蓝蝽对马铃薯甲虫低龄幼虫的捕食潜能初探
2012-11-22克尤木维勒木罗庆怀刘伟伟牟刘森冯丽凯王佩玲
舒 敏,克尤木·维勒木,罗庆怀,刘伟伟,牟刘森,冯丽凯,马 宁,王佩玲
(1.贵州师范大学生命科学学院,贵阳550001;2.石河子大学农学院,新疆石河子,832000;3.伊犁出入境检验检疫局综合技术服务中心,伊犁835000)
蓝蝽 Zicrona caerula(Linnaeus),又名纯蓝蝽、蓝盾蝽、琉璃蝽象,属半翅目蝽科益蝽亚科Asopinae。国外分布于前苏联、日本、朝鲜、蒙古、阿富汗、巴基斯坦、越南、缅甸、印度、马来西亚、印尼、伊朗、加拿大、美国、欧洲等(甘肃省自然科技资源昆虫资源库)。国内分布于海南、台湾、广东、广西、云南、福建、湖南、江西、浙江、湖北、四川、江苏、河南、山东、山西、河北、陕西、甘肃、北京、内蒙古、辽宁、新疆、黑龙江 (David and Zheng,2002)。新疆主要分布于博乐、喀什、巴楚、麦盖提、疏勒、库车、昌吉、石河子、奎屯、霍城、伊宁和阿勒泰等地 (新疆生产建设兵团农业局,1983)。其食性较杂,寄主植物包括玉米、高粱、豇豆、大豆、花生、水稻、帚石楠、大戟、甘草、毛薄荷、多枝白蕊草、桦及各种杂草等,同时作为天敌昆虫的作用也很早被发现,是蛾蝶类幼虫、叶甲幼虫和成虫的克星 (甘肃省自然科技资源昆虫资源库)。近年的调查中发现,蓝蝽在新疆地区马铃薯田间捕食检疫性害虫马铃薯甲虫Leptinotarsa decemlineata(Say)幼虫 (图8),国外也有相关报道 (张生芳译,1994a),然而,有关蓝蝽捕食性研究的报道和文献却很少 (玛丽雅,1984;姚徙义等,1995;李勤忠等,1998),系统定量的研究更为缺乏。田间观察到杂食性蓝蝽捕食入侵检疫害虫马铃薯甲虫的现象,但捕食能力如何,是否可以作为有效天敌资源并不明确。为此,本文拟通过蓝蝽对马铃薯甲虫低龄幼虫捕食能力的研究,评估其对马铃薯甲虫的潜在控制作用,以期为新疆马铃薯甲虫的可持续防控提供理论依据。
1 材料与方法
1.1 蓝蝽对马铃薯甲虫的潜在捕食能力
1.1.1 蓝蝽对马铃薯甲虫的捕食功能反应
采集的蓝蝽各虫态先在室内饲养,试验前选取龄期和体型大小一致的蓝蝽个体饥饿处理24 h,随后单头放入饲养容器中 (透明无色矿泉水瓶底高10.5 cm,直径6.2 cm),再投入带有马铃薯甲虫低龄幼虫的马铃薯茎叶;茎叶基部加脱脂棉球,容器底部铺单层滤纸,滴水保湿;顶部罩以纱网。
蓝蝽刚孵化后喜好聚集在卵块周围,达到2龄才会分开取食,故供试虫态从2龄开始,根据预试验设置各供试虫态的食物密度,具体数值见下表:

表1 蓝蝽对马铃薯甲虫的捕食功能反应各处理设置Table 1 The experiment program of functional response of Z.caerula to L.decemlineata
各处理及对照均在恒温培养箱内进行 (温度25℃ ±0.5℃,RH 70%,光照 L∶D=16∶8)。每隔24 h检查蓝蝽捕食量,详细记录每个饲养容器中马铃薯甲虫幼虫被捕食数。各处理重复10~70次。
在上述同一条件的培养箱中,用原生境猎物—兰叶甲幼虫饲养各虫态蓝蝽个体,饲养容器同上,测定蓝蝽各虫态初期体长、体宽、头壳宽及各虫态发育历期。
1.1.2 数据处理
用HollingII型圆盘方程Na=aTtN0/(1+aThN0)和 HollingⅢ型功能反应新模型Na=a×exp(-b/N0)分别拟合 (Holling,1959;丁岩钦,1980;汪世泽和夏楚贵,1988;郭玉杰,1993;金正开,2002;吴坤君等,2004),其中,Na为被捕食猎物数量,N0为初始猎物数量;Ⅱ型中a为瞬间攻击率,Tt为猎物暴露于捕食者的时间 (此次试验为1 d,因此取Tt为1),Th为处置时间;Ⅲ型中a等于猎物密度N→∞时的Na(天敌的最大捕食量),b为天敌密度P=l头时的最佳寻找密度。
1.2 蓝蝽对马铃薯甲虫和原生境中猎物的选择性
在培养容器 (同上)同时放入有5头马铃薯甲虫1龄幼虫的马铃薯茎叶和带有5头兰叶甲低龄幼虫的甘草茎叶 (甘草及兰叶甲为实验用蓝蝽的原生境植物及猎物),茎叶基部用脱脂棉球加水保湿,底端用泡沫圆盘固定,使之直立如生长状,在两植株中间投入蓝蝽成虫1头,记录其对两种幼虫的捕食情况。间隔24 h更换1次带有同等虫量的植物茎叶。5次重复,连续3 d观察蓝蝽对食物的选择性。
用Cain指数来计算捕食者的猎物选择效应,Cain指数 D=Np1×N2/Np2×N1,式中:Np1、Np2为2种猎物被捕食的数量,N1、N2为2种猎物的供试数量,D>1表示捕食者对猎物1的捕食选择性强于对猎物2的捕食选择性 (刘锦乾等,2006)。
2 结果与分析
2.1 蓝蝽各虫态形态特征
卵:圆柱形,蓝色有光泽。卵盖周围有一圈放射状突出物,卵盖与卵体界限明显,孵化前卵颜色发红,孵化时若虫从破开的卵盖一侧爬出。
若虫:共5龄,各龄期体长、体宽、头宽有明显区别。若虫除头部、胸部、附肢和翅芽全部为蓝色外,腹部主要为红色,腹部腹板正面有四个蓝色横斑,从前向后逐渐变小,腹部侧板也有蓝色横斑,每一侧板1个,每侧各6个,腹部末端蓝色 (新疆生产建设兵团农业局,1983)。若虫初孵和每次蜕皮后,初期颜色均为橘红色,随后虫体大部逐渐变成红紫色,再过度到蓝紫色,最后变成蓝色。
成虫:椭圆形,蓝、蓝黑或紫黑色,有光泽,密布同色刻点。头略呈梯形,中叶与侧叶等长,触角5节,棒状,蓝黑色,有细毛。喙4节,蓝黑色末端伸达中足基节后缘。前胸背板侧角圆微外突。小盾片三角形,端部圆。前翅膜片超出腹末,棕色。足与体同色,侧接缘几不外露。雌虫腹板较粗糙雄虫则较光滑。
在新疆5~9月田间各虫态均有,世代重叠。其各虫态特征比较见表2和附图1-8。
2.2 蓝蝽对马铃薯甲虫的潜在捕食能力
2.2.1 HollingII圆盘方程拟合结果
利用HollingII圆盘方程拟合蓝蝽对马铃薯甲虫1龄幼虫的功能反应结果见表3。R、R2为1/Na和1/N0的相关系数及决定系数,其值显示蓝蝽各虫态捕食量与猎物密度呈显著正相关。蓝蝽若虫处理完1头猎物所花费的时间 (Th)及最大日平均捕食量 (1/Th)均随龄期的增加呈规律性下降或增加,成虫则介于4龄和5龄若虫之间。瞬时攻击效率 (a)则为3龄>2龄>4龄>5龄,成虫小于2龄若虫而大于4龄若虫。
a/Th可以用来衡量天敌对害虫的控制能力,其比值越大,天敌对害虫的控制能力越强 (周集中和陈常铭,1986;牟吉元等,1997)。表3可见,随着龄期增加蓝蝽对害虫的控制能力增强,其控制能力2龄<3龄<4龄<5龄<成虫。

表2 蓝蝽成虫及各若虫期特征Table 2 The feature of adult and nymph of Z.caerula

表3 蓝蝽对马铃薯甲虫1龄幼虫的HollingII型功能反应Table3 TheHollingIIfunctionalresponseofZ.caerulatothe1stinstarlarvaeofL.decemlineata
各理论值与实际值χ2适合性检验P<0.05,差异不显著,说明模型能够反映蓝蝽各虫态对马铃薯甲虫1龄幼虫的捕食变化规律。
2.2.2 寻找效应的估算
根据Holling的理论,寻找效应 (S)与寄主密度 (N0)的关系是:S=a/(1+a+ThN0)(任月萍和刘生祥,2006)。由此得出蓝蝽若虫及其成虫在不同食物密度下的寻找效应 (图1):随着食物密度的增加,各龄期若虫及成虫的寻找效应均逐渐下降;在食物密度相同的条件下,成虫及高龄若虫的寻找效应为成虫>5龄>4龄,不同虫态同食物密度下或同一虫态不同食物密度下的寻找效应下降趋势较一致;龄期较小时 (2、3龄),随食物密度的增加,寻找效应的下降趋势逐渐变缓。

图1 蓝蝽寻找效应与食物密度的关系Fig.1 Therelationshipbetweenhuntingeffectandpreydensityof Z.caerula
2.2.3 HollingⅢ型新模型拟合结果
HollingⅢ型功能反应新模型Na=a×exp(-b/N0)拟合结果见表4。R、R2显示lnNa和1/N0显著正相关。最大日平均捕食量a随着若虫龄期的增加而增大,成虫的捕食量大于若虫的捕食量,与圆盘方程中成虫的该值小于5龄而大于4龄若虫的结果不同,其值均比圆盘方程拟合值要小。b为最佳寻找密度,若虫的最佳寻找密度随龄期增大而增大,成虫的最佳寻找密度与4龄幼虫最为接近,根据最佳寻找密度可以确定利用蓝蝽防治马铃薯甲虫时的最佳益害比,2~5龄若虫及成虫分别为1∶2、1∶4、1∶13、1∶14、1∶13。
卡方检验P<0.05,理论值与实测值差异不显著,说明模型拟合较好。
2.2.4 成虫对马铃薯甲虫2龄幼虫的捕食
对1/Na和1/N0,lnNa和1/N0进行相关性分析,所得R分别为0.445、0.461,小于0.5,因此不能用HollingⅡ和HollingⅢ型模型进行拟合。但是成虫对马铃薯甲虫2龄幼虫的捕食符合多项式y=0.007x2-0.311x+7.419,R2=0.9167(见图2)。

图2 蓝蝽成虫对2龄马铃薯甲虫幼虫的捕食曲线Fig.2 ThepredationcurveofadultofZ.caerulato2ndinstarL.decemlineatalarvae
2.2.5 蓝蝽对马铃薯甲虫和原生境猎物的选择效应
蓝蝽对原生境猎物—兰叶甲的平均捕食量为4.60头/天,对马铃薯甲虫幼虫的平均捕食量为0.67头/天,因此,Cain指数D=4.60×5/0.67×5=6.87,大于1,蓝蝽对兰叶甲的捕食选择性强于对马铃薯甲虫的捕食选择性。
试验观察发现在投入食物以后,蓝蝽能快速和准确的寻找到兰叶甲。在兰叶甲够满足食物量的情况下,蓝蝽仅取食兰叶甲,只有食物量不足或兰叶甲较隐蔽的情况下,蓝蝽才会取食马铃薯甲虫幼虫,其捕食情况见表5。
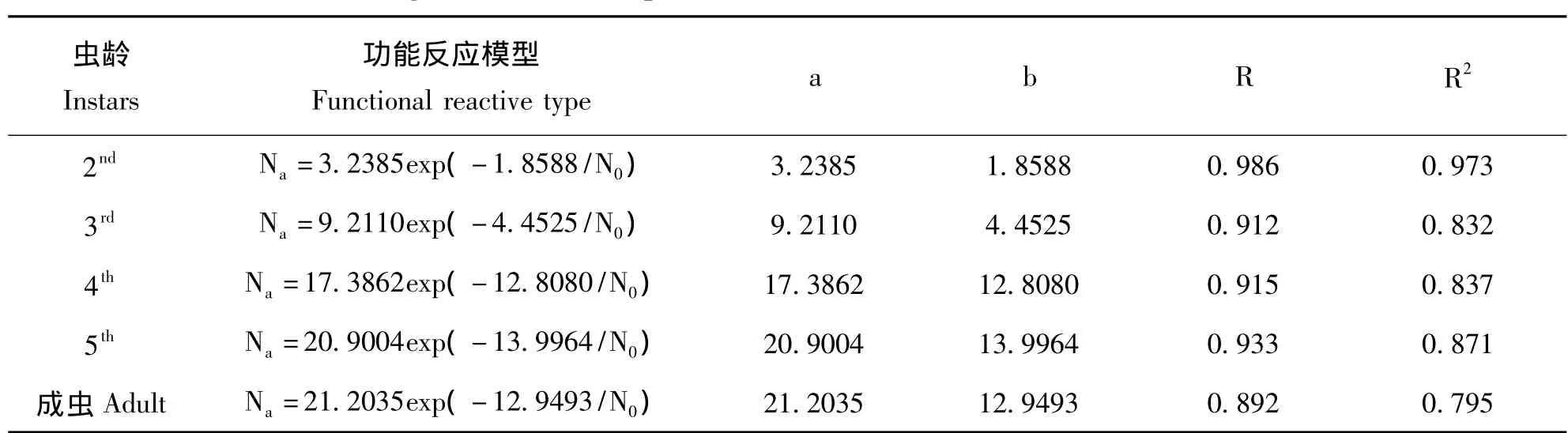
表4 蓝蝽对马铃薯甲虫1龄幼虫的HollingⅢ型功能反应Table4 TheHollingⅢ functionalresponseofZ.caerulatothe1stinstarlarvaeofL.decemlineata

表5 蓝蝽对马铃薯甲虫幼虫和原生境猎物的取食选择Table5 ThefoodselectivityofZ.caerulatooriginalhabitatpreyandL.decemlineata
3 结论与讨论
蓝蝽是全北区和东洋区的区系共有种,欧洲地区发现其能捕食马铃薯甲虫卵,一昼夜平均取食11~12粒,最多达28粒,但该种昆虫在马铃薯上相当少见 (张生芳译,1994a),尽管调查同样显示蓝蝽在马铃薯田间虫口较小,但经HollingⅡ圆盘方程和HollingⅢ型模型拟合,其2~5龄若虫及成虫的日最大捕食量分别为:3.497、10.2506、22.0932、30.6014、28.7748 和 3.2385、9.2110、17.3862、20.9004、21.2035。两种模型拟合结果虽然有所差异,但是均显示蓝蝽对马铃薯甲虫1龄幼虫具有一定的控制作用,4、5龄若虫及成虫的捕食量均在17头以上,且室内饲养也发现其对幼虫的捕食量明显大于卵。蓝蝽在新疆的发生时间主要集中在5~9月,在新疆马铃薯甲虫主要发生地多有分布,这与田间马铃薯甲虫的发生具有时间上和空间上的一致性。
蓝蝽寄主植物较广泛,但并不取食马铃薯,因此通过寄主植物与马铃薯邻作的方式可以诱集蓝蝽进入马铃薯田间。人工投放2~5龄若虫及成虫时,最佳益害比分别为 1∶2、1∶4、1∶13、1∶14、1∶13。但是,猎物选择效应显示,蓝蝽对原猎物兰叶甲的选择性明显大于马铃薯甲虫,因此,在利用原始寄主植物作为诱集植物时,怎样合理布局和管理也是一个需要研究的问题。作者认为,可用兰叶甲的寄主植物与马铃薯邻作、间作或套种进行对比试验,如有可能,可用兰叶甲的多种原寄主植物进行试验,然后对其农业生产的收益进行综合评估后再确定马铃薯的种植模式。
马铃薯甲虫是世界性的检疫性害虫,国内外的调查研究目前没有发现专性的捕食者,这是生物防治中的重要瓶颈。尽管国外研究发现斑腹刺益蝽、二点益蝽对马铃薯甲虫的控制作用较好(张生芳译,1994b),新疆的调查研究也发现中华通草蛉、原姬蝽、多异瓢虫、蓝蝽等对马铃薯甲虫均有一定的控制作用,但在实际生产中还没有得到广泛应用和推广。所以系统评估各天敌在田间的实际控制作用,除了室内捕食功能反应研究外,天敌干扰效应的评估、田间试验观察及有效天敌的人工保护及饲养等研究是亟待进行的。
致谢:本研究得到了新疆农业科学院植物保护研究所郭文超研究员的支持与帮助,昆虫的采集和饲养过程中得到了石河子大学农学院2008级研究生杨涛、2010级研究生王银方,贵州师范大学2011级研究生孙翠英的帮助,论文的修改过程中,得到了中国科学院新疆生态与地理研究所吕昭智研究员的悉心指导,附图中图8为南京农业大学昆虫系/农业部病虫害监测与治理重点开放实验室卢伟平所提供,在此一并表示感谢!

References)
David AR,Zheng LY,2002.Checklist and nomenclatural notes on the Chinese Pentatomidae(Heteroptera)I.Asopinae.Entomotaxonomia,24(2):107-115.
Ding YQ,1980.Principle and Application of Insects Population Mathematics Ecology.Beijing:Science Publishing Company Press.190-210.[丁岩钦,1980.昆虫种群数学生态学原理与应用.北京:科学出版社.190-210]
Guo YJ,1993.A review on functional response models for predation.Chin.J.Biol.Contr.,9(1):40 -45.[郭玉杰,1993.捕食作用中的功能反应模型.生物防治通报,9(1):40-45]
Holling CS,1959.Some characteristics of simple type of predation and parasitism.J.Can.Entomol.,91(7):385 -398.
Insect databases of natural science and technology resources of Gansu province. [甘肃省自然科技资源昆虫资源库.http://www.gsagr.ac.cn/zypt/kunchong/showsoft.asp?id=134]
Jin ZK,2002.Study on statistic model of functional response of predator to prey.J.Environ.Entomol.,24(4):175-179.[金正开,2002.捕食者-猎物功能反应的统计模型研究.昆虫天敌,24(4):175-179]
LI QZ,Zhang HD,Niu BZ,Li XB,1998.Two kinds of natural enemies of poplar leaf red beetles was founded for the first time in Xinjiang.China Forestry,(4):42.[李勤忠,张互朵,朱保志,李学斌,1998.新疆首次发现杨赤叶甲的两种天敌.中国林业,(4):42]
Liu JQ,Li YY,Zhang HJ,Qi YQ,2006.Research on the functional responses of Coccinella septempunctata L.to prey on Stephanitisanagustata Bu.Sp.Nov.J.Northwest A.﹠ F.Univ.(Nat.Sci.Ed.),34(3):111-113.[刘锦乾,李玉英,张海江,齐跃强,2006.七星瓢虫成虫对狭冠网蝽的捕食功能反应研究.西北农林科技大学学报 (自然科学版),34(3):111-113]
Ma LY,1984.Preliminary observation of predation habits of Ziocrona caerulea L.in Tarim,conservation area of Populus euphratica.Journal of Xinjiang University(Science& Engineering),(1):78-79.[玛丽雅,1984.塔里木胡杨林保护区兰蝽 Ziocrona Caerulea L.捕虫习性的初步观察.新疆大学学报 (自然科学版),(1):78 -79]
Mou JY,Xu HF,Li HG,1977.Insect Ecology and Agricultural Pests Forecasting.Beijing:China Agricultural Science Press.56 - 57.[牟吉元,徐洪富,李火苟,1997.昆虫生态与农业害虫预测预报.北京:中国农业科学出版社.56-57]
Ren YP,Liu SX,2006.Research of the predated and hunted functional response of Propylaea japonica to wheat aphid.J.Agric.Sci.,27(1):20-21,33.[任月萍,刘生祥,2006.龟纹瓢虫对麦蚜的捕食功能反应及寻找效应研究.农业科学研究,27(1):20 -21,33]
The Xinjiang Production and Construction Corps Agriculture Bureau,1983.Atlas of Crop Pests and Their Natural Enemies in the Xinjiang Production and Construction Corps Agricultural Reclamation Area.Xinjiang:The Xinjiang Production and Construction Corps Agriculture Bureau Press.93.[新疆生产建设兵团农业局,1983.新疆生产建设兵团垦区农作物害虫及其天敌图册.新疆:新疆生产建设兵团农业局.93]
Wang SZ,Xia CG,1988.New model of Holling-Ⅲ functional response.Chin.J.Ecol.,7(1):1 -3.[汪世泽,夏楚贵,1988.Holling-Ⅲ型功能反应新模型.生态学杂志,7(1):1-3]
Wu KJ,Sheng CF,Gong PY,2004.Equation of predator functional response and estimation of the oarameters in it.Entomol.Knowl.,41(3):267-269.[吴坤君,盛承发,龚佩瑜,2004.捕食性昆虫的功能反应方程及其参数的估算.昆虫知识,41(3):267 -269]
Yao TY,Chang R,Liu JH,Li Y,Bao HS,Si GY,1995.Natural enemies list of sugarbeet pests in Shanxi Province(continuation).China Sugarbeet,(2):59.[姚徙义,常儒,刘吉海,李元,鲍恒生,司贵雨,1995.山西省甜菜害虫天敌昆虫名录(续).中国甜菜,(2):59]
Zhang SF translate,1994a.Insectivorous natural enemies of Leptinotarsa decemlineata in Europe.Plant Quaran.,8(5):276 - 278.[张生芳译,1994a.欧洲的马铃薯甲虫食虫天敌.植物检疫,8(5):276 -278.]
Zhang SF translate,1994b.Natural enemies of Leptinotarsa decemlineata introduced from America to Europe.Plant Quaran.,8(6):342-344.[张生芳译,1994b.由美洲引入欧洲的马铃薯甲虫天敌.植物检疫,8(6):342-344]
Zhou JZ,Chen CM,1986.Predation of wolf spider,Lycosa pseudoannulata on brown planthopper,Nilaparvata lugens,and its simulation model.I.Functional response.Chin.J.Biol.Contr.,2(1):229.[周集中,陈常铭,1986.拟环纹狼蛛对褐飞虱的捕食作用及其模拟模型的研究Ⅰ.功能反应.生物防治通报,2(1):229]
