Cia RH生物信息学分析及其在不同猪链球菌血清型中的分布
2012-09-26冯晓丹顾平清唐家琪
冯晓丹,王 晶,顾平清,唐家琪
猪链球菌2型(Streptococcus suis 2,S.suis 2)是一种重要的人兽共患病病原体[1]。S.suis 2感染可致猪急性败血症、脑膜炎、关节炎、心内膜炎及急性死亡,并且可通过伤口和呼吸道等传播途径感染人类,造成发病和死亡[2]。1998年和2005年我国江苏省和四川省分别暴发大规模S.suis 2感染疫情,患者出现链球菌中毒性休克综合征(Streptococcal toxic shock syndr o me,STSS),病情凶险,病死率高(62.7%~81.3%),具体致病机制尚不明确[3]。
二 元 信 号 转 导 系 统 (Two-co mponent signal transduction systems,TCSTSs)是广泛存在于细菌中的调控单元,由细胞膜上的组氨酸激酶(Histidine kinase,HK)和细胞内的反应调节因子(Response regulator,RR)两部分组成,在病原菌感染宿主的过程中能调控多种毒力因子的表达以完成其致病过程[4]。我实验室已针对 TCSTSs与S.suis 2致病的关系进行了初步的研究。李明等[5]利用基因敲除技术证实位于89 kb毒力岛上的TCSTS Sal KR在S.suis 2致病过程中发挥重要作用,动物实验结果显示,基因敲除株对仔猪不致病。对孤因子Cov R的研究显示,该蛋白对S.suis 2的不同毒力因子进行全局负调控;仔猪攻毒实验证实,该蛋白编码基因失活后菌株毒力增强[6]。为研究TCSTS Cia RH在S.suis 2致病过程中所起的作用,我们之前对cia R基因进行敲除,并比较了敲除株和野生株在生物学及致病方面的差异,结果显示,敲除株的生长速度减缓,抗氧化能力减弱,耐酸能力下降,但动物实验显示敲除株对小鼠的毒力没有发生明显改变[7]。
针对病原菌TCSTSs的研究目前主要集中在其调控的基因上,对于TCSTSs本身的结构特征关注较少,而相似的结构特征可能具有相似的信号感受方式及调控模式。为深入了解S.suis 2 Cia RH感受信号及调控基因的方式,本研究利用生物信息学方法分析其组分蛋白的结构域。同时,利用PCR及序列分析手段评估Cia RH在不同血清型S.suis中的分布情况。
1 材料和方法
1.1 菌株 31个S.suis血清型标准株由加拿大蒙特利尔大学 Marecelo Gottschalk教授惠赠;另外的9株2型S.suis包括:T15(cps2+sl y+)来自荷兰,S735(cps2+mr p+epf+sl y+)来自加拿大,SS2-N(cps2+mr p+sl y+)来自德国,98012(cps2+mr p+epf+sl y+)、Habb (cps2+mr p+epf+sl y+)、HA68(cps2+ep f+)及98 T003(cps2+mr p+ep f+sl y+)为中国江苏分离菌株,266及05ZYH33(cps2+mr p+ep f+sl y+)为中国四川省资阳市分离菌株。
1.2 试剂及仪器 Taq DNA聚合酶为Ta Ka Ra公司产品;1 kb DNA分子量标准为Fer mentas公司产品;PCR扩增仪为HYBAID公司产品。
1.3 Cia RH生物信息学分析 使用NCBI网站的BLASTN获得S.suis 2 Cia RH编码基因cia H 及cia R同源核酸序列;BLASTP获得其组分Cia H及Cia R 同 源 蛋 白 氨 基 酸 序 列 (http://blast.ncbi.nl m.nih.gov/)。多序列比对使用 Cl ustal X2[8]进行,利用 Genedoc(http://www.psc.edu/biomed/genedoc)修饰并输出比对后的序列。序列进化树使用MEGA5中的NJ法构建[9]。分别使用Signal P4.0[10]和 T MH MM 2.0(http://www.cbs.dt u.dk/services/T MH MM/)在线对 Cia RH 组分蛋白的氨基酸序列进行信号肽和跨膜区预测。利用Batch CD-Search工具[11]预测 Cia RH 组分蛋白的结构域。蛋白二级结构预测使用PSIPRED v3.0在线软件(http://bioinf.cs.ucl.ac.uk/psipred/)。
1.4 PCR方法检测cia R基因在猪链球菌不同血清型中的分布 以S.suis 2强毒株05ZYH33为模板,设计cia R 基因的上下游引物,cia1:5′-GCCGGATCCATGATGATTAAAATATTG-3′; cia2:5′-CGCAAGCTTTTAGGCATTTTTTAGAAT-3′。引物由上海赛百盛基因技术有限公司合成。PCR反应条件为:95 ℃,5 min;94 ℃,30 s;52 ℃,40 s;72℃,1 min。共反应30个循环。电泳检测扩增产物。
2 结 果
2.1 Cia RH 的一般特性 在S.suis 2强毒株05ZYH33中,Cia RH的 HK蛋白Cia H (Gen Bank accession No.YP_001198460.1)基因长度为1 380 bp,GC含量为36.67%,蛋白长度为459 aa,预测的分子质量为52.0 k D。RR蛋白Cia R(Gen Bank accession No.YP_001198461.1)基因长度为675 bp,GC含量为35.71%,蛋白长度为224aa,预测的分子质量为25.4 k D。两个基因在S.suis 2染色体上位置毗邻,且有8 bp的重叠,这与TCSTSs编码基因的结构特征相符。
2.2 Cia RH同源蛋白比对 利用 NCBI网站BLASTP进行同源检索,结果显示,S.suis 2 Cia RH的同源蛋白在链球菌属不同菌种中广泛分布。其中,Cia R与变形链球菌UA159(S.mutans UA159)、肺炎链球菌R6(S.pneu moniae R6)、无乳链球菌2603 V/R(S.agal actiae 2603 V/R)及化脓链球菌 M1 GAS(S.p yogenes M1 GAS)同源蛋白的氨基酸序列一致性(identity)依次为90%、88%、87%及82%;相比之下,Cia H与这些菌株的同源蛋白相似性较低,一致性分别为48%、49%、47%及49%。将S.suis 2 Cia H及Cia R与其各自同源蛋白进行多序列比对,结果显示,不同菌种Cia H的保守区域主要集中在后半部分(C-端),前半部分(N-端)保守型较低(图1);而Cia R的全长序列均较为保守(图2)。与其他菌种的Cia R序列相比,S.pneumoniae同源序列在N-端明显多出约20个氨基酸的序列,这可能是由于在蛋白编码框预测时选择了不同的起始密码子所致。
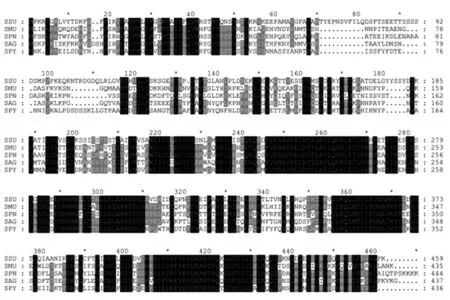
图1 Cia H及其同源蛋白氨基酸序列比对Fig.1 Align ment of amino acid sequence of Cia H with its orthologs The amino acid sequences of Cia H were derived from SSU (S.suis 05ZYH33),SMU (S.mutans UA159),SPN(S.pneumoniae R6),SAG (S.agal actiae 2603 V/R),and SPY (S.pyogenes M1 GAS).

图2 Cia R及其同源蛋白氨基酸序列比对Fig.2 Alignment of amino acid sequence of Cia R with its orthologs The amino acid sequences of Cia R were derived from SSU (S.suis 05ZYH33),SMU (S.mutans UA159),SPN (S.pneu moniae R6),SAG (S.agal actiae 2603 V/R),and SPY (S.pyogenes M1 GAS).
2.3 Cia RH蛋白组分结构预测 信号肽预测结果显示,Cia H及Cia R蛋白均不含信号肽序列。Cia H的N-端含有两个跨膜区,中间存在168个氨基酸的间隔(38aa~205aa),推测为胞外的信号感应区(Sensor)。结构域分析结果显示,Cia H的胞内(C-端)含有HK中高度保守的信号传递(Transmitter)结构域 His KA及 HATPase_C;Cia R蛋白N-端为保守的磷酸受体(Receiver)结构域REC,C-端为具有DNA结合功能的效应(Effector)结构域trans_reg_C(图3)。Cia H可能的胞外感应区未发现保守的结构域。为了解其可能的结构,我们选取该区的168aa进行二级结构预测。结果显示,该区域的两端存在长短不一的α-螺旋结构,中间区域存在5个β-折叠,其余部分为无规卷曲(图4)。根据Cia H同源蛋白序列比对结果(图1)找出S.mutans UA159、S.pneu moniae R6、S.agal actiae 2603 V/R及S.p yogenes M1 GAS在168aa对应区域的序列并进行二级结构预测。有意思的是,虽然这些氨基酸序列相互之间同源性较低,但却具有相似的二级结构(结果未显示)。推测这些菌株的Cia RH可能具有空间结构保守的胞外信号感受结构域。

图3 S.suis 2 Cia RH组分蛋白结构示意图Fig.3 Schematic diagram of S.suis 2 Cia RH component proteins

图4 S.suis 2 Cia H信号感应区(38aa~205aa)二级结构预测Fig.4 Secondary str ucture prediction of Cia H sensor(38aa-205aa)in S.suis 2
2.4 Cia RH在S.suis多种血清型中的分布 考虑到Cia R序列更为保守,我们设计了cia R基因的上下游引物:cia1和cia2,并以不同血清型S.suis的菌液为模板,进行PCR扩增。结果显示,选取的10株2型S.suis菌株中,除了T15菌株,均能扩增出目的条带;30株其他血清型中有17株扩增出目的条带(图5)。

图5 PCR方法检测cia R基因在S.suis多种血清型中的分布Fig.5 Distribution of gene cia R in different serotypes of S.suis by PCR detection 1:S.suis 1/2;2:S.suis 1;3:S.suis 2;4:S.suis 3;5:S.suis 4;6:S.suis 5;7:S.suis 7;8:S.suis 8;9:S.suis 9;10:S.suis 10;11:S.suis 11;12:S.suis 13;13:S.suis 15;14:S.suis 16;15:S.suis 17;16:S.suis 18;17:S.suis 19;18:S.suis 20;19:S.suis 21;20:S.suis 21;21:S.suis 23;22:S.suis 25;23:S.suis 26;24:S.suis 27;25:S.suis 28;26:S.suis 29;27:S.suis 30;28:S.suis 31;29:S.suis 32;30:S.suis 33;31:S.suis 34;32:05ZYH33;33:Negative control;34:266;35:T15;36:98012;37:HA68;38:Habb;39:S735;40:98T003;41:SS2-N;M:1 kb DNA ladder mar ker.
使用NCBI网站BLASTN分别对S.suis不同血清型Cia RH的编码基因cia R和cia H 进行同源序列比对。结果显示,S.suis 2 cia R基因与其他血清型序列一致性为94%~100%,cia H序列一致性为93%~100%。针对cia R的进化关系分析显示,不同血清型来源的序列通常位于不同的分支上,1/2型、14型与2型聚集在一起,而2型菌株GZ1的序列却与D12发生聚集,位于距离05ZYH33最远的分支上(图6)。推测2型S.suis T15菌株未能扩增出目的片段是由于该菌株所含的cia R基因与05ZYH33的cia R基因序列同源性较低。其他13株不同血清型未能扩增出目的条带,可能也是由于相同的原因。综合PCR检测及S.suis不同血清型已知序列分析结果,推测Cia RH在S.suis的不同血清型中广泛存在,不同菌株之间的DNA序列存在不同程度的差异。

图6 S.suis不同血清型cia R基因序列进化树Fig.6 Phylogenetic tree of cia R gene sequences in differentserotypes of S.suis
3 讨 论
TCSTSs是细菌中感受并处理环境信号的主要方式[12]。一般的TCSTSs都包括4个基本的功能模块:位于HKs上的信号感受区(Sensor)及信号传递区(Trans mitter)和位于RRs上的信号接收区(Receiver)及调节/效应区(Regulat or/Eff ector)。信号感受区用于感受特定的环境信号,在此基础上,HKs上的信号传递区保守的组氨酸残基发生ATP依赖的自体磷酸化。这个磷酸基团紧接着被转移到对应的RRs信号接收区保守的天冬氨酸残基上,并使效应区活化,发挥调节作用[13]。
本研究发现在猪链球菌2型05ZYH33 Cia H的N-端存在两个跨膜区,中间存在168aa的间隔区,可能为胞外的信号感受区,但未能找到保守的结构域。通过二级结构预测发现,该区域可能含有一个混合α-螺旋/β-折叠型(又称作PDC)结构域,且可能分布于不同的链球菌属细菌的Cia H中[14]。近期的一个研究结果也显示,这种结构域在不同细菌HKs的胞外感应区广泛分布[15]。由于该类型初级序列的高度多变,普通结构域分析方法无法检出,使其存在被严重低估。因其与PAS结构域在空间结构上的相似性,又被称作PAS样结构域(PAS-like do main)。含有该类型结构域的代表型HKs包括Pho Q、Dcu S及Cit A,它们依次感受环境中的二价离子、C4-双羧酸及柠檬酸成分。针对肺炎链球菌的Cia RH的研究结果显示,该系统可能直接或间接感受Ca2+信号[16]。以上结果提示,S.suis 2 Cia RH可能也通过含有PBC型结构域的胞外信号感受区与环境中的离子或其他小分子成分结合,产生调节效应。
PCR检测及Cia RH编码基因同源序列分析结果提示,该TCSTS在猪链球菌不同血清型中广泛分布,且相互间存在不同程度的序列差异。该系统可能参与S.suis的正常生理及代谢过程,同时,也可能在致病性S.suis感染宿主的特定阶段起到调控作用。我们下一步将针对Cia RH具体感受的信号、调控的靶基因及其在S.suis 2感染宿主不同阶段的作用进行更加深入的研究。
(在序列分析及文章的写作过程中得到中科院微生物研究所程功博士的帮助,特表示感谢!)
[1]Gottschal k M,Xu J,Calzas C,etal.Streptococcus suis:a new emerging or an old neglected zoonotic pathogen[J].Future Micr obiol,2010,5(3):371-391.DOI:10.2217/f mb.10.2
[2]Werthei m HF,Nghia HD,Taylor W,etal.Streptococcus suis:an emerging hu man pat hogen[J].Clin Infect Dis,2009,48:617-625.DOI:10.1086/596763
[3]Tang J,Wang C,Feng Y,etal.Streptococcal toxic shock syndr o me caused by Streptococcus suis serotype 2[J].PLoS Med,2006,3:e151.DOI:10.1371/journal.p med.0030151
[4]Li M,Hu FQ,Tang JQ,etal.Dual signal transduction system and bacterial pat hogenicity[J].J Microbiol,2007,27:50-54.(in Chinese)李明,胡福泉,唐家琪.二元信号转导系统与细菌的致病性[J].微生物学杂志,2007,27:50-54.
[5]Li M,Wang C,Feng Y,etal.Sal K/Sal R,a t wo-co mponent signal transduction system,is essential for f ull vir ulence of highly invasive Streptococcus suis serotype 2[J].PLoS One.2008:3(5):e2080.DOI:10.1371/journal.pone.0002080
[6]Pan X,Ge J,Li M,etal.The or phan response regulator Cov R:a globally negative modulator of virulence in Streptococcus suis serotype 2[J].J Bacteriol,2009,191:2601-2612.DOI:10.1128/JB.01309-08
[7]Feng XD,Cheng G,Wang J,etal.Constr uction and characterization of a t wo-co mponent signal transduction system cia R gene knock-out mutant of Streptococcus suis serotype 2[J].Chin J Zoonoses,2009,25:214-219.(in Chinese)冯晓丹,程功,王晶,等.猪链球菌2型二元信号转导系统cia R基因敲除突变株的构建及生物学功能研究[J].中国人兽共患病学报,2009,25:214-219.
[8]Larkin MA,Blackshields G,Brow NP,etal.Cl ustal W and Cl ustal X version 2.0[J].Bioinfor matics 2007,23:2947-2948.DOI:10.1093/bioinfor matics/bt m404
[9]Tamura K,Peterson D,Peterson N,etal.MEGA5:Molecular evol utionary genetics analysis using maxi mu m likelihood,evol utionary distance,and maxi mu m parsi mony methods[J].Mol Biol Evol,2011,28(10):2731-2739.DOI:10.1093/mol bev/msr121
[10]Petersen TN,Brunak S,Heijne G,etal.Signal P 4.0:discri minating signal peptides from trans membrane regions[J].Nat Met hods,2011,8:785-786.DOI:10.1038/n met h.1701
[11]Marchler-Bauer A,Lu S,Anderson JB,etal.CDD:a conserved domain database for the f unctional annotation of proteins[J].Nucleic Acids Res,2011,39:D225-229.DOI:10.1093/nar/gkq1189
[12]Casino P,Rubio V,Marina A.The mechanism of signal transduction by t wo-component systems[J].Curr Opin Struct Biol,2010,20:763-771.DOI:10.1016/j.sbi.2010.09.010
[13]Stephenson K,Hoch JA.Virulence and antibiotic resistance associated t wo-co mponent signal transduction systems of Grampositive pathogenic bacteria as targets for anti microbial therapy[J].Phar macol Ther,2002,93:293-305.DOI:10.1016/S0163-7258(02)00198-5
[14]Cheung J,Hendrickson WA.Sensor do mains of t wo-co mponent regulatory systems[J].Curr Opin Micr obiol,2010,13:116-123.DOI:10.1016/j.mib.2010.01.016
[15]Chang C,Tesar C,Gu M,etal.Extracytoplas mic PAS-like do mains are co mmon in signal transduction pr oteins[J].J Bacteriol,2010,192:1156-1159.DOI:10.1128/JB.01508-09
[16]Giammarinar o P,Sicard M,Gasc A M.Genetic and physiological st udies of the Cia H-Cia R t wo-co mponent signal-transducing system involved in cefotaxi me resistance and co mpetence of Streptococcus pneu moniae[J].Micr obiology,1999,145(Pt 8):1859-1869.DOI:10.1099/13500872-145-8-1859
