3T3-L1前体脂肪细胞分化过程中G蛋白偶联受体48的表达研究
2012-10-30孙海燕彭永德董维平王煜非范能光刘瑞赵立丁晓颖
孙海燕 彭永德 董维平 王煜非 范能光 刘瑞 赵立 丁晓颖
肥胖是由脂肪细胞的数量增多和体积增大所致,而脂肪细胞的增多是由于前脂肪细胞不断分化所引起,近年来脂肪细胞的分化调控已成为肥胖及其相关疾病的研究热点。
前期研究结果提示G蛋白偶联受体48(G protein coupled receptor 48,GPR48)基因敲除小鼠脂肪组织含量明显减少,但该受体参与脂肪细胞的增殖与分化机制不明[1]。过氧化物酶体增殖体激活受体g2(peroxisome proliferatoractivated receptorγ2,PPARg2)和 CCAAT 增强子结合蛋白(CCAAT Enhancer binding protein CEBP α,C/EBPα)是调节脂肪形成的主要转录因子,在脂肪细胞分化中起着决定性作用[2-4]。因此探讨GPR48与核转录因子在脂肪细胞分化过程的表达及调控关系具有重要意义。
本研究观察前体脂肪细胞分化过程中GPR48、PPARγ2和C/EBPα的表达水平变化,探讨GPR48参与脂肪细胞分化过程及其与转录因子调控机制的关系。
材料与方法
一、材料
小鼠胚胎成纤维细胞(3T3-L1)株,购自中国科学院上海细胞研究所。高糖Dulbecco's Modified Eagle Medium(DMEM)培养基和胎牛血清(美国Gibco公司),地塞米松和1-甲基-3-异丁基-黄嘌呤(IBMX)(美国Sigma公司),合成人胰岛素(美国礼来公司),逆转录一聚合酶链反应试剂盒(美国promega公司),SYBR Green PCR master mix(日本Takara公司 ),Trizol(美国Invitrogen公司),荧光定量PCR仪器(美国Bio-Rad公司)。
二、方法
(一)3T3-L1前脂肪细胞的培养和诱导
将3T3-L1细胞接种在6孔培养板,在含10﹪胎牛血清和1﹪青链霉素的高糖DMEM培养液37℃、5﹪CO2培养,当细胞生长至融合,加细胞分化诱导A液(0.5 mmol/L IBMX、1 µmol/L地塞米松、10 µg/ml胰岛素),继续培养 48 h,更换培养液,加诱导B液(10 µg/ml胰岛素)培养48 h后更换培养液,以后每2 d更换培养液,分别在诱导前0 d和诱导后2、3、6、10和14 d收集细胞。
(二)总核糖核酸(ribonucleic acid,RNA)提取和Real-time PCR
按照Trizol说明提取细胞总RNA,紫外分光光度计定量,取2 μg RNA加入随机引物,反转录成cDNA。在荧光定量PCR仪器上进行扩增反应。PCR 反应体系:cDNA 2.0 μl,SYBR Green Master 10 μl,上 下 游 引 物(2.5 Μm)各 2 μl,用DEPC-H2O补足至20 μl体积。PCR反应条件:95℃ 5 min预变性,95℃15 s、60℃1 min,循环40次。融解曲线显示为单峰。应用2-△△Ct法进行相对定量分析。实验重复3次,每组设3孔重复(N = 3,n= 3)。引物由上海博尚生物工程技术服务公司合成,引物序列见表1。

表1 Real-time PCR引物
三、统计学分析
采用SPSS 11.0统计软件对实验结果进行统计分析。受体表达水平以±s表示。不同时间表达水平比较采用随机区组F检验。以P< 0.05为差异有统计学意义。
结 果
一、3T3-L1前脂肪细胞分化前后形态变化
体外培养的3T3-L1前脂肪细胞在分化前呈梭形,无脂滴纤维母样细胞形态。诱导分化的第3~4天细胞变大变圆,部分细胞内出现脂滴,随着分化时间的延长细胞内脂滴富集,到诱导分化的1周左右,80﹪细胞呈成熟脂肪细胞形态,细胞呈圆形,胞浆中富含脂滴,围绕核周呈戒环样结构。(图1,2)
二、3T3-L1前脂肪细胞分化过程中GPR48、PPARγ2和C/EBPα的表达水平变化
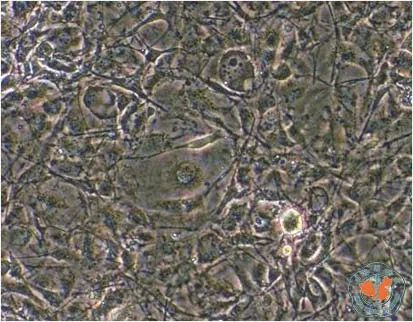
图1 诱导分化前3T3-L1前脂肪细胞 (×100)
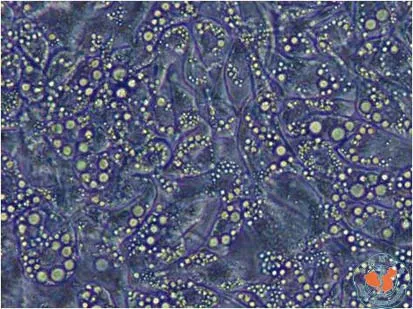
图2 诱导分化1周脂肪细胞 (×100)
比较前体脂肪细胞不同分化阶段GPR48表达水平变化,比较脂肪细胞分化标志基因PPARγ2和C/EBPα表达水平变化。由图3可知,与诱导前期相比,GPR48基因在诱导分化第2天表达显著上调,差异有统计学意义(t= 4.12,P= 0.015),分化第3天仍较高表达,差异有显著统计学意义(t= 6.21,P= 0.003)。分化第6天至第14天下调,与分化前比较差异无统计学意义。图4结果显示,PPARγ2表达在诱导分化后明显上调,分化第6天达高峰,第10~14天持续处于较高水平,各时段表达水平与诱导前期相比差异均有统计学意义(t值在4.17~22.65间,P均 < 0.01)。图5结果显示,C/EBPα表达在诱导分化后明显上调,分化后第3天达高峰,第6~10天持续保持在较高水平,与诱导前期相比各时段表达水平差异均有统计学意义(t值在4.38~13.87间,P均< 0.01),第14天趋于下调,与分化前比较差异无统计学意义(表2)。

图3 3T3-L1前脂肪细胞分化过程中GPR48表达
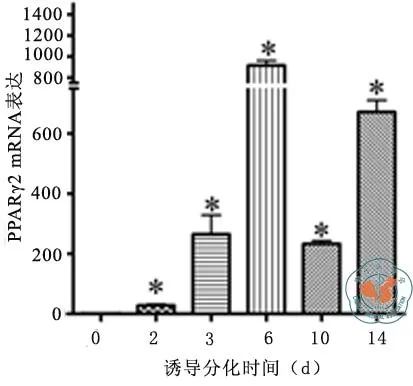
图4 3T3-L1前脂肪细胞分化过程中PPARγ2表达

图5 3T3-L1前脂肪细胞分化过程中C/EBPα表达
讨 论
肥胖是冠心病、高血压、2型糖尿病及高脂血症等许多严重疾病的共同致病基础。肥胖的形成主要包括2个方面,脂肪细胞数量的增加和脂肪细胞体积增大。脂肪细胞分化是决定脂肪细胞体积数量的关键因素,脂肪分化调控也是研究肥胖及其相关疾病发生机制的热点问题。3T3-L1前体脂肪细胞通过经典鸡尾酒诱导方案可以分化为脂肪细胞,是目前体外研究脂肪细胞分化应用最为广泛的细胞系。PPARγ和C/EBPα是调节脂肪形成最主要的转录因子,它们调控着许多中间型转录因子[2-4]。
表2 3T3-L1前脂肪细胞分化过程中PR48、PPARγ、C/EBPα的表达水平变化(±s)

表2 3T3-L1前脂肪细胞分化过程中PR48、PPARγ、C/EBPα的表达水平变化(±s)
注:与诱导前比较aP < 0.05
mRNA相对表达 诱导前 诱导第2天 诱导第3天 诱导第6天 诱导第10天 诱导第14天 F值 P值GPR48 1.01 ± 0.14 2.91 ± 0.78a 2.39 ± 0.36a 1.09 ± 0.28 1.28 ± 0.15 1.05 ± 0.23 13.2340.000 PPARγ 1.02 ± 0.2227.95 ± 2.32a265.11 ± 109.69a916.76 ± 79.23a233.00 ± 17.73a671.98 ± 67.31a104.3400.000 C/EBPα 1.00 ± 0.0658.21 ± 16.85a172.36 ± 21.39a 78.00 ± 14.36a 37.11 ± 2.78a 15.34 ± 21.39 48.2850.000
尽管PPARg2和C/EBPα对脂肪细胞分化的重要性已得到广泛认可,然而二者如何调控脂肪细胞分化的分子机制仍知之甚少。本研究数据显示PPARγ2和C/EBPα的表达在诱导分化后明显上调,以后持续保持在较高水平并趋于稳定。二者的表达与脂肪细胞分化、脂质积聚过程相一致。
GPR48是近年发现的G蛋白偶联受体家族中功能未知的孤儿受体,属于糖蛋白激素受体。近期对GPR48敲除小鼠研究均发现纯合敲除小鼠体型消瘦,腹腔网膜和性腺旁白色脂肪发育不良,脂肪组织含量明显减少,并伴有生育缺陷和围生期死亡[1,5]。GPR48在体内分布广泛,白色脂肪、棕色脂肪、肝脏、肌肉、性腺和肾上腺均有高丰度表达,这些组织对于机体的脂肪代谢和能量平衡意义重大。GPR48在脂肪组织高丰度表达,跨膜蛋白GPR48是否通过核转录因子PPARγ2参与脂肪细胞的增殖与分化机制,本研究数据提供了跨膜蛋白GPR48和转录因子PPARγ2、C/EBPα基因在脂肪细胞分化不同时期的表达,GPR48表达高峰出现于分化初期,早于转录因子PPARγ2、C/EBPα的表达高峰,GPR48可能在分化早期阶段参与调控脂肪细胞分化。
1 Li XY, Lu Y, Sun HY, et al. G protein-coupled receptor 48 upregulates estrogen receptor α expression via cAMP/PKA signaling in the male reproductive tract[J]. Development,2010, 137(1):151-157.
2 Odegaard JI, Ricardo-Gonzalez RR, Goforth MH, et al.Macrophage-specific PPARgamma controls alternative activation and improves insulin resistance[J]. Nature, 2007,447(7148):1116-1120.
3 Bai P, Houten SM, Huber A, et al. Peroxisome proliferatoractivated Receptor (PPAR)-2 controls adipocyte differentiation and adipose tissue function through the regulation of the activity of the retinoid X receptor/PPAR{gamma} heterodimer[J]. J Biol Chem, 2007,282(52):37738-37746.
4 Zuo Y, Qiang L, Farmer SR. Activation of CCAAT/enhancer-binding protein (C/EBP) alpha expression by C/EBP beta during adipogenesis requires a peroxisome proliferator-activated receptor-gamma-associated repression of HDAC1 at the C/ebp alpha gene promoter [J].J Biol Chem, 2006, 281(12):7960-7967.
5 Tzameli I, Fang H, Ollero M, et al. Regulated production of a peroxisome proliferator-activated receptor-gamma ligand during an early phase of adipocyte differentiation in 3T3-L1 adipocytes [J]. J Biol Chem, 2004, 279(34):36093-36102.
6 Mazerbourg S, Bouley D, Sudo S, et al. Leucine-rich repeat-containing, G protein-coupled receptor 48 null mice exhibit intrauterine growth retardation associated with embryonic and perinatal lethality[J]. Mol Endocrinol,2004, 18(9):2241-2254.
