在北京和河北局部地区Q型烟粉虱取代了B型烟粉虱
2010-11-30潘慧鹏戈大庆王少丽吴青君徐宝云张友军
潘慧鹏, 戈大庆, 王少丽, 吴青君, 徐宝云, 谢 文, 张友军
(中国农业科学院蔬菜花卉研究所,北京 100081)
烟粉虱[Bemisia tabaci(Gennadius)]属半翅目,粉虱科,是一种世界性的多食性害虫,以植物韧皮部汁液为食,通过直接刺吸为害、传播植物病毒、引起植物生理混乱、分泌蜜露诱发真菌病害给作物生产造成巨大的经济损失[1-2]。依据其寄主范围、生殖力、为害习性和传毒能力等方面的差异,至少可以分为24种生物型[3],其中B型烟粉虱是近20年来传播世界各地并暴发成灾的重要入侵害虫[3-6],在我国华北和其他地区发生并造成严重的危害[7],应用分子生物学等手段研究发现在北京市发生为害的烟粉虱主要是B型烟粉虱[8-11]。近年来,随着花卉和其他经济作物苗木的调运,Q型烟粉虱有从其起源地伊比利亚半岛向世界各地扩张蔓延的趋势。在中国,Q型烟粉虱首次于2003年在云南昆明市的一品红上发现[12],随后在北京、河南、浙江、山东、江苏等地都发现了Q型烟粉虱[13-15]。
不同生物型的烟粉虱在形态学上极其相似,但在酯酶多态性、核酸多态性等方面则存在明显的差异。而快速鉴定烟粉虱的生物型是进一步深入研究、制定相关防治措施的前提。本文利用两种方法进行烟粉虱生物型的鉴定。第1种是Shatters,Jr等[16]设计的B型烟粉虱和Q型烟粉虱的mtDNA COⅠ基因片段特异性引物,PCR产物琼脂糖凝胶电泳检测后直接判断烟粉虱的生物型。第2种是应用Simon等[17]设计的烟粉虱mtDNA COⅠ基因片段通用引物,PCR产物通过测序判定烟粉虱的生物型。上述两种方法都证实了2009年采集自蔬菜所3个试验基地的烟粉虱种群为Q生物型,同时也验证了Shatters,Jr等[16]设计的B型烟粉虱和Q型烟粉虱mtDNA COⅠ基因片段的特异性引物能够很好地鉴别B型烟粉虱和Q型烟粉虱。
1 材料与方法
1.1 供试昆虫
所用的烟粉虱种群于2009年采自蔬菜所的3个试验基地(表1);对照为本实验室饲养的B型烟粉虱种群。
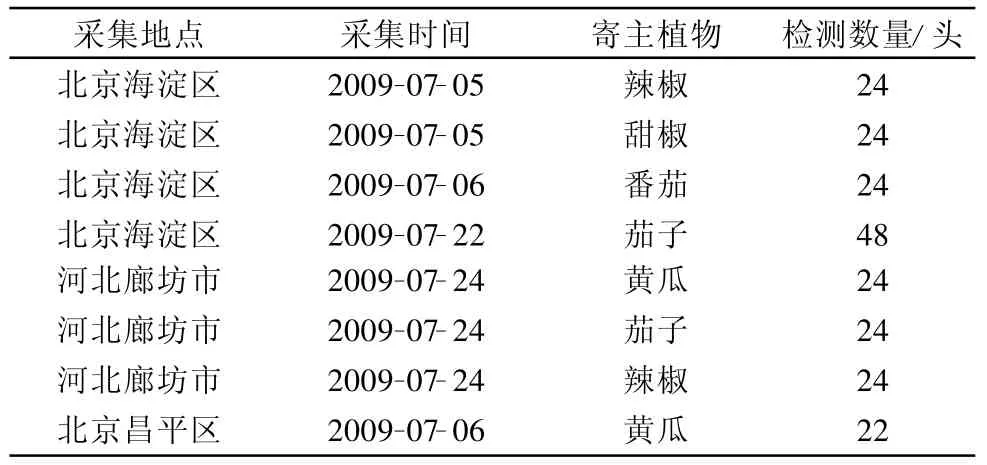
表1 2009年烟粉虱样本来源及相关信息
1.2 酶和试剂
PCR试剂盒购自TaKaRa公司;DNA提取试剂盒、PCR产物回收试剂盒购自北京百泰克生物技术有限公司;蛋白酶K、RNAase购自天根生化科技(北京)有限公司。引物由上海生工生物工程公司合成。
1.3 DNA的提取
参考滕海媛[18]的方法并略作修改,将其方法中的第1步改为吸20μL裂解液于封口膜上,将1头烟粉虱置于提取液中,用0.2 mL PCR管底部将烟粉虱充分研磨匀浆,把匀浆液吸入1.5 mL离心管中,再取40μL裂解液清洗封口膜研磨处,将清洗液与匀浆液合并。
1.4 PCR扩增及电泳
Q型烟粉虱mtDNA COⅠ基因片段PCR扩增特异性引物[16]分别为5′CTTGGTAACTCTTCTGTAGATGTGTGTT 3′和 5′CCTTCCCGCAGAAGA-AATTTTGTTC 3′;B型烟粉虱 mtDNA COⅠ基因片段PCR扩增特异性引物[16]分别为5′CTAGGGTTTATTGTTTGAGGTCATCATATATTC 3′和 5′AATATCGACGAGGCATTCCCCCT 3′,PCR热循环程序[16]为:94℃2 min,94℃30 s,64℃1 min,72℃1 min,共35个循环,最后72℃延伸10 min扩增产物大小分别为303 bp和478 bp。烟粉虱mtDNA COⅠ基因片段 PCR扩增通用引物[17]分别为 5′TTGATTTTTTGGTCATCCAGAAGT3′和 5′CTGAATATCGAGGCATTCC3′。PCR热循环程序[13]为:94℃5 min,94℃1 min,50℃1 min,72℃1 min,共34个循环,最后72℃延伸5 min,扩增产物大小为620 bp。PCR产物用1.0%琼脂糖(0.5×TBE)电泳检测,在Biorad凝胶成像仪观察,记录结果。
1.5 序列测定与比较
用B型烟粉虱和Q型烟粉虱的特异性引物扩增后的电泳检测结果判断烟粉虱的生物型;对其中部分样品应用烟粉虱mtDNA COⅠ基因片段通用引物扩增得到的PCR产物回收、纯化后送到北京擎科生物技术有限公司进行双向测序。通过同源性比较,鉴定所采集的烟粉虱种群的生物型。
2 结果与分析
2.1 Q型和B型烟粉虱特异性引物扩增结果
使用Q型烟粉虱特异性引物从3个地区采集到的烟粉虱中均扩增到1条303 bp大小的mtDNA COⅠ基因片段(图1),应用B型烟粉虱的特异性引物可以从本实验室饲养的B型烟粉虱中扩增到1条478 bp大小的mtDNA COⅠ基因片段(图1)。

图1 B型和Q型烟粉虱特异性引物对田间和温室内烟粉虱的扩增结果
2.2 mtCOⅠ通用引物扩增结果
采用mtCOⅠ通用引物从田间采集的烟粉虱及温室饲养的B型烟粉虱中均扩增到一条大小为620 bp的mtCOⅠ基因片段(图2)。经回收、纯化、测序,进行相似性比对后发现,从海淀区、昌平区及廊坊市采集到的烟粉虱种群与以色列Q型烟粉虱(登录号为AY582869)、美国佛罗里达州B型烟粉虱(登录号为EF080824)的相似性分别为99.85%、98.80%。所以2009年在海淀区、昌平区及廊坊市试验基地发生为害的烟粉虱全部为Q生物型(图3)。

3 讨论
一些学者利用对mtDNA COⅠ基因测序和RAPD标记技术对北京部分烟粉虱种群进行了鉴定,发现在北京市发生为害的烟粉虱主要是B型烟粉虱[8-11],本实验室已有的研究表明在2004年3个试验基地发生为害的烟粉虱全部为B型[19],而本研究发现在北京市海淀区、昌平区及河北省廊坊市发生为害的烟粉虱为Q型,故Q型烟粉虱已经取代了B型烟粉虱在这些地区发生为害。然而,RAPD标记为显性标记不能有效鉴别杂合子,同时易受反应条件的影响,稳定性较差;对mtDNA COⅠ基因测序能够很好地鉴别烟粉虱生物型,但是这种方法费时、费力,并且价格昂贵。在本研究中,联合使用B型和Q型烟粉虱mtCOⅠ基因特异性引物能够快速地鉴别B型与Q型烟粉虱,该结果与Shatters,Jr等[16]的研究结论是一致的。因此,对于局部地区烟粉虱B型、Q型混合种群,使用这两个特异性引物来快速鉴别这2种生物型具有一定的价值。
B型烟粉虱近20年来迅速入侵世界许多地区,并引起部分入侵地土著烟粉虱种群的下降或灭绝,从而成为最受关注的一个生物型[20-21]。而近年来Q型烟粉虱逐渐入侵到B型烟粉虱已经建立种群并暴发的地区,成为继B型烟粉虱后另一备受关注的研究对象。
Nauen等[22]对采自欧洲不同地区的烟粉虱种群进行了抗药性研究,结果发现,西班牙Almeria地区的Q型烟粉虱对吡虫啉产生了大于100倍的抗性,除了对新烟碱类药剂噻虫嗪、啶虫脒产生了较高的交互抗性,还对噻嗪酮、硫丹、久效磷等常用杀虫剂有交互抗性。另外研究还发现来自意大利和德国的Q型烟粉虱种群在吡虫啉、噻虫嗪和啶虫脒之间也存在较高的交互抗性;且Q型烟粉虱对新烟碱类杀虫剂的抗性更稳定,在未施加杀虫剂选择压力的实验室条件下,Q型烟粉虱对这类杀虫剂的抗性在两年后仍维持在较高水平[22]。Horowitz等[23]的研究发现,以色列地区的Q型烟粉虱对昆虫生长调节剂类农药如吡丙醚产生了很高的抗性。Roditakis等[24]的研究发现,采自地中海克里特岛的许多Q型烟粉虱种群都对吡虫啉和高效氯氰菊酯产生了高抗性。上述研究结果表明,Q型烟粉虱对一些化学杀虫剂能够产生很强且稳定的抗药性以及交互抗性。因此,Q型烟粉虱较B型烟粉虱治理的成本更高、难度更大。
自然条件下Q型烟粉虱竞争取代B型烟粉虱的假说很多。首先,Q型烟粉虱较B型烟粉虱对于很多种药剂具有较高的抗药性被很多学者认同[25],它可能决定了B型烟粉虱和Q型烟粉虱在田间的种群动态和扩张趋势。如Q型烟粉虱在西班牙的自然条件下较B型烟粉虱发生量大、为害严重,尤其在昆虫生长调节剂类农药如吡丙醚和新烟碱类农药广泛使用的地区,B型烟粉虱已基本被Q型取代[25]。研究者推测Q型烟粉虱所具备的高抗药性可能在其竞争取代B型过程中发挥了重要作用[25]。蔬菜所的3个试验基地常年种植蔬菜,药剂使用历史较长,尤其在过去十几年新烟碱类药剂大量用于防治粉虱类害虫。短短5年时间在蔬菜所基地作物上已经采集不到B型烟粉虱,并且目前蔬菜所番茄上发生的Q型烟粉虱对吡虫啉的抗性是室内饲养B型烟粉虱的819.04倍(未发表),所以作者推测是因为Q型烟粉虱较B型烟粉虱对于很多种药剂具有高抗性而导致Q型烟粉虱取代B型烟粉虱在蔬菜所基地作物上发生为害。但是在自然条件下不同地区烟粉虱生物型的转换与农药的关系尚需进一步深入研究。
其次,Q型烟粉虱较B型烟粉虱在一些杂草和部分农作物上具有更强的生物学优势。比如:在小花锦葵(Malva parvif lora Linn.)、荠[Capsella bursa-pastoris(Linn.)Medic.]、田芥菜[Brassica kaber(DC)]3种不同的杂草上,Q型烟粉虱雌虫产卵率、化蛹数、成虫羽化率等均比B型烟粉虱明显高[26];在不同品种的番茄上,Q型烟粉虱比B型具有更强的危害性[27];在甜椒上,Q型烟粉虱在17、33℃时平均世代历期比B型的要短[27]。另外Q型烟粉虱较B型烟粉虱具有更强忍受极端温度的能力[28],这也可能是Q型烟粉虱取代B型烟粉虱的一个重要因素。弄清楚温度与烟粉虱种群动态之间的关系对于改进控制策略和评估它在不同地区之间的扩张能力也是非常重要的。
[1] Brown JK,Czosnek H.Whitefly transmission of plant viruses[J].Advancesin Botanical Research,2002,36:65-100.
[2] Inbar M,Gerling D.Plant-mediated interactions between whiteflies,herbivores,and natural enemies[J].Annual Review of Entomology,2008,53:431-448.
[3] Perring T M.The Bemisia tabaci species complex[J].Crop Protection,2001,20:725-737.
[4] Maruthi M N,Seal S,Colvin J,et al.East Af rican cassava mosaic Zanzibar vir us-a recombinant begomovirus species with a mild phenotype[J].Archives of Virology,2004,149:2365-2377.
[5] Dennehy T J,DeGain B A,Harpold V S,et al.New challenges to management of whitefly resistance to insecticides in Arizona[R].University of Arizona,College of Agriculture Vegetable Rep ort,2005.
[6] Bird T L,Kruger K.Response of the polyphagous whitefly Bemisia tabaci B-biotype(Hemiptera:Aleyrodidae)to crop diversification-influence of multiple sensory stimuli on activity and fecundity[J].Bulletin of Entomological Research,2006,96:15-23.
[7] 张芝利.关于烟粉虱大发生的思考[J].北京农业科学,2000(增刊):1-3.
[8] Wu X X,Hu D X,Li Z X,et al.Using RAPD-PCR to distinguish biotypes of Bemisia tabaci(Homoptera:Aleyrodidae)in China[J].Insect Science,2002,9:1-8.
[9] 罗晨,姚远,王戎疆,等.利用mtDNA COI基因序列鉴定我国烟粉虱的生物型[J].昆虫学报,2002,45(6):759-763.
[10]褚栋,张友军,丛斌,等.烟粉虱不同地理种群的mt DNA COI基因序列分析及其系统发育[J].中国农业科学,2005,38(1):76-85.
[11]Zhang L P,Zhang Y J,Zhang W J,et al.Analysis of genetic diversity among different geographical populations and determination of biotypes of Bemisia tabaci in China[J].Journal of Applied Entomology,2005,129:121-128.
[12]褚栋,张友军,丛斌,等.云南Q型烟粉虱种群的鉴定[J].昆虫知识,2005,42(1):54-56.
[13]Chu D,Zhang Y J,Brown JK,et al.The introduction of the exotic Q biotype of Bemisia tabaci from the Mediterranean region into China on ornamental crops[J].Florida Entomologist,2006,89:168-174.
[14]徐婧,王文丽,刘树生.Q型烟粉虱在浙江局部地区大量发生危害[J].植物保护,2006,32(4):121.
[15]徐文华,左文惠,王瑞明,等.烟粉虱种群在江苏沿海城市市区的寄主分布与虫源性质[J].华东昆虫学报,2007,16(3):187-195.
[16]Shatters Jr R G,Powell C A,Boykin L M,et al.Improved DNA barcoding method for Bemisia tabaci and related Aley rodidae:development of universal and Bemisia tabaci biotype-specific mitochondrial cytochrome c oxidase I polymerase chain reaction primers[J].Journal of Economic Entomology,2009,2:750-758.
[17]Simon C,Frati F,Beck enbach A,et al.Evolution,weighting,and phylogenetic utility of mitochond rial gene sequences and a compilation of conserved polymerase chain reaction primers[J].Annals of the Entomological Society of America,1994,87:651-701.
[18]滕海媛,吴青君,徐宝云,等.SRAP技术研究烟粉虱遗传多样性[J].昆虫知识,2009,46(1):139-144.
[19]娄蕴萍.寄主植物对B型烟粉虱种群遗传分化的影响[D].北京:中国农业科学院研究生院,2006.
[20]Liu S S,De Barro P J,Xu J,et al.Asymmetric mating interactionsdrive widespread invasion and displacement in a whitefly[J].Science,2007,318:1769-1772.
[21]Delatte H,Duyck P F,Triboire A,et al.Differential invasion success among biotypes case of Bemisia tabaci[J].Biological Invasions,2009,11:1059-1070.
[22]Nauen R,Stumpf N,Elbert A.Toxicological and mechanistic studies on neonicotinoid cross resistance in Q-type Bemisia tabaci(Hemiptera:Aley rodidae)[J].Pest Management Science,2002,58:868-875.
[23]Horowitz A R,Denholm I,Gorman K,et al.Biotype Q of Bemisia tabaci identified in Israel[J].Phy toparasitica,2003,31:94-98.
[24]Roditakis E,Grispou M,Morou E,et al.Current status of insecticide resistance in Q biotype Bemisia tabaci populations from Crete[J].Pest Management Science,2008,65:313-322.[25]Horowitz A R,Kontsedalov S,Khasdan V,et al.Biotypes B and Q of Bemisia tabaci and their relevance to neonicotinoid and pyriproxyfen resistance[J].Archives of Insect Biochemistry and Physiology,2005,58:216-225.
[26]Nombela G,Beitia F,Muniz M.A differential interaction study of Bemisia tabaci Q-biotype on commercial tomato varieties with or without the mi resistance gene,and comparative host responses with the B-biotype[J].Entomologia Experimentalis et Ap plicata,2001,98:339-344.
[27]Muniz M,Gloria N.Differential variation in development of the B-and Q-biotypesof Bemisiatabaci(Homop tera:Aleyrodidae)on sweet pepper at constant temperatures[J].Environmental Entomology,2001,30:720-727.
[28]Bonato O,Lurette A,Vidal C,et al.Modelling temperaturedependant bionomics of Bemisiatabaci(Q-biotype)[J].Physiological Entomology,2007,32:50-55.
