异并不明显。
2.4 三种豆科植物蒸腾速率的差异
三种豆科植物蒸腾速率的变化见图4。从图中可知,光量子通量密度为0~50μmol/m2·s时,白三叶、紫云英、紫花苜蓿的蒸腾速率均随光强的增大而增大,而后分别保持5.4、1.5、2.4mmol/m2·s左右。在整个光合过程中,白三叶的蒸腾速率明显大于其它两种植物。
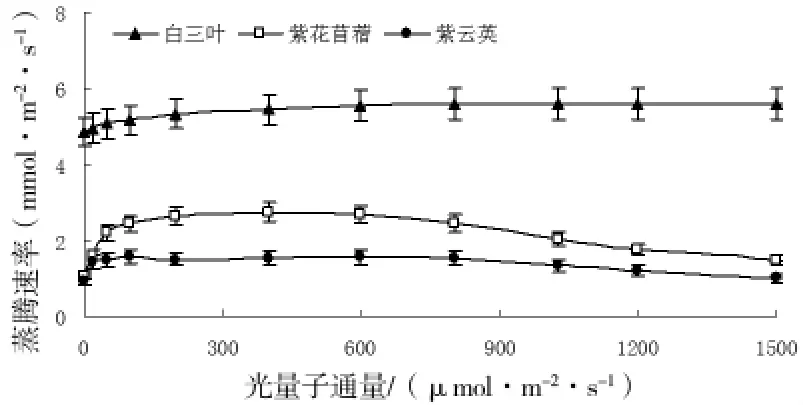
图4 蒸腾速率对光强的响应
3 结论
通过对三种豆科植物气体交换参数的测定,结果表明,白三叶、紫云英、紫花苜蓿的光补偿分别为24.16、12.20、15.14μmol/m2·s,光饱和点分别为758.00,510.67,512.35μmol/m2·s,最大净光合速率为23.58 15.55 10.70μmol CO2m s在整个光合过程中白三叶的气孔导度、腾速率明显大于紫云英和紫花苜蓿,而胞间二氧化碳浓度在植物间的差异并不明显。白三叶光合能力明显高于其它两种植物,仅从光合特征进行分析,山核桃林下以栽培白三叶为好,从而可以产生较大的生物量。
[1] 王冀平,李亚南,马建伟.山核桃仁中主要营养成分的研究[J].食品科学,1998,194:44-46.
[2] 王延平,张海,刘生禹,等.黄土区杏—苜蓿复合系统的土壤水分环境及效益研究[J].中国水土保持科学,2004,2(1):74-78.
[3] 王齐瑞,樊巍,谭晓风.杏—紫花苜蓿生草栽培系统根系分布及生长动态[J].林业科学,2008,44(8):141-144.
[4] 杨英,宋继学,杨继民,等.论苜蓿在农牧业和秀美山川中的作用[J].西安联合大学学报,2001,4(13):97-100.
[5] 樊巍,孔令省,阴三军,等.干旱丘陵区苹果—紫花苜蓿复合系统对苹果生长、产量和品质的影响[J].河南农业大学学报,2004,38(4):423-426,440.
[6] 刘芳,田春丽,刘世亮,等.有机酸—磷配施对紫花苜蓿生长及磷吸收的影响[J].河南农业大学学报,2007,41(4):382-386.
[7] 李文华,赖世登.中国农林复合经营[M].北京:科学出版社,1994.
[8] 陈玉香,周道玮,张玉芬.玉米、苜蓿间作的产草量及光合作用[J].草地学报,2004,12(2):107-112.
[9] 张久海,安树青,李国旗,等.林牧复合生态系统研究评述[J].中国草地,1999(4):52-60.
[10] 王建丽,张永亮.杂花苜蓿光合生理生态特性[J].草地学报,2006,14(2):1382142.
[11] 万素梅,胡建宏,胡守林,等.不同紫花苜蓿品种水分利用效率研究[J].西北农业学报,2004,20(4):133-137.
[12] Vaughn D L,Viands D R.Nut ritive value and forage yield of alfalfa synthetics under three harvestmanagement systems[J].Crop Sci.1990,30:69 9-703.
[13] Olssont,Leverenz JW.Nonuniform stomatal closure and the apparent convexity of the photosynthetic photon flux density response curve[J].Plant,Cell and Environment,1994,17:701-710.
[14] Bhagsari A S,Brown R H.Leaf Photosynthesis and It s Correlation with Leaf Area[J].Crop Sci.1986,26:1272132.
