干旱胁迫对玉米耐旱自交系与旱敏感自交系苗期生理特性的影响
2012-03-13彭云玲张为龙方子森
彭云玲,王 涛,李 燕,张为龙,方子森
(甘肃省干旱生境作物学重点实验室 甘肃省作物遗传改良与种质创新重点实验室 甘肃农业大学农学院,甘肃 兰州 730070)
玉米(Zeamays)是世界上主要的粮食和饲料作物之一,而干旱是影响我国及世界玉米稳产的重要环境因子。我国2/3的玉米种植面积分布在东北、华北、西南和西北依靠自然降水的丘陵旱地或平原旱地上。玉米耐旱性已愈来愈引起人们的重视。目前,在玉米耐旱指标方面已经作了大量工作[1-3],斐英杰等[4]的研究表明,用幼苗耐旱性鉴定结果可以评价玉米自交系的耐旱性。同一植物不同品种之间由于基因型的差异其抗旱能力存在差异[5],周树峰[6]鉴定了57份玉米自交系的耐旱性,认为苗期耐旱性与成株期的耐旱性显著相关。刘贤德等[7]采用叶面积指数、叶片相对含水量和耐旱系数等指标在苗期评价了9份耐旱系和9份旱敏感系的性状差异。本试验以2010年在54份玉米自交系中初步筛选出耐旱性有差异的自交系廊黄、昌7-2和TS141为材料,探讨玉米耐旱系与旱敏感系的耐旱性差异,以期揭示耐旱玉米基因型适应干旱的机理,从而为玉米种质改良和最大限度地挖掘玉米自身的生物学潜力提供材料,为大幅度地提高玉米水分利用效率和产量提供理论依据。
1 材料与方法
1.1试验材料 玉米耐旱自交系廊黄、昌7-2和旱敏感自交系TS141由甘肃省农业科学院提供。其中廊黄来源于四平头系,昌7-2来源于(黄早4×潍95)×S901的后代,TS141引自临洮农业学校,亲缘不详。
1.2材料培养与干旱处理 2011年7月在温室中将3个玉米自交系播种在高15 cm、内径9 cm的塑料花盆中,栽培基质由1∶1的花卉土、蛭石构成,每盆10粒,播种前统一配土、装钵、浇透底水。试验设对照组和干旱胁迫组,每个品种每个处理3个重复。播种后5 d出苗,对照组每钵每隔5 d浇水50 mL。干旱胁迫组在出苗后每钵浇50 mL水,而后停止浇水。2周后旱敏感自交系基部叶片开始萎蔫,以此为标准,5 d后观察3个自交系植株的叶片萎蔫和卷曲程度,7 d后分别测定植株的各个生理指标。
1.3幼苗干旱胁迫后的存活率 将供试玉米种子播于宽20 cm、高15 cm、长40 cm的盆钵中,在栽培基质中生长2周后进行干旱胁迫,控水处理至玉米叶片永久萎蔫后,复水1周,统计存活率,试验设3次重复。
1.4测定指标与方法 超氧化物歧化酶(SOD)和过氧化物酶(POD)活性测定参照邹琦[8]的方法,叶片组织含水量、脯氨酸(Pro)含量和质膜相对透性(相对电导率)的测定参照李合生[9]的方法。
1.5数据处理 所有数据采用Microsoft Office Excel 2003软件进行统计分析,通过学生t检验对正常浇水和干旱胁迫条件下耐旱自交系廊黄、昌7-2与旱敏感自交系TS141的耐旱性分别进行比较。
2 结果与分析
2.1干旱胁迫下玉米自交系的形态特征和存活率 在水分供应充足的情况下,3个自交系的植株生长正常且一致。轻度干旱胁迫时,旱敏感自交系TS141植株的基部叶片边缘开始向内卷曲,而耐旱自交系廊黄和昌7-2植株的叶子边缘没有出现卷曲现象。干旱胁迫处理5 d后,所有旱敏感自交系TS141植株的成熟叶片出现萎蔫,最上部的幼嫩叶片也出现严重的卷曲,基部叶片变黄。而耐旱系廊黄植株叶片仍然保持绿色,昌7-2叶片出现略微的萎蔫,但是叶片仍然保持绿色,基部叶片没有变黄(图1)。

图1 干旱胁迫下玉米耐旱自交系与旱敏感自交系幼苗的生长状况Fig.1 Seediling growth of drought-tolerant and -sensitive maize lines under drought stress
在水分供应充足的条件下,3个自交系的存活率都是100%。玉米幼苗经干旱处理后,3个玉米自交系的存活率均下降,耐旱自交系廊黄和昌7-2的存活率分别是88%和75%,而旱敏感自交系TS141的存活率仅为33%,耐旱自交系的存活率显著高于与旱敏感自交系的存活率(P<0.05)(图2)。
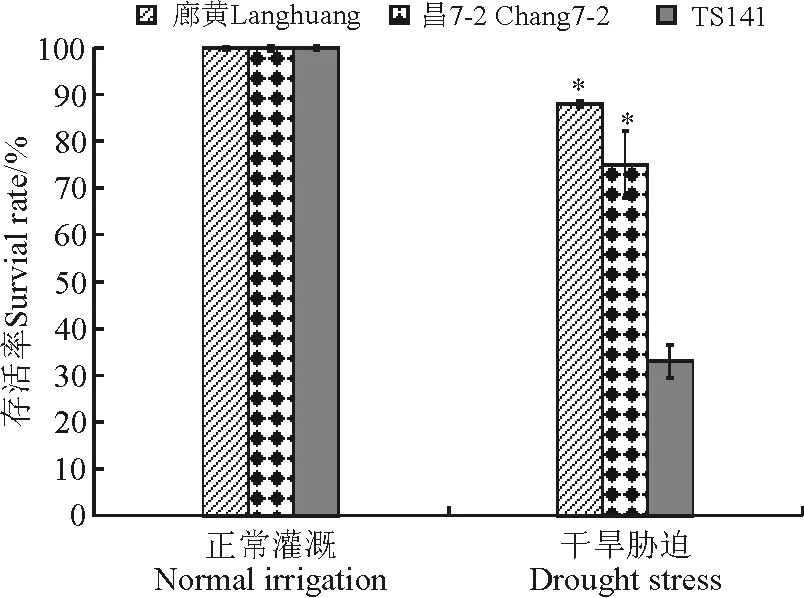
图2 干旱胁迫下玉米耐旱自交系与旱敏感自交系的幼苗存活率 Fig.2 Survival rates of seedlings of drought-tolerant and -sensitive maize lines under drought stress
2.2干旱胁迫对玉米自交系生理指标的影响
2.2.1相对含水量 水分充足条件下,3个玉米自交系的相对含水量在94%左右。幼苗遭受干旱胁迫后,耐旱系和旱敏感系的相对含水量都下降,耐旱系廊黄和昌7-2分别为89%和90%,而旱敏感自交系的相对含水量为37%。显著性差异分析表明耐旱自交系廊黄、昌7-2与TS141相对含水量的差异达到极显著水平(P<0.01)(图3),说明耐旱自交系的相对含水量下降幅度小,而旱敏感自交系相对含水量的下降幅度大。
2.2.2叶片质膜透性 在水分充足条件下,耐旱自交系廊黄、昌7-2和旱敏感自交系都有较低的相对电导率,分别为11.45%、11.14%和12.56%。当植株遭到干旱胁迫后,3个自交系的相对电导率明显上升,耐旱自交系廊黄和昌7-2的相对电导率为28%和44%,旱敏感自交系的为57%,且耐旱自交系廊黄、昌7-2与旱敏感系TS141相对电导率的差异达到显著水平(P<0.05)(图4)。
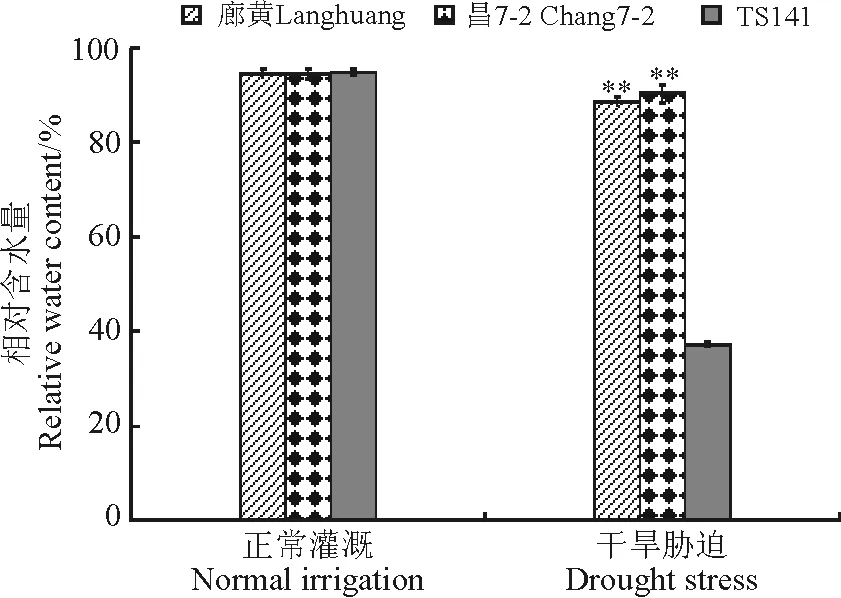
图3 干旱胁迫下玉米耐旱自交系与旱敏感自交系的叶片组织含水量Fig.3 Relative water contents of seedlings of drought-tolerant and -sensitive maize lines under drought stress

图4 干旱胁迫下玉米耐旱自交系与旱敏感自交系的叶片相对电导率Fig.4 Relative conductivities of seedlings of drought-tolerant and -sensitive maize lines under drought stress
2.2.3叶片脯氨酸含量 水分充足条件下,3个自交系廊黄、昌7-2和TS141植株中的Pro含量都比较低,分别为159.44、101.46和185.04 μg·g-1。干旱胁迫处理后,3个玉米自交系的Pro含量均有不同程度的增加,但是耐旱自交系增加幅度较大,高于旱敏感自交系(图5)。干旱胁迫后,耐旱自交系廊黄和昌7-2中的Pro含量分别达到291和270 μg·g-1,而旱敏感自交系中的Pro含量是223 μg·g-1,耐旱自交系中的Pro含量高于旱敏感自交系。
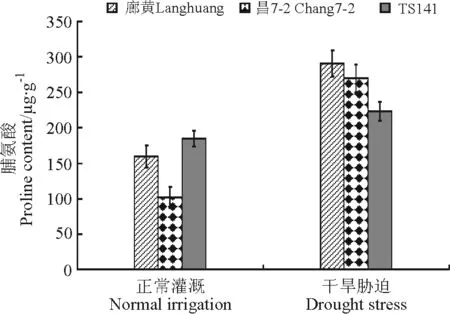
图5 干旱胁迫下玉米耐旱自交系与旱敏感自交系的叶片脯氨酸含量Fig.5 Proline contents of seedlings of drought-tolerant and -sensitive maize lines under drought stress
2.2.4叶片SOD活性 在未受干旱胁迫时,3个玉米自交系都有较低的SOD活性,耐旱自交系廊黄和昌7-2的SOD活性分别为23和49 U·g-1,旱敏感自交系的SOD活性为94 U·g-1;胁迫处理后,3个玉米自交系中的SOD活性都升高,但是耐旱自交系廊黄中的SOD含量显著高于旱敏感自交系(P<0.05)(图6)。
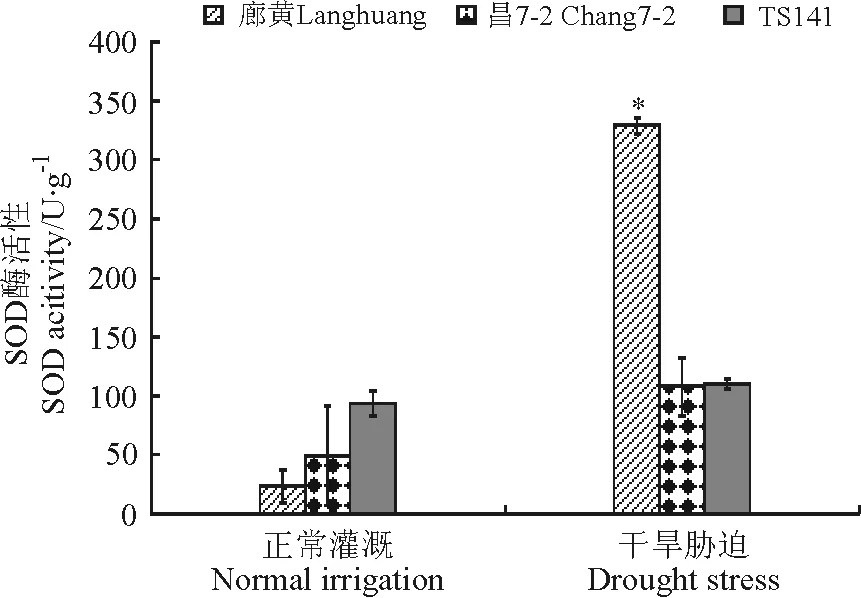
图6 干旱胁迫下玉米耐旱自交系与旱敏感自交系的叶片SOD活性Fig.6 SOD activities of seedlings of drought-tolerant and -sensitive maize lines under drought stress
2.2.5叶片POD活性 在水分充足条件下,3个玉米自交系中的POD活性都很低,但是有一定的差异,这可能是由不同自交系的基因型差异造成的。干旱胁迫后,廊黄、昌7-2和TS141的POD含量都增加了,分别达到47、35和23 U·g-1,耐旱自交系中廊黄和昌7-2的POD活性分别极显著(P<0.01)和显著(P<0.05)高于旱敏感自交系(图7)。
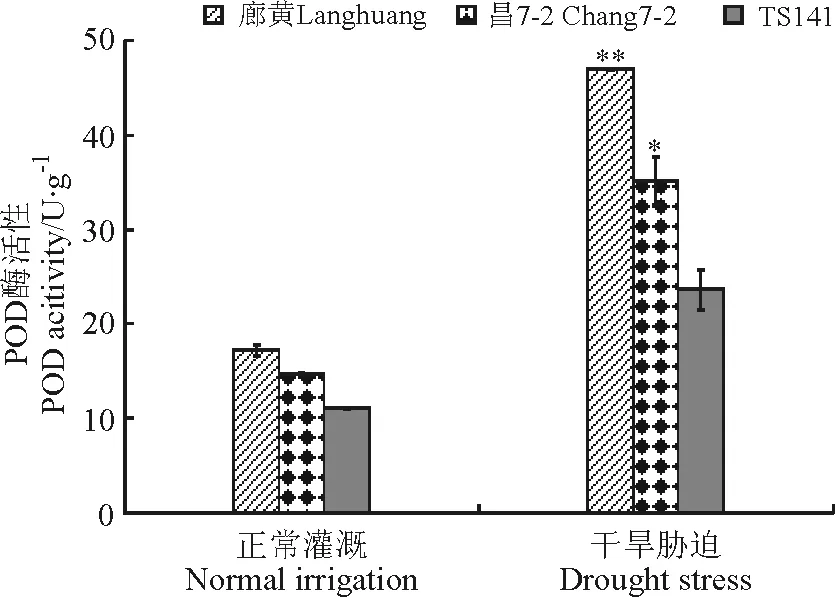
图7 干旱胁迫下玉米耐旱自交系与旱敏感自交系的幼苗叶片POD活性Fig.7 POD activities of seedlings of drought-tolerant and -sensitive maize lines under drought stress
3 讨论与结论
玉米的耐旱机理十分复杂,耐旱性可能存在于生长发育的各个阶段,主要包括萌发期、苗期、开花期和成熟期。不同时期,在耐旱性鉴定中需分别采用相应的形态及生理生化等指标。相对于玉米生长的其他时期,苗期鉴定耐旱性具有时间短、重复性强等优点,并且苗期和成株期抗旱性具有比较高的相关性,因此进行苗期抗旱性研究具有重要意义。
孙彩霞等[10]、王敏等[11]报道干旱胁迫后幼苗的存活率能够从表观上较好地反映植物的耐旱性。本研究表明,TS141在受到干旱胁迫后的存活率最低,对干旱胁迫最为敏感,廊黄存活率最高,对干旱胁迫最不敏感,这与干旱胁迫后耐旱自交系与旱敏感自交系形态特征相一致。
刘惠芬等[12]报道在干旱胁迫条件下,植株的叶片含水量越高,则耐旱性越强,反之则耐旱性越弱。本研究中,干旱胁迫条件下,耐旱自交系廊黄和昌7-2的叶片相对含水量较高,耐旱性较强,而旱敏感自交系TS141的叶片相对含水量较低,耐旱性较弱。
细胞膜透性是反映质膜破坏程度的重要指标,相对电导率可以反映植物细胞膜透性。干旱胁迫条件下,相对电导率升高,说明叶片的质膜受到了伤害,引起细胞内容物质的外渗,而且相对电导率随干旱胁迫程度的加深而不断升高[13-15]。本研究中3个玉米自交系受到干旱胁迫后,相对电导率增加,但是廊黄和昌7-2的增幅较小,TS141的增幅最大,说明旱敏感自交系细胞膜被破坏的程度最大,耐旱性最弱,而耐旱自交系能更好地保护叶片细胞免于受到伤害,有较强的耐旱性。
目前,脯氨酸与植物耐旱性之间的关系说法不一。一种说法认为,当植物受到干旱胁迫时,植物体内脯氨酸的含量积累增加[14,16-17],且耐旱性强的植物种类(自交系)比耐旱性弱的脯氨酸含量上升幅度大[18]。另外一种说法认为,脯氨酸积累量与自交系的耐旱性无关,因为有些在干旱胁迫条件下脯氨酸含量高的材料或植物种类耐旱性并不一定高[19-21]。史燕山等[22]和宋海鹏等[23]报道,随干旱胁迫程度的加重,叶片中游离脯氨酸含量呈增加趋势,增加幅度越大的植物,其抗旱能力越小。本研究表明,在水分充足条件下,不同自交系中脯氨酸的含量有一定的差异,旱敏感自交系TS141中的含量稍微高一点,这可能与玉米的品种不同有关系。在干旱胁迫下,3个玉米自交系受到干旱胁迫时幼苗叶片的脯氨酸含量均增加,但增加的幅度不同,耐旱自交系廊黄和昌7-2增加幅度大,旱敏感自交系TS141增加幅度小,这与第一种说法相一致。
在自然干旱条件下,保护酶活性往往升高,但不同植物以及同种植物不同生长时期上升的程度不同。在本研究中,3个玉米自交系在干旱胁迫后幼苗叶片SOD、POD活性均呈上升趋势,耐旱自交系廊黄幼苗叶片中两种酶的活性上升幅度较大,旱敏感自交系TS141中两种酶的活性上升幅度较小,说明在干旱胁迫下,与旱敏感自交系相比,耐旱系保持了较高的酶活性,具有较强的活性氧清除能力。这一结果与吕长平等[24]和Frazeli等[25]有关植物SOD活性变化与耐旱性强弱关系的研究结果一致,而与袁晓晶[26]和何淼等[27]等报道的保护酶活性随着干旱胁迫程度的加深而呈现出先增加后降低再增加趋势的研究结果不一致,推测可能与不同试验所用的胁迫强度不同有关。
在本研究中,廊黄、昌7-2和TS141在干旱胁迫下表现出了不同的响应机制。干旱胁迫下,廊黄和昌7-2叶片脯氨酸含量增加较快,且叶片具有较强的保护酶活性,细胞膜受到的伤害较轻,使叶片保持了相对高的相对含水量和较低的相对电导率,其耐旱性较强,而TS141的表现则相反。该结果对揭示玉米耐旱机理有重要意义,并且可以为玉米耐旱种质改良和分子育种提供宝贵的材料。
[1] 李晚忱,付凤玲,袁佐清.玉米苗期耐旱性鉴定方法研究[J].西南农业学报,2001,14(3):29-32.
[2] 魏秀俭,杨婉身,潘光堂,等.22个玉米自交系的耐旱性综合分析[J].干旱地区农业研究,2005,23(1):134-137.
[3] 武斌,李新海,肖木辑,等.53份玉米自交系的苗期耐旱性分析[J].中国农业科学,2007,40(4):665-676.
[4] 斐英杰,郑家玲,庾红,等.用于玉米自交系抗旱性鉴定的生理生化指标[J].华北农学报,1992,7(1):31-35.
[5] Beley J D,Oliver M J.Morphological and physiological alterations in adaptation to water stress in wheat[A]. Somero G N,Osmond CB,Bolis C L.Water and Life:Comparative Analysis of Water Relationships at the Organismic,Cellular and Molecular Levels[C].Heidelberg:Springer-verlag,1992:141-160.
[6] 周树峰.玉米耐旱性鉴定及其数量遗传学研究[D].雅安:四川农业大学,2002.
[7] 刘贤德,李晓辉,李文华,等.玉米自交系苗期耐旱性差异分析[J].玉米科学,2004,12(3):63-65.
[8] 邹琦.植物生理学实验指导[M].北京:中国农业出版社,2000:163-164.
[9] 李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000:105-263.
[10] 孙彩霞,武志杰,张振平,等.玉米抗旱性评价指标的系统分析[J].农业系统科学与综合研究,2004,20(1):43-47.
[11] 王敏,张从宇,江文斌.7个玉米自交系萌发和幼苗抗旱性研究[J].种子,2007,26(3):3-6.
[12] 刘惠芬,史铭均,高玉葆.不同种群羊草幼苗对渗透胁迫的反应和生理生态适应[J].南开大学学报(自然科学版),2003,36(2):31-36.
[13] 孟林,毛培春,张国芳,等.17个苜蓿自交系苗期抗旱性鉴[J].草业科学,2008,25(1):21-25.
[14] 许桂芳.PEG胁迫对2种过路黄抗性生理生化指标的影响[J].草业学报,2008,17(1):66-70.
[15] 曲涛,南志标.作物和牧草对干旱胁迫的响应及机理研究进展[J].草业学报,2008,17(2):126-135.
[16] 赵瑞雪,朱慧森,程钰宏,等.植物脯氨酸及其合成酶系研究进展[J].草业科学,2008,25(2):90-95.
[17] 张红萍,牛俊义,轩春香,等.干旱胁迫及复水对豌豆叶片脯氨酸和丙二醛含量的影响[J].甘肃农业大学学报,2008,43(5):50-54.
[18] 欧毅,谢永红,王进.温度和PEG胁迫对甜柿叶片游离脯氨酸累积的影响[J].西南农业学报,2006,19(3):470-472.
[19] 卢少云,陈斯萍,陈斯曼,等.3种暖季型草坪草在干旱条件下脯氨酸含量和抗氧化酶活性的变化[J].园艺学报,2003,30(3):303-306.
[20] 孔德政,于红芳,李永华,等.干旱胁迫对不同品种菊花叶片光合生理特性的影响[J].西北农林科技大学学报(自然科学版),2010,38(11):103-108.
[21] Liu J P,Zhu J K.Proline accumulation and salt stress-induced gene expression in salt hypersensitive mutant ofArabidopsis[J].Plant Physiology,1997,114:591-596.
[22] 史燕山,骆建霞,王煦,等.5种草本地被植物抗旱性研究[J].西北农林科技大学学报(自然科学版),2005,33(5):130-134.
[23] 宋海鹏,刘君,李秀玲,等.干旱胁迫对5种景天属植物生理指标的影响[J].草业科学,2010,27(1):11-15.
[24] 吕长平,石雪晖,杨国顺,等.水分胁迫对草莓叶片SOD活性以及MDA和Vc含量的影响[J].湖南农业大学学报,1996,22(5):451-455.
[25] Frazeli F,Ghorbanli M,Niknam V.Effect of drought on biomass,protein content,lipid zeroxidation and antioxidant enzymes in two sesame cultivars[J].Biologia Plantarum,2007,51(1):98-103.
[26] 袁晓晶.六个自交系菊花的抗旱性研究[D].郑州:河南农业大学,2009:31-45.
[27] 何淼,李文鹤,卓丽环.野菊幼苗对自然干旱胁迫的生理响应[J].草业科学,2011,28(2):1456-1460.
