B型烟粉虱羧酸酯酶对6种寄主植物时间响应的初步研究
2012-11-22柴芳华韩诗畴安新城
柴芳华,韩诗畴,安新城*
(1.中国科学院华南植物园,广州510650;2.广东省昆虫研究所,广州510260;3.中国科学院研究生院,北京100049)
烟粉虱Bemisia tabaci是粉虱科Aleyrodidae小粉虱属Bemisia的一种植食性昆虫,原产于热带亚热带地区,最早由Gennadius在希腊的烟草上发现(Gennadius,1889)。它是一种极端广食性的昆虫,寄主范围多达900余种 (Brown et al.,1995),被世界公认为“超级害虫”。它不仅能够通过刺吸植物的汁液,导致植物衰弱和干枯,分泌蜜露诱发煤污病,影响植物光合作用,烟粉虱还是许多病毒的媒介,传毒种类多达40余种 (Oliveira et al.,2001)。烟粉虱有着异乎寻常的寄主适应能力,这是它寄主谱广的重要原因。每年由烟粉虱对全球蔬菜、瓜果、花卉以及经济作物造成的损失高达数十亿美元 (Byrne and Bellows,1991;Secker et al.,1998)。
植食性昆虫与寄主植物通过长期的协同进化,形成了相互选择、相互作用、相互适应的关系。寄主植物通过分泌有毒次生物质以及调节自身的营养结构等措施来抵御昆虫的取食;而昆虫则通过调节自己的行为机制、生理机制、分子生态机制等策略来适应不同的寄主植物 (Wan et al.,2009),生理机制是烟粉虱最主要的寄主适应机制之一。有研究指出,烟粉虱体内解毒酶的活性可被寄主植物的次生物质诱导增加,从而增强烟粉虱的寄主适应性以及抗药性。烟粉虱在解毒代谢过程中,多功能氧化酶、谷胱甘肽S转移酶、非特异性酯酶、细胞色素P450等组成了昆虫主要的外源化合物防御体系,对于进入昆虫体内的植物毒素或合成农药进行有效地降解、阻隔或异化,使其转变为无毒或低毒的化合物,从而保障昆虫体内代谢环境的稳定性 (Li et al.,2007;Lindroth,1989)。研究表明,羧酸酯酶可能是烟粉虱化学防御体系中最为重要的一类解毒酶系,在寄主适应性、农药抗性、激素调节、发育代谢等过程中广泛地发挥着作用 (Gao,1992;Byrne et al.,1993;Byrne et al.,2000)。本实验通过研究B型烟粉虱在不同寄主植物上繁殖多代后,其体内羧酸酯酶的时间变化规律,旨在揭示烟粉虱适应寄主的生理机制,从而为更深层次的研究烟粉虱的寄主适应机制、抗药性机制、种群爆发机制以及防治策略提供理论依据。
1 材料与方法
1.1 材料
B型烟粉虱:供试烟粉虱来源于广东省昆虫研究所养虫室内黄瓜Cucumis sativus寄主上繁殖的实验种群,分别转移至苦瓜Momondica charantia、木瓜 Carica papaya、水瓜 Luffa acutangula、薄荷Mentha haplocalyx、金银花Lonicera Japonic上饲养超过40天的B型烟粉虱。
植物材料:在自然条件下生长的黄瓜、苦瓜、水瓜、木瓜、薄荷、金银花。
1.2 试剂与仪器
α-乙酸萘酯 (α-Naphthylacetate)、牛血清蛋白 (AlbuMin bovine V)(BSA)、α-萘酚 (1-Naphthol)、毒扁豆碱 (Eserine)、考马斯亮蓝G-250(Coomassie Brilliant Blue G250)、固蓝 B盐(Fsat blue Bsalt)、十二烷基硫酸钠 (sodium dodecyl sulfate,SDS),均购自Sigma公司。低温高速离心机 (型号3K15),美国sigma公司产品;VICTORTM3多标记微孔板检测仪,美国-PerkinElmer公司产品。
1.3 烟粉虱寄主转移的处理
让烟粉虱在自然生长的黄瓜样地里繁殖多代后,分别放几盆苦瓜、水瓜、木瓜、薄荷、金银花,让黄瓜寄主上的烟粉虱自由扩散到这些非嗜好性寄主植物上,再把这些植物移出黄瓜样地,放置在自然条件下单独种植。烟粉虱在其上自由繁殖40 d左右后,便开始分时间段取各种植物上的烟粉虱进行羧酸酯酶活性的测定。由于每种植物的生长周期有别,导致寄主植物上的落虫量有差别,各种寄主植物上烟粉虱的采集时间存在一定差异。
1.4 羧酸酯酶活性的测定方法
羧酸酯酶活性测定的原理:用α-萘酚制作标准曲线。底物α-乙酸萘酯在羧酸酯酶 (CarE)的作用下发生水解反应,生成的α-萘酚与显色剂固蓝B盐生成深蓝色或深红色物质,在595 nm处比色测定其吸光度 (OD值)。
CarE酶源制备:取大小一致的供试烟粉虱20头,在0.04 mo1/L pH为7.0的磷酸缓冲液中清洗,然后将其转移到玻璃匀浆器中,加入1 mL 0.04 mo1/L pH为7.0的磷酸缓冲液 (PBS)冰浴匀浆,匀浆液在4℃下,10000 r/min离心10 min,取上清液作为酶源。
测定方法:参照Van Asperen(1962)的方法,略加改进。取待测酶液0.1 mL,加0.5 mLα-乙酸萘酯 (含终浓度为lx10-5mol/L的毒扁豆碱液)混匀,在30℃下水浴反应10 min后加入0.1 mL显色剂并终止反应,显色30 min。测定使用96孔可拆卸式酶标板,每孔加200μL反应终止液,每组反应液加3孔,借助酶标仪于595 nm测定反应终止液的OD值。以失活酶液做对照,通过α-萘酚标准曲线求出反应生成的α-萘酚的量。每个处理重复3次。
酶源蛋白质含量的测定:参照Bradford(1976)考马斯亮蓝 G250法。取0.1 mL待测酶液,加0.5 mL考马斯亮蓝G-250染色液,混合均匀后2 min至1 h内测OD595值。根据标准曲线求出酶液蛋白质含量。
羧酸酯酶比活力的计算公式:羧酸酯酶的比活力 (nmol·min-1·mgPro-1)=反应生成的 α -萘酚量/(酶液蛋白质含量*反应时间)。
2 结果与分析
烟粉虱在苦瓜、黄瓜、木瓜、水瓜、金银花、薄荷六种寄主植物上,从7月至10月的时间段内,其羧酸酯酶比活力随时间的变化趋势,如图1-6。

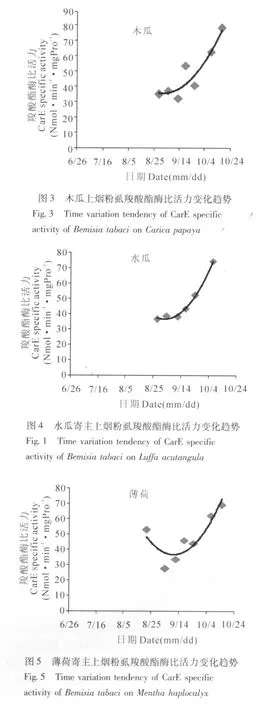
从图中可以看出,烟粉虱在不同寄主植物上羧酸酯酶比活力随时间的变化趋势,用二项式曲线进行拟合,其相互关系较为一致地呈开口向上的抛物线型。苦瓜寄主的二项式曲线为:y=0.012x2-979.0x+2E+07,相关系数R2=0.667;黄瓜寄主的二项式曲线为:y=0.012x2-1046.x+2E+07,相关系数R2=0.888;木瓜寄主的二项式曲线为:y=0.020x2-1661.x+3E+07,相关系数R2=0.867;水瓜寄主的二项式曲线为:y=0.029x2-2431.x+5E+07,相关系数R2=0.994;薄荷寄主的二项式曲线为:y=0.028x2-2303.x+5E+07,相关系数R2=0.831;金银花寄主的二项式曲线为:y=0.014 x2-1184.x+2E+07,相 关 系数R2=0.951。
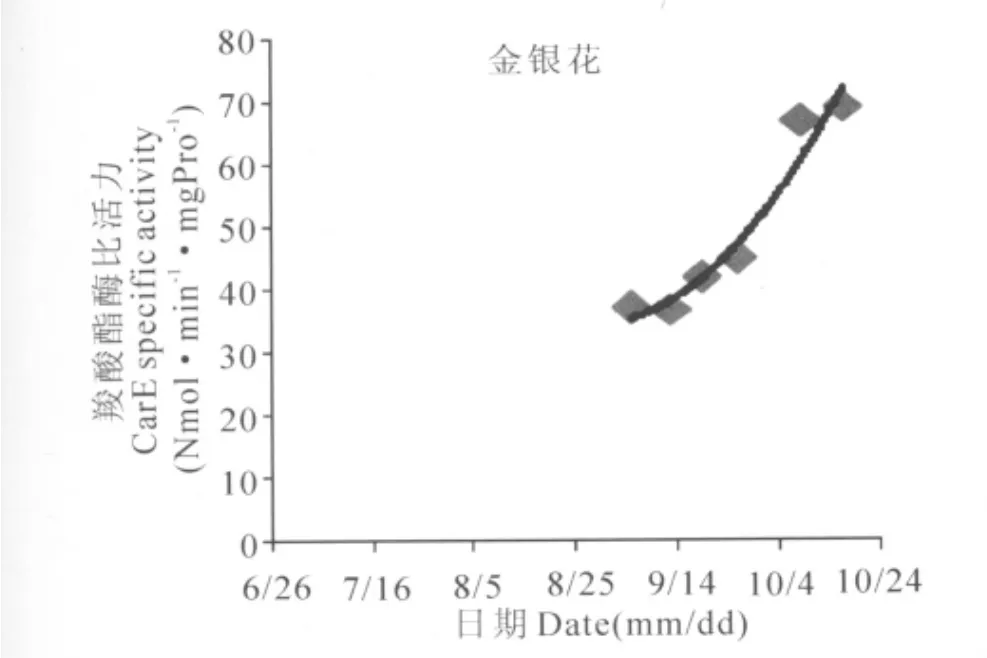
图6 金银花寄主上烟粉虱羧酸酯酶比活力变化趋势Fig.6 Time variation tendency of CarE specific activity of Bemisia tabaci on Lonicera Japonic
3 结论与讨论
烟粉虱从嗜食寄主转移到非嗜食寄主植物过程中,其解毒酶系通常会表现出活性增加的趋势,在已有研究中,多功能氧化酶、羧酸酯酶、谷胱甘肽-S-转移酶、乙酰胆碱酯酶等都受植物的影响而表现出不同程度的差异,基本上与烟粉虱的寄主适应性关系相一致 (王海鸿等,2007;王红等,2007;周福才等,2010),但是却未提到解毒酶活性与时间相关性及趋势的研究报道。
本实验的研究结果表明,烟粉虱从嗜好寄主(黄瓜)转移至非嗜好寄主 (苦瓜、木瓜、水瓜、薄荷、金银花)后,其羧酸酯酶的活性虽有显著增加的趋势 (另文发表),但其变化规律却在整体上与时间相关,这与预期不一致,其规律性在嗜好寄主黄瓜上也表现的较为典型。周福才等在转移至棉花、辣椒上后测得的烟粉虱羧酸酯酶在2~3代后达至平衡,但本实验结果的酶活性增加趋势却并非是对数曲线,而是更加符合开口向上的抛物线型,分析认为,这种趋势可能暗示了寄主植物在生长季节过程中其次生物质含量发生了规律性的变化,该变化的诱导因子可能来自于其自身的发育代谢,也可能受环境因子的影响,而与周福才等结果的不一致,则可能是由于本实验的植物材料生长在接近自然的室外环境中,更多地受外界环境的影响而使植物次生物质的代谢发生了更为复杂的变化而导致。总之,烟粉虱在寄主植物取食过程中,其包括羧酸酯酶在内的解毒酶系与植物次生物质之间可能存在着紧密的互动关系,反复而细致地研究其联动机制将有助于我们更好地理解烟粉虱的寄主植物生理适应机制。
References)
Bradford MM,1976.A rapid and sensitive methord for the quantification of microgram quantities of protein using the principle of proteindye binding.Analytical Biochem.,72:248 -254.
Brown JK,Frohlich DR,Rosell RC,1995.The sweetpotato or silverleaf whiteflies:biotype of Bemisia tabaci of a species complex.Annual Review of Entomology,40:511 -534.
Byrne DN,Bellows TS,1991.Whitefly biology.Annual Review of Entomology,36:431 -457.
Byrne FJ,Devonshire AL,1993.Insensitive acetylcholinesterase and esterases polymorphism in susceptible and resistant populations of the Tabacco Whitefly,Bemisia tabaci(Genn).Pestic Biochem.Physiol.,(45):34 -42.
Byrne FJ,Gorman KJ,Cahill M,2000.The role of B -type esterases in conferring insecticide resistance in the tabacco Whitefly,Bemisia tabaci(Genn).Pest Manag.Sci.,56(10):867-874.
Gao XW,1992.Effect of host plant on carboxylesterase activity in cotton aphid Aphis gossypii Glov.Acta Entomologica Sinica,5(3):267-272.
Gennadius P,1889.Disease of Tobacco Plantations in the Trikonia:The aleurodid of Tobacco.Ellenike Georgia,5:1-3.
Li X,Schuler MA,Berenbaum MR,2007.Molecular mechanisms of metabolic resistance to synthetic and natural xenobiotics.Annu.Rev.Entomol.,52:231 -253.
Lindroth RL,1989.Host plant alteration of detoxification activity in Papilio glaucus.Entomol.Exp.Appl.,50:29 -36.
Oliveira MRV,Henneberry TJ,Anderson P,2001.History,current status,and collaborative research projects for Bemisia tabaci.Crop Protection,20(9):709 -723.
Secker AE,Bedford ID,Markham PG,1998.Squash,a reliable field indicatior for the presence of B biotype of tabacco whitefly,Bemisia tabaci.British Crop Protection Council,837 -842.
van Asperen K,1962.A study of housefly esterase by means of a sensitive colorimetric methos.J.Insect Physiol.,8:401-416
Wang H,Wang DS,Yang YZ,Li LY,2007.Effects of host plants on enzymes related to pesticide resistance of Bemisia tabaci.Plant Pretection,33(3):36-39.[王红,王冬生,杨益众,李琳一,2007.寄主植物对烟粉虱后代种群抗性相关酶活性的影响.植物保护,33(3):36-39]
Wang HH,Li X,Zhang N,2007.The variation of carboxylesterase and acetylcholinesterase of Bemisia tabaci after host plant transferring.Plant Protection and Modern Agriculture——China Society of Plant Protection,2007 Conference Proceedings,25-30.[王海鸿,李雪,张妮,2007.寄主转换后B型烟粉虱羧酸酯酶和乙酰胆碱酯酶活力的变化,植物保护与现代农业——中国植物保护学会2007年学术年会论文集,25-30]
Zhou FC,Li CM,Zhou GS,Gu AX,Wang P,2010.Responses of detoxification enzymesin Bemisia tabaci(Genadius)to host shift.Acta Ecologica Sinica,30(7):1806-1811.[周福才,李传明,周桂生,顾爱祥,王萍,2010.烟粉虱体内几种抗性酶对寄主转换的响应.生态学报,30(7):1806-1811]
