铅锌尾矿区优势草本植物体内铅及氮、磷、钾含量变化特征
2012-08-20张树金李廷轩邹同静熊剑
张树金,李廷轩,邹同静,熊剑
(四川农业大学资源与环境学院,四川 成都611130)
随着工业的发展和农业生产的现代化,环境问题越来越受到人们关注,重金属引起的土壤污染也日益成为环境、土壤科学工作者研究的热点问题。铅(Pb)已成为环境中最普遍的重金属污染物之一,其主要来源于人类活动如采矿、冶炼、焚烧、含铅汽油、颜料及化肥和杀虫剂的施用等[1]。目前,国内外用来降解或消除土壤重金属的基本方法分为物理法、化学法和生物修复法三大类,但前2类方法往往投资昂贵,需用复杂的设备,大多数只能暂时缓解重金属危害,而且有可能造成二次污染[2,3]。近年来,包括微生物、植物和动物3种方法在内的生物修复技术成为土壤污染修复的热点研究领域[4,5]。1983年,美国科学家Chaney首次提出了利用某些能够富集重金属的植物来清除土壤重金属污染的设想——植物修复技术的设想,与传统方法相比,这项技术以其高效、经济和生态协调性等优势显示出巨大的生命力,很快成为国内外研究的热点[6-8]。
国际上报道的超富集植物已有400多种,但其富集重金属的种类还比较有限,其中超富集镍的植物约占73%[9]。但有关植物对铅超富集或富集以及其生物学过程则报道较少。通过对我国植物进行广泛的筛选和调查,首次发现我国一种原生铅富集植物——东南景天(Sedum alfredii),其地上部最高铅浓度可达1 182 mg/kg[10]。但现已发现的超富集植物通常生物量低,生长缓慢,修复效率低,难以形成较完善的植物修复技术以直接大规模应用于修复实践[11]。因此,各国学者就提高超富集植物的生物产量和积累重金属的能力作了大量研究,主要通过采取改良土壤环境、增加土壤肥力等农艺措施提高超富集植物的生长速度、生物产量以及地上部的重金属积累量[12,13]。
氮、磷、钾是植物必需的大量营养元素,在物质组成和代谢过程中有重要的作用,是植物体内蛋白质、核酸等重要物质的合成和代谢过程中不可或缺的组分[14]。植物缺乏大量营养元素的一种或几种均会导致物质组成和代谢的紊乱,进而引起植物品质、生物量和产量的下降。已有研究表明,氮、磷、钾均能显著影响超富集植物对铅的吸收能力[15,16]。但对重金属与大量营养元素分布特点及其交互关系的研究不多,而从植物整个生长时期,研究植物重金属与大量营养元素的相关性甚少[17]。因此,在2007年对三合铅锌尾矿区的调查基础上,从22种草本植物中筛选出9种优势植物,并于2008年对其氮、磷、钾和铅含量进行分析研究。旨在探讨不同生长时期植物铅及氮、磷、钾元素分布情况及其相关性,弄清大量营养元素与铅的交互关系,以期为尾矿区重金属污染的治理和修复提供理论依据。
1 材料与方法
1.1 研究区域概况
研究区位于四川省雅安市荥经县三合乡境内(102°31′E,29°47′N),属中纬度亚热带湿润气候区,年均气温15.3℃,雨量充沛。荥经县矿产资源丰富,各类矿产规模有64处,其中铅锌矿14处,探明贮量共50万t。三合铅锌矿为该地区内具有开采价值的一处,平均含铅7.62%。采样区域为2003年停用的尾矿区,面积约1hm2,海拔1 358~1 445m。区域内土壤类型为黄壤。尾矿区土壤表面被矿渣覆盖,植物以草本植物为主。
1.2 采样方法
2008年在生长前期(春季,5月)、生长旺盛期(夏季,8月)和生长后期(秋季,10月)3个时期进行采样。9种优势植物分别属于7科(表1),每种植物至少采集3个重复,每个重复取6株以上混合样。
1.3 样品分析
采集的植物样品分为地下部和地上部,分别用自来水充分冲洗,去除粘附于植物样品上的泥土和污物,再用去离子水冲洗,在105℃下杀青30min,然后在70℃下于烘箱中烘至恒重,称重。植物样品粉碎后备用。植物样品采用HNO3+HClO4(5∶1,V∶V)在160℃用密闭的聚四氟乙烯罐消煮至澄清。消煮后的溶液洗至100mL容量瓶中,用高纯水定容。样品待测液分别用原子吸收分光光度计测定铅含量[18]。植物样品通过H2SO4+H2O2消煮法消煮,采用蒸馏法测定全氮,钼锑抗比色法测定全磷,火焰光度计法测定全钾[18]。
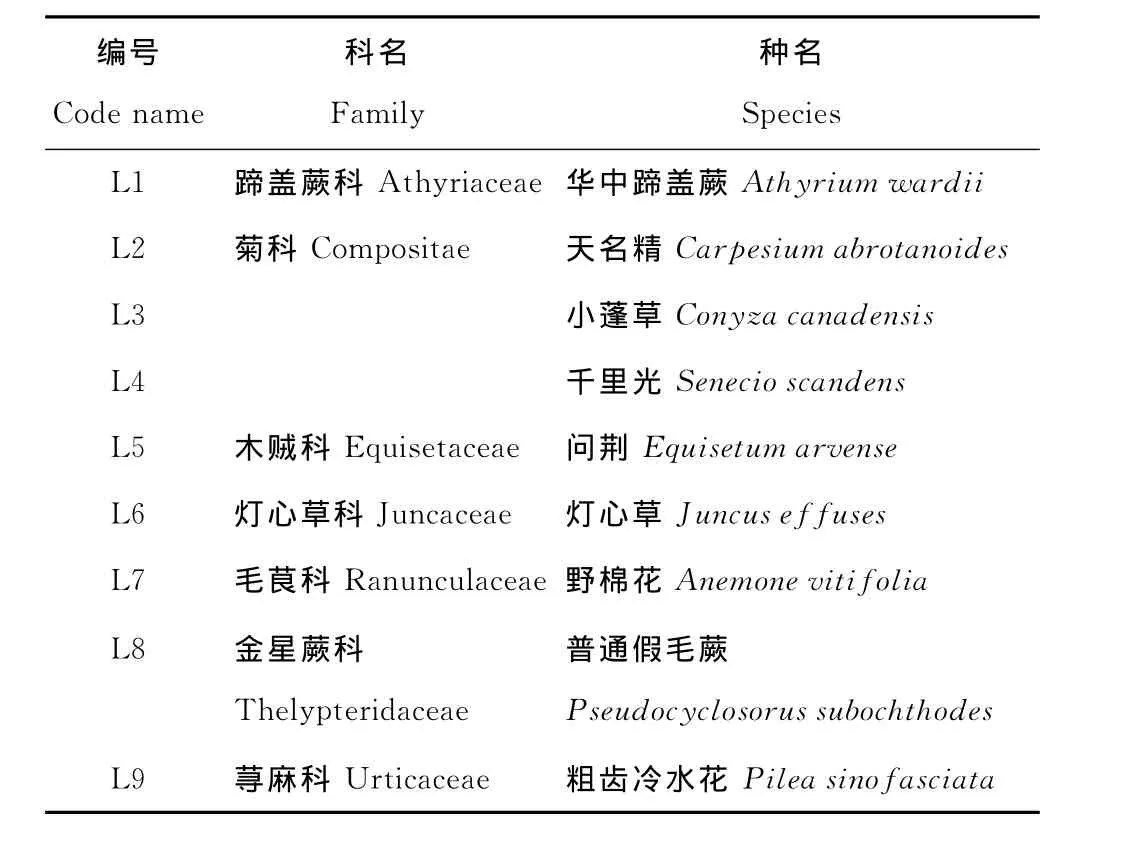
表1 铅锌尾矿区优势草本植物种类Table 1 Family and species composition of the predominant herbaceous plants investigated in the lead-zinc mining tailing
1.4 数据分析
数据用Excel 2003和SPSS 13.0软件进行统计分析。
2 结果与分析
2.1 植物铅含量变化特征
植物吸收铅的能力随着生长环境的变化而变化,影响植物吸收铅的因素是多方面的。在不同生长时期,不同植物铅含量差异很大。植物地下部表现出铅含量高,变异程度大的特点,且明显高于地上部(表2)。在3个生长时期,植株地上部铅含量均小于1 000mg/kg,未达超富集植物的标准。除L1和L8外,其余植物均在生长旺盛期地下部和地上部铅含量同时达到最大值。
在生长前期,L1、L5、L8地下部铅含量超过500mg/kg,与其他植物达到极显著差异。其中L1地下部铅含量为最高,达到15 542.1mg/kg,为其余植物的21.4~251.1倍;地上部仍以L1为最高,达到656.8mg/kg,为其余植物的2.3~16.1倍。在生长旺盛期,多数植物地下部和地上部铅含量均有所增加。地下部铅含量以L1为最高,达到10 720.1mg/kg,为其余植物的25.8~68.8倍;地上部铅含量以L9为最高,达475.4mg/kg。在生长后期,多数植物地下部和地上部铅含量表现出下降的趋势,可能是由于随着生长期的继续延长,其生物量增大,产生稀释效应所致。其地下部和地上部铅含量均以L1为最高,分别达4 132.2和188.0mg/kg,极显著高于其余植物的铅含量。
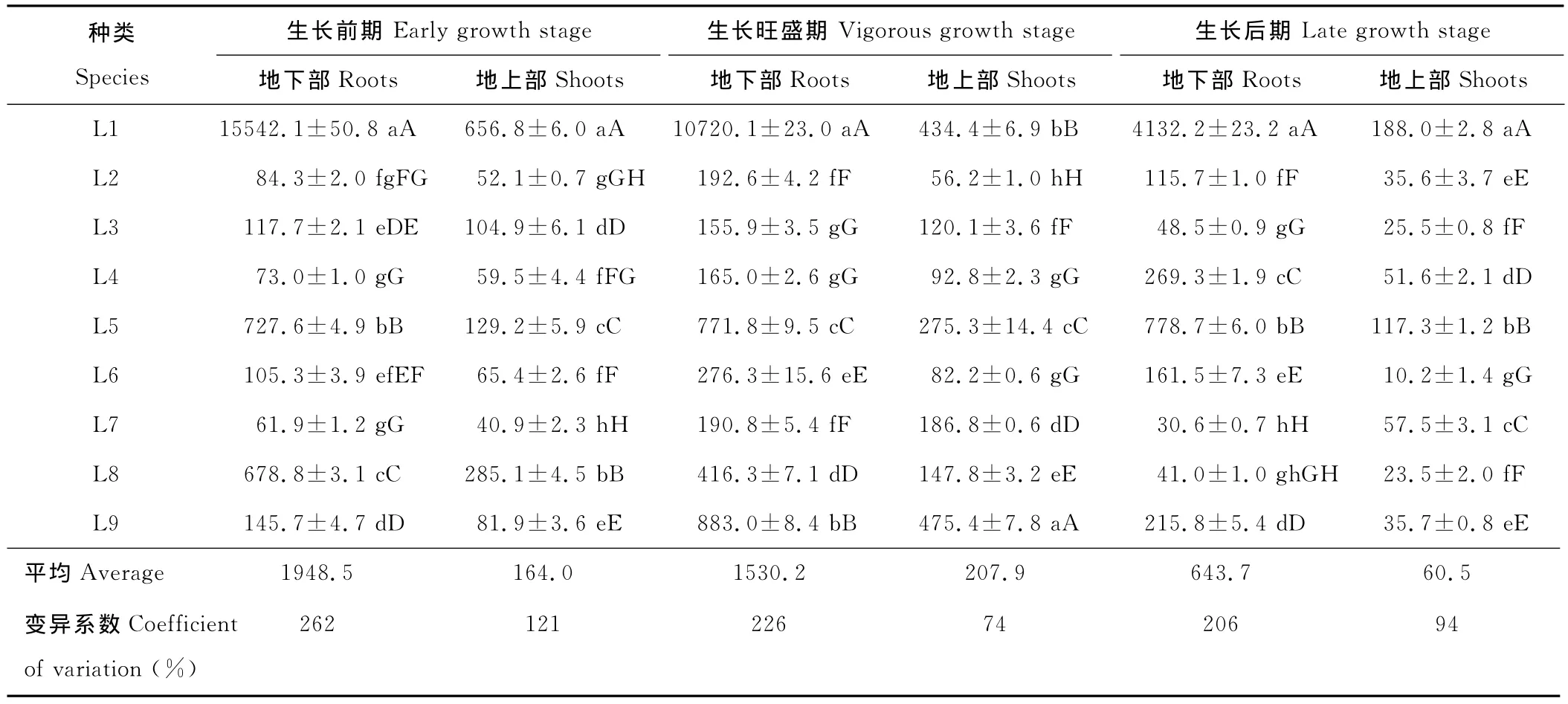
表2 9种优势草本植物3个生长时期铅含量差异Table 2 Lead concentration in the roots and shoots of 9predominant herbaceous plant species at three growing stagesmg/kg
2.2 植物氮含量变化特征
氮被称为生命的元素,它是蛋白质、核酸和磷脂的主要成分,在生命活动中有特殊作用[19,20]。在不同生长时期,不同植物氮含量差异较大(表3)。总体上表现出地上部氮含量高于地下部,在3个生长时期,植物地上部平均氮含量分别为地下部的1.99,2.00和1.74倍。多数植物地下部和地上部氮含量在生长前期达到最大值,随生长时期的延长,氮含量总体上呈现出下降的趋势。在生长前期,地下部氮含量为5.52~10.92g/kg,平均为7.93 g/kg。地上部则表现出氮含量相对较高,变异程度大的特点,平均氮含量为15.82g/kg。在生长旺盛期,多数植物氮含量呈现出降低的趋势,其地下部和地上部平均氮含量分别为5.81和11.62g/kg,与生长前期相比均降低27%。在生长后期,植物氮含量继续下降,地下部氮含量为2.44~6.76g/kg,平均为4.46g/kg;地上部氮含量为5.64~10.79g/kg,平均为7.77g/kg。与生长前期相比,其地下部和地上部平均氮含量分别下降44%和51%。
2.3 植物磷含量变化特征
磷素有“能量元素”之美誉,是植物正常生长发育不可或缺的营养元素之一,在植物整个生命历程中起着极其重要的作用[21]。尾矿区9种优势草本植物磷含量与氮含量的变化趋势相似(表4),在不同生长时期,不同植物磷含量差异较大。在3个生长时期,总体上表现出地上部磷含量高于地下部,其平均磷含量分别为地下部的1.67,1.85和1.28倍。随着生长时期的延长,磷含量呈现下降的趋势。在生长前期,植物地下部磷含量为0.81~1.85 g/kg,平均磷含量为1.28g/kg。地上部则表现出磷含量相对较高,变异程度大的特点,为0.80~3.05g/kg,平均磷含量为2.14g/kg。在生长旺盛期,植物磷含量的变异程度增大,可能是由于在生长旺盛期,不同植物对磷的需求差异较大,导致其选择吸收和积累的量有差别所致。但其平均磷含量仍略有降低,与生长前期相比,磷含量分别降低16%和7%。在生长后期,植物地下部和地上部平均磷含量分别为1.02和1.31g/kg,与生长前期相比,分别降低20%和39%。
2.4 植物钾含量变化特征
钾在植物的生长发育以及产量形成过程中都有十分重要的作用。钾并不直接参与植物体内组织、器官的形成,而是通过促进植物体内生理、生化活动及代谢反应来调节植物的生长发育及产量形成[22]。在不同生长时期,不同植物钾含量差异较大,总体上表现出地下部钾含量低于地上部(表5)。植物地下部和地上部钾含量均在生长前期或生长旺盛期时达到最大值,随着生长时期的继续延长,钾含量呈现出下降的趋势,并在生长后期达到最小值。在生长前期,植物地下部钾含量为2.78~16.04g/kg,平均为10.81g/kg;地上部钾含量为10.63~32.59 g/kg,平均为19.89g/kg,是地下部平均钾含量的1.84倍。在生长旺盛期,植物地下部和地上部平均钾含量分别为10.75和18.95g/kg,后者为前者的1.76倍。在生长后期,植物钾含量处于较低水平,地下部和地上部平均钾含量分别为8.42和13.96g/kg,与生长前期相比,分别下降22%和30%。

表3 9种优势草本植物不同生长时期氮含量差异Table 3 Nitrogen concentration in the roots and shoots of 9predominant herbaceous plant species at three growing stagesg/kg

表4 9种优势草本植物3个生长时期磷含量差异Table 4 Phosphorus concentration in the roots and shoots of 9predominant herbaceous plant species at three growing stagesg/kg

表5 9种优势草本植物3个生长时期钾含量差异Table 5 Potassium concentration in the roots and shoots of 9predominant plant herbaceous species at three growing stagesg/kg
3 讨论
3.1 植物铅与氮、磷、钾含量变化特征
李裕红等[17]在对泉州湾海岸植物研究指出,钾为含量最高的大量营养元素,高含量的钾有助于增强植物的抗逆性。自然环境下生长的泉州湾海岸植物的氮、磷、钾含量与其铜、锌、铅含量之间无显著相关关系。本研究得出,钾含量最高,氮次之,磷最低。尾矿区9种优势草本植物总体上表现出地上部氮、磷、钾含量高于地下部,并随着生长时期的延长,呈现出下降的趋势。这可能是由于随着生长时期的延长,植物发生木质化,并主要以糖类、纤维素等碳水化合物的形式增加生物量,导致氮、磷、钾的含量相对降低。植物铅含量则是地上部小于地下部,可能是由于铅为植物非必需营养元素,凯氏带的阻隔将大量的铅滞留在地下部,这也是至今尚未发现真正意义上的铅超富集植物的根本原因所在[23]。
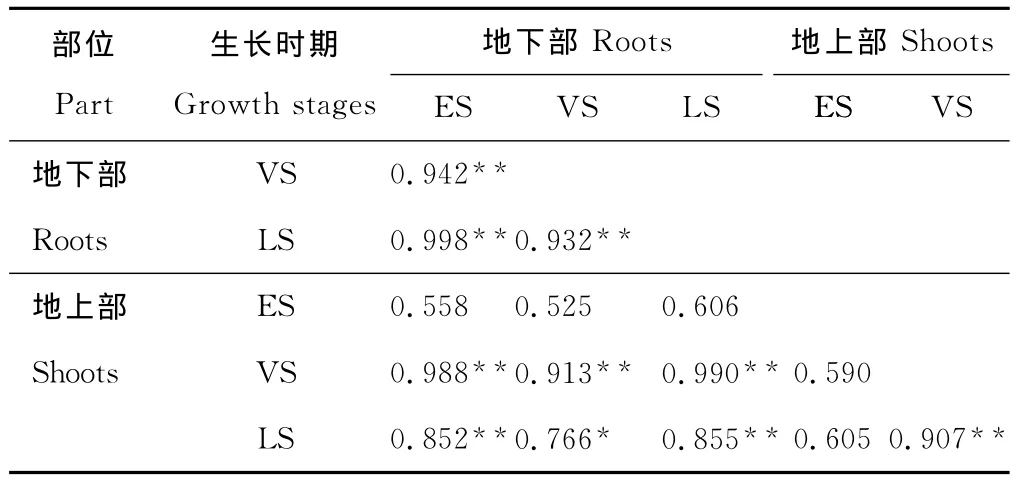
表6 9种优势草本植物地上部和地下部铅含量在3个生长时期的相关性分析Table 6 Relationship between lead concentration of root and shoot of 9predominant herbaceous plant species at three growth stages
相关性分析得出,在植物生长前期,其地上部铅含量与地下部铅含量以及与生长旺盛期和生长后期的地上部或地下部铅含量均不存在显著的相关关系,而在生长旺盛期和生长后期,同一时期地上部和地下部铅含量之间以及不同时期地上部与地下部铅含量之间均存在显著或极显著的相关关系(表6)。这是因为植物主要通过根系吸收重金属铅,并从地下部往地上部运输,但铅在植物体内移动性小,不易转移。由于不同的植物生长速度和转移能力不同[24],导致在生长前期,9种优势草本植物地上部吸收和累积铅的能力存在明显差异,故无显著的相关关系。因此,在尾矿区筛选铅植物修复材料时,应避开在植物生长前期进行采样。但不同类型的矿区及气候条件对植物重金属的吸收累积存在差异,筛选时期可能存在一定的差异。
3.2 大量营养元素在植物修复中的作用
现已发现的超富集植物通常生物量低,生长缓慢,修复效率低,阻碍了植物修复技术以直接大规模应用于修复实践[25]。因此,前人的研究致力于通过施加外源物质等来 提 高 修 复 效 果[26]。 李 继 光 等[27,28]研 究 表明,在外界生长环境的氮素含量未达到东南景天的最适临界点情况下,适当施加氮肥显著促进东南景天的生长,提高其干物质产量,同时促进了东南景天对镉的吸收及向地上部分的转运。孙琴等[12]研究得出,适当增磷显著促进东南景天的生长,提高其干物质产量,同时促进了东南景天对锌的吸收及锌向地上部分的运转和积累。聂俊华等[16]在对单一营养元素氮、磷、钾对植物富集铅的研究中得出,少量的氮和钾可促进富集植物干质量的增加,促进植物对铅的吸收,随着氮和钾水平的增加,植物对铅的吸收能力降低,但钾的抑制作用不如氮的显著;土壤供磷会降低植物对铅的吸收,且铅含量下降达极显著水平。相关性分析得出,9种优势草本植物地下部氮、磷、钾含量与铅含量均不存在显著的相关关系(表7),这可能是由于植物吸收氮、磷、钾的方式与吸收铅的方式不同及铅在植物体内的移动性相对较小所致。在生长后期,9种优势植物地上部氮和钾含量与铅含量存在显著的正相关关系(0.750*和0.773*),表明氮和钾可能对解重金属铅的毒害起到一定的作用。
华中蹄盖蕨(L1)是一种常见的多年生蕨类植物,这种蕨类植物根系发达、根部铅吸收能力强、转运能力弱,是一种新型的铅稳定修复材料,具有潜在的应用价值[29]。本研究得出,华中蹄盖蕨能够将大量铅囤积在地下部,在3个生长时期,其根部累积量分别为32.64,41.81和21.49mg/株,并维持正常的生长,这可能与其在3个生长时期地上部、地下部氮含量均处于较高水平(表2),大量蛋白质与重金属结合降低其毒性有关。然而,能否通过增施氮肥,提高华中蹄盖蕨的稳定修复效率仍需进一步的研究。
4 结论
尾矿区9种优势草本植物以钾为含量最高的大量营养元素,氮次之,磷最低,且植物地上部氮、磷、钾含量均高于地下部,并随着生长时期的延长呈下降趋势。植物铅含量差异很大,在不同生长时期其含量变化也较明显,表现出地下部明显高于地上部。9种优势草本植物均未表现超富集铅的能力,但华中蹄盖蕨地下部能够囤积大量铅,并保持正常生长,进一步证实其能作为稳定修复的先锋植物,应进一步研究探讨其富集忍耐重金属的机理。
尾矿区9种优势草本植物地下部氮、磷、钾含量在3个生长时期均与铅含量无显著的相关关系。在生长后期,地上部氮、钾含量与铅含量存在显著的正相关关系,表明氮和钾可能对植物解重金属铅的毒害起到一定的作用,其机理目前尚不清楚,还有待进一步研究。
在植物生长前期,其地上部铅含量与地下部铅含量以及与生长旺盛期和生长后期的地上部或地下部铅含量均不存在显著的相关关系,而在生长旺盛期和生长后期,同一时期地上部和地下部铅含量之间以及不同时期地上部与地下部铅含量之间均存在显著或极显著的相关关系。因此,在尾矿区筛选铅植物修复材料时,应避开在植物生长前期采样。但不同类型的矿区及气候条件对植物重金属的吸收累积存在差异,筛选时期可能存在一定的差异。
[1] 王云,魏复盛.土壤环境元素化学[M].北京:中国环境科学出版社,1995.
[2] Gworek B.Lead inactivation in soils by zeolites[J].Plant and Soil,1992,143(1):71-74.
[3] Vangronsveld J,Colpaert J V,Van Tichelen K K.Reclamation of a bare industrial area contaminated by non-ferrous metals:Physico-chemical and biological evaluation of the durability of soil treatment and revegetation[J].Environmental Pollution,1996,94(2):131-140.
[4] 龙新宪,杨肖娥,倪吾钟.重金属污染土壤修复技术研究的现状与展望[J].应用生态学报,2002,13(6):757-762.
[5] 丁佳红,刘登义,储玲,等.重金属污染土壤植物修复的研究进展和应用前景[J].生物学杂志,2004,21(4):6-9,20.
[6] Chaney R L,Malik M,Li Y M,et al.Phytoremediation of soil metals[J].Current Opinion in Biotechnology,1997,8(3):279-284.
[7] 李源,李金娟,魏小红.镉胁迫下蚕豆幼苗抗氧化能力对外源NO和H2O2的响应[J].草业学报,2009,18(6):186-191.
[8] 李影,王友保.4种蕨类草本植物对Cu的吸收和耐性研究[J].草业学报,2010,19(3):191-197.
[9] González R C,González-Chávez M C A.Metal accumulation in wild plants surrounding mining wastes[J].Environmental Pollution,2006,144(1):84-92.
[10] He B,Yang X E,Ni W Z,et al.Sedum alfredii:A new lead-accumulating ecotype[J].Acta Botanica Sinica,2002,44(11):1365-1370.
[11] Lelie D V D,Schwitzguébel J P,Glass D J,et al.Peer Reviewed:Assessing phytoremediation’s progress in the United States and Europe[J].Environmental Science & Technology,2001,35(21):446-452.
[12] 孙琴,倪吾钟,杨肖娥,等.磷对超积累植物——东南景天生长和积累锌的影响[J].环境科学学报,2003,23(6):818-824.
[13] 李新博,谢建治,李博文,等.镉对紫花苜蓿不同生长期生物量的影响及饲用安全评价[J].草业学报,2009,18(5):266-269.
[14] 安志装,王校常,施卫明,等.重金属与营养元素交互作用的植物生理效应[J].土壤与环境,2002,11(4):392-396.
[15] 聂俊华,刘秀梅,王庆仁.Pb超富集植物对营养元素N、P、K的响应[J].生态环境,2004,13(3):306-309.
[16] 聂俊华,刘秀梅,王庆仁.营养元素N、P、K对Pb超富集植物吸收能力的影响[J].农业工程学报,2004,20(5):262-265.
[17] 李裕红,袁建军,陈怀宇,等.泉州湾若干海岸植物铜、锌、铅及氮、磷、钾元素分布特征[J].中国生态农业学报,2008,16(3):630-634.
[18] 鲁如坤.土壤农业化学分析方法[M].北京:中国农业科技出版社,2000.
[19] 王忠.植物生理学[M].北京:中国农业出版社,2000.
[20] 赵平,孙谷畴,彭少麟.植物氮素营养的生理生态学研究[J].生态科学,1998,17(2):37-42.
[21] 陆景凌.植物营养学(上册)[M].北京:中国农业大学出版社,2003.
[22] 曲婷婷.不同低钾耐性大豆叶片显微结构及光合特性差异[D].沈阳:沈阳农业大学,2009.
[23] Meyers D E R,Auchterlonie G J,Webb R I,et al.Uptake and localisation of lead in the root system of Brassica juncea[J].Environmental Pollution,2008,153(2):323-332.
[24] Huang H,Li T,Tian S,et al.Role of EDTA in alleviating lead toxicity in accumulator species of Sedum alfredii H.[J].Bioresource Technology,2008,99(14):6088-6096.
[25] 杨肖娥,龙新宪,倪吾钟,等.古老铅锌矿山生态型东南景天对锌耐性及超积累特性的研究[J].植物生态学报,2001,25(6):665-672.
[26] 傅华,王玉梅,周志宇,等.施用污泥对黑麦草草坪绿地土壤理化性质和重金属元素含量的影响[J].草业学报,2003,12(2):82-86.
[27] 李继光,朱恩,李廷强,等.氮对镉胁迫下东南景天根系形态及镉积累的影响[J].环境污染与防治,2007,29(4):271-275.
[28] 李继光,李廷强,朱恩,等.氮对超积累植物东南景天生长和镉积累的影响[J].水土保持学报,2007,21(1):54-58.
[29] Zou T,Li T,Zhang X,et al.Lead accumulation and tolerance characteristics of Athyrium wardii (Hook.)as a potential phytostabilizer[J].Journal of Hazardous Materials,2011,186(1):683-689.
