新疆野生黄花苜蓿群体果实的形态变异
2012-03-13王玉祥
王玉祥,张 博
(新疆农业大学草地资源与生态重点实验室,新疆 乌鲁木齐 830052)
种子是植物进行有性生殖最重要的器官之一,研究种子的大小,可以了解种子的生物学功能,且种子大小与干扰、种子产量、成苗、植被动态等有密切关系。种子大小在种群内、种群间,甚至个体间,都有很大差异[1-3],可以作为种子形态变异的重要内容,种子大小的研究是掌握种子表型变异规律及开展遗传育种研究的基础。
对种子表型性状的研究结论均证明了种子与环境之间有密切的关系:在大格局上种子随纬度或海拔的变化,其表型性状呈现出一定梯度性规律变化[4-6];从微环境上看,土壤养分、土壤含水量等因素亦影响着种子的形态在种群内和种群间的变异[7-10]。本研究就黄花苜蓿(Medicagofalcata)种子表型及质量变异方面进行研究,一方面是为了弄清其荚果的变异特征和主要影响因素,另一方面是为牧草改良和育种等研究提供科学依据。同时,开展荚果表型变异研究是黄花苜蓿遗传多样性研究的一部分,有利于了解苜蓿的遗传多样性背景,为苜蓿质资源评价、保存和利用提供依据。
1 材料与方法
1.1试验材料 试验材料由新疆草地资源与生态实验室提供,2007-2011年采自新疆各地(表1)。
1.2试验方法 每个群体随机选取10个单株,每个单株随机选取30个荚果和30粒种子,用游标卡尺(±0.02 mm)测量荚果长、荚果宽、每荚种子数、粒长和粒宽等指标。
1.3数据分析 利用DPS 6.5软件对试验数据进行分析,分析各个性状的变异程度和群体间显著性,对形态的各个指标标准化后用算术平均法(UPGMA)对各群体进行聚类分析。
2 结果与分析
2.1黄花苜蓿果实性状的差异分析 13个群体的黄花苜蓿的荚果颜色为褐色,形状多为镰刀状。种子土黄色,形状为肾形、圆形(图1)。
13个群体中,荚果最长的是北屯(1.25 mm), 最短的是额敏(0.79 mm),相差0.46 mm;荚果最宽的是哈巴河(0.28 mm),最窄的是察布查尔(0.17 mm),相差0.07 mm;每荚含种子数最多的是北屯(7.8粒),最少的是裕民(2.8粒),相差5粒;种子粒长最长的是乌市南山(0.25 mm),最短的是吉木乃(0.18 mm),相差0.07 mm;粒宽最宽的是阿勒泰(0.16 mm),最窄的是乌市南山(0.11 mm),相差0.05 mm;千粒重最大的为布尔津(1 715.5 mg),最小的是哈巴河(1 062.0 mg),相差653.5 mg(表2)。以上结果表明,不同区域的黄花苜蓿在荚果和种子的形态特征方面存在较大差异。

表1 黄花苜蓿信息Table1 Information of Medicago falcata

图1 黄花苜蓿荚果和种子形态特征Fig.1 Morphological characteristics of pod and seed of Medicago falcata
变异系数能够反映表型性状的变异程度。同一群体内部,不同表型性状的变异系数不同(表2)。在福海群体中,荚果宽变异系数为31.91%,千粒重变异系数为0.10%。同一性状的变异系数值在不同群体中存在一定程度的变化,荚果长最大变异系数为裕民(48.02%),最小为北屯(0.67%)。方差分析表明(表3),在荚果和种子的调查性状中,各指标在群体间都呈现显著分化(P<0.05),其中荚果长和千粒重呈现极显著分化(P<0.01),这可能与其生境有关系。在总变异中,各性状种群内最小的是每荚果种子数(71.11%),最大的是粒宽(89.42%)。群体间变异最大的是每荚果种子数(28.89%),最小的是粒宽(10.58%)。群体内平均变异组占84.51%,群体间平均变异组占15.50%,这说明苜蓿表型变异主要来自群体内,但群体间也出现了一定的分化。
2.2相关性分析 荚果及种子的特征与地理因子之间无相关性(P>0.05);荚果长、每荚种子数和粒长与千粒重呈极显著正相关(P<0.01),粒宽与千粒重呈显著正相关(P<0.05),这表明荚果越长、越宽,则种子相应地变长、变宽、质量增加。荚果长与每荚种子数之间呈极显著正相关,荚果宽与每荚种子数之间呈正相关关系,即荚果越长、越宽,种子数越多。粒长与粒宽有极显著正相关(表4)。
2.3聚类分析 为了进一步探讨各个群体间的亲缘关系,采用最短距离法对13个黄花苜蓿群体进行聚类分析,在欧氏距离2.18处将13个群体聚为3类:其中,阿勒泰群体单独聚为一类,该区地处山坳中间,海拔较低,气候温暖湿润,植株低矮,荚果扁平,种子圆滑,形状接近圆形,可划为圆籽粒型;北屯、塔城和富蕴群体聚为一类,虽然这3个群落相距较远,但样品具有共同特征,籽粒较小,形状不规则,似多边形,可划为多边籽粒型;布尔津、裕民、哈巴河、吉木乃、额敏、察布查尔、特克斯、乌市南山和福海群体聚为一类,每荚种子粒数相对较多,种子形状接近肾形,可划为肾形籽粒型(图2)。这表明,在居群水平上,形态分化与居群间的地理距离远近不一定成正比,各个群体间地理距离虽然相距很远,但它们群体之间的形态差异不一定大[11-12],这与植物的生境、传播及演化路径有一定的关系。
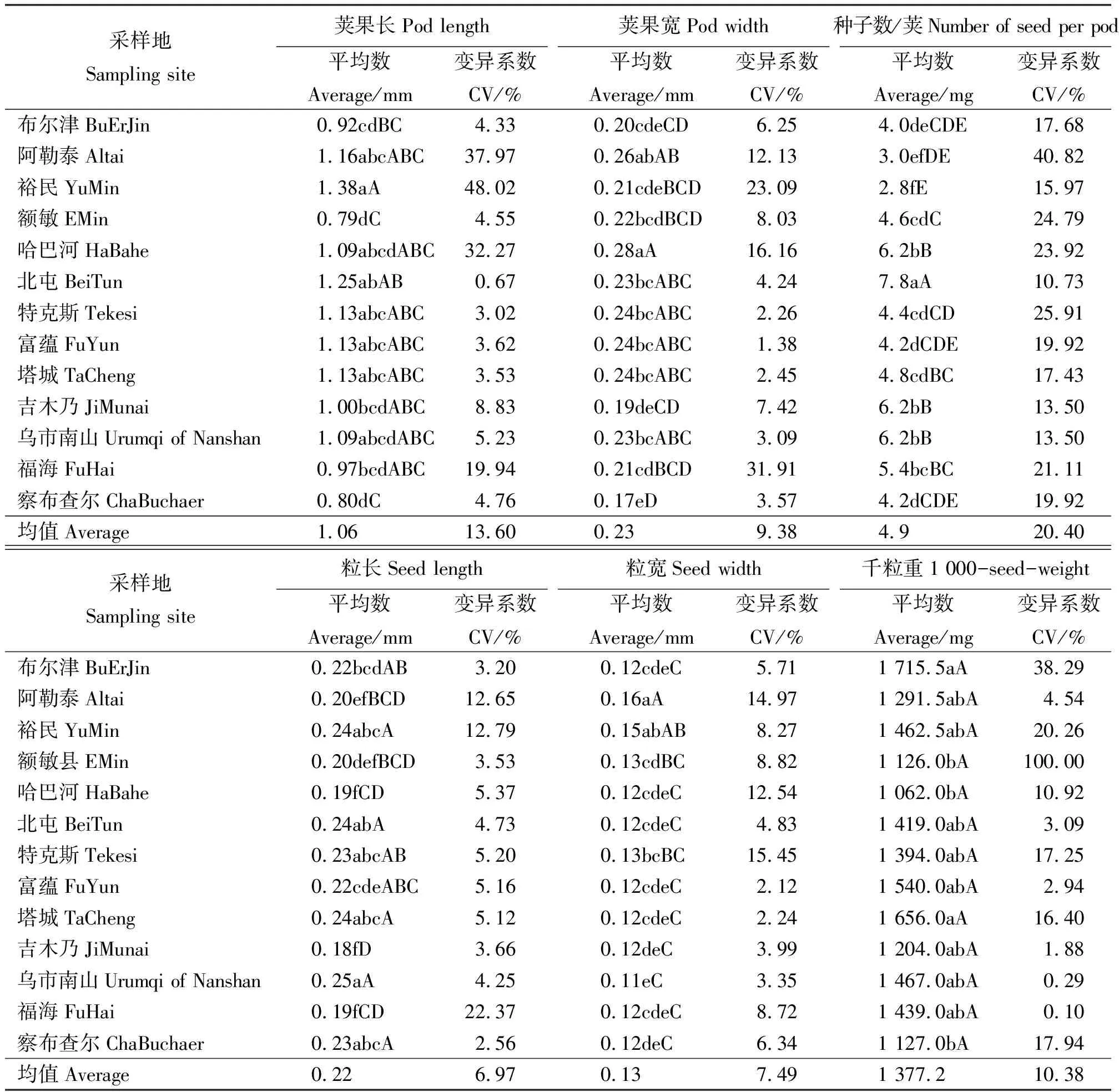
表2 黄花苜蓿荚果和种子的表型特征 Table 2 Phenotypic characteristics of pod and seed of Medicago falcata
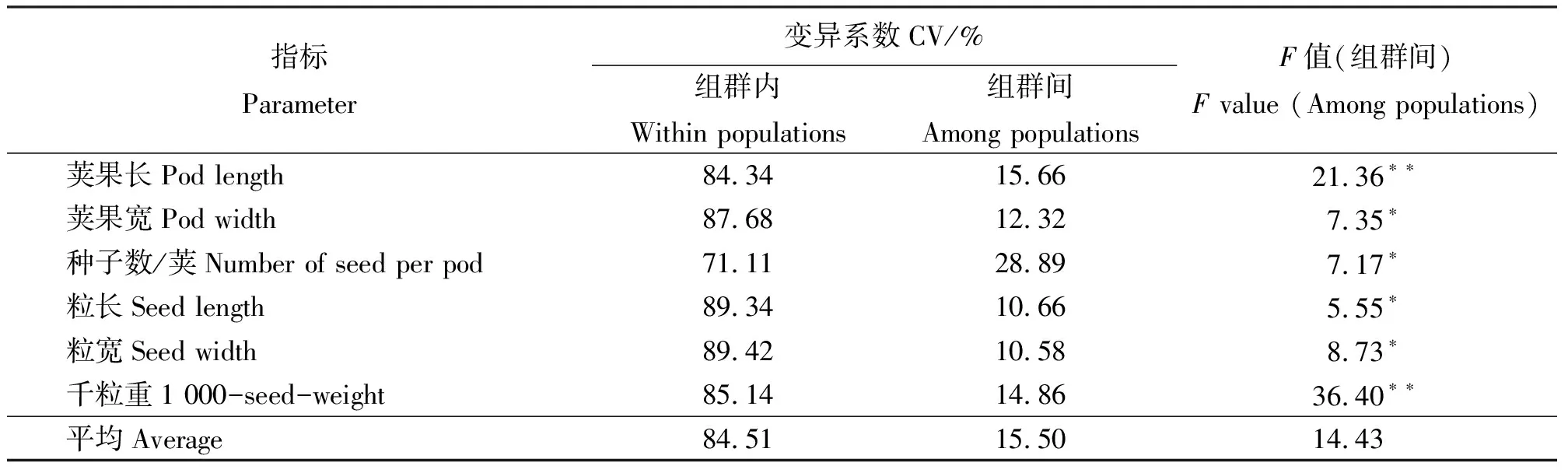
表3 荚果和种子性状的变异分析Table 3 Variance distribution between pods and seed traits
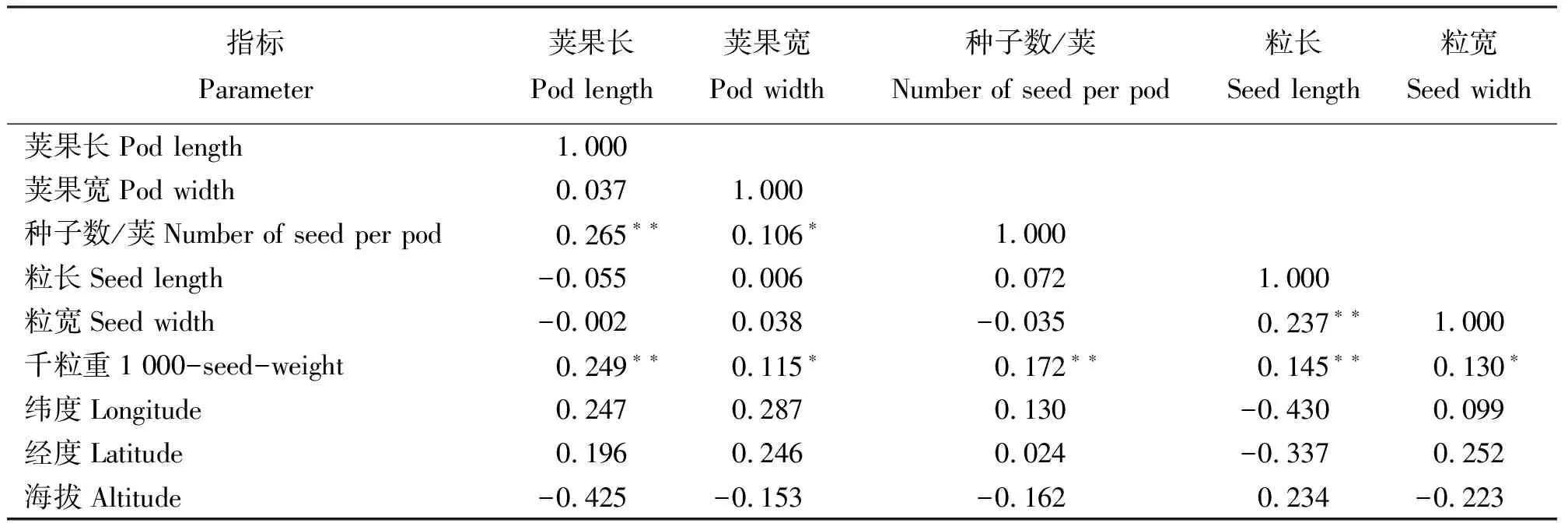
表4 荚果和种子性状与地理位置之间的相关分析Table 4 Correlation coefficient among morphological trait and between morphological traits and geological factors
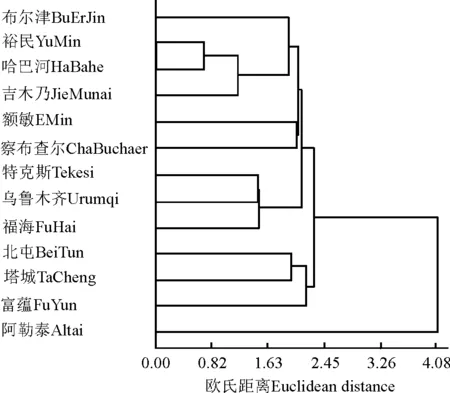
图2 聚类分析Fig.2 Cluster Ananlysis
3 讨论与小结
本研究对13个群体的苜蓿荚果及种子特性进行分析得出,在群体间各性状均有显著或极显著差异。其中,荚果长、荚果宽、每荚种子数、粒长、粒宽和千粒重在各群体之间的差异均达到极显著水平。这进一步说明了不同生境条件下的黄花苜蓿群体经过长期的自然选择和适应,荚果和种子形态特征在居群间已产生明显的分化,这与于林清等[11]的研究结果一致 ,黄花苜蓿野生种群拥有较丰富的遗传多样性,在种群内和种群间存在着丰富的遗传变异。
植物性状变异是环境因素和遗传因素共同作用的结果,地理阻隔和不同生境条件的长期作用可使植物群体间产生较大的差异,这是植物对生存环境的一种本能反应,是长期演化的结果[12-14]。植物的形态特征具有一定的稳定性和变异性,受本身的遗传组成和所处环境两方面的影响。尽管形态变异具有一定的遗传基础,但是环境压力在植物形态变异中也起着重要作用。植物很难处于生长发育所需的最适宜环境条件,无论是气候变化还是生长发育过程的群落条件,在其生长过程中各个植株之间体总要产生或大或小的差异[13-15]。
植物在生长发育过程中,由于地理位置的差异和生境的多样化,不同群体间以及种内不同居群为适应生态环境,均表现出丰富的形态多样性。育种者希望材料间遗传差异大和多样性丰富,这样有利于优质育种材料的选择[11-12,16-17]。本研究表明,黄花苜蓿不同居群适应环境能力强,其性状在种群内和种群间均存在丰富的变异,是较好的育种资源。
[1] 管俊娇,虞泓,何露,等.小桐子居群种子表型变异研究[J].西南农业学报,2011,24(3):10086-10089.
[2] 赵志刚,郭俊杰,沙二,等.我国格木的地理分布与种实表型变异[J].植物学报,2009,44(3):338-344.
[3] 谢春平,方彦,方炎明.福建茫荡山乌冈栎种子性状变异研究[J].安徽农业大学学报,2011,38(5):656-661.
[4] 刘桂丰,杨传平,刘关君,等.白桦不同种源种子形态特征及发芽率[J].东北林业大学学报,1999,27(4):1-4.
[5] 王小平,刘晶岚,王九龄,等.白皮松种子及球果形态特征的地理变异[J].北京林业大学学报,1998,20(3):25-31.
[6] 蔡永立,王希华,宋永昌.中国东部亚热带青冈果实形态变异的研究[J].生态学报,1999,19(4):81-86.
[7] 窦全丽,何平,肖宜安,等.濒危植物缙云卫矛果实、种子形态分化研究[J].广西植物,2005,25(3):219-225.
[8] 孙玉玲,李庆梅,杨敬元,等.秦岭冷杉球果与种子的形态变异[J].生态学报,2005,25(1):176-181.
[9] 赵志刚,郭俊杰,沙二,等.我国格木的地理分布与种实表型变异[J].植物学报,2009,44(3):338-344.
[10] 佘诚棋,方升佐,杨万霞.青钱柳种子形态特征的地理变异[J].南京林业大学学报:自然科学版,2008,32(4):63-66.
[11] 于林清,王照兰,萨仁,等.黄花苜蓿野生种群遗传多样性的初步研究[J].中国草地,2001,23(1):23-25.
[12] 李世雄,王彦荣,孙建华.中国苜蓿品种种子产量性状的遗传多样性[J].草业学报,2003,12(1):23-29.
[13] 德英,穆怀彬,刘新亮,等.披碱草属8 种野生牧草居群穗部形态多样性[J].草业科学,2011,28(9):1623-1631.
[14] 严学兵,郭玉霞,周禾,等.青藏高原垂穗披碱草变异的地理因素分析[J].西北植物学报,2007,27(2):328-333.
[15] 王金龙,高玉葆,赵念席,等.内蒙古中东部草原克氏针茅形态特征和RAPD遗传分析的相关性分析[J].植物研究,2006,26(6):709-714.
[16] 杨允菲,李建东.东北羊草草原种群单穗数量性状的生态可塑性[J].生态学报,2011,21(5):753-758.
[17] 陈学林,杨建美,王文敏.青藏高原东缘常见40种禾本科植物种子大小变异研究[J].植物研究,2011,31(1):9-14.
