白僵菌培养基继代培养对蛋白酶(cdep1)、几丁质酶(chit1)基因表达的影响
2012-02-28孙召朋张正坤徐文静汪洋洲潘洪玉李启云
孙召朋, 张正坤, 徐文静, 汪洋洲, 潘洪玉, 李启云*
(1.吉林大学,长春 130062; 2.吉林省农业科学院,长春 130033)
白僵菌(Beauveria bassiana)是目前国内生物防治中应用最广泛的昆虫病原真菌[1],具有防治害虫效果好,不伤害天敌,对人、畜、植物无毒害,容易大量生产,无残毒,害虫不易产生抗性等优点[2],在玉米螟和松毛虫等的防治中已取得很好的防效[3]。研究表明,在穿透寄主体壁的过程中,球孢白僵菌会产生多种降解酶。在穿透昆虫体壁的早期,昆虫病原真菌高水平表达蛋白酶,降解昆虫的体表蛋白[4]。同时,也表达用于降解昆虫体壁的主要组成成分几丁质的几丁质酶[5]。球孢白僵菌在防治应用中菌种资源主要来源于自然直接筛选的菌株,生产的菌株“退化”是防效不稳定的重要原因之一。高毒力菌株经多代培养后毒力退化的现象已被认识[6],但原因不是十分清楚。本研究对两株田间采集分离的野生白僵菌菌株进行了培养基继代培养,通过对亚洲玉米螟的室内毒力测定,评价了继代培养对白僵菌毒力的影响。同时,明确了每一世代白僵菌蛋白酶和几丁质酶基因的表达水平,分析了继代培养对白僵菌毒力及其蛋白酶和几丁质酶基因的影响。
1 材料与方法
1.1 试验材料
供试昆虫:亚洲玉米螟公主岭种群[Ostrinia f ur nacalis(Guenée)]2龄幼虫,由吉林省农业科学院植物保护研究所汪洋洲博士提供。
供试菌株:球孢白僵菌D1-5和D6-2分离自德惠市玉米田的玉米螟僵虫虫体,由本实验室沙土管保存于-80℃超低温冰箱(Ther mo For ma)。
1.2 主要试剂
限制性核酸内切酶Eco RⅠ、Hin dⅢ购自MBI Fer mentas,Trizol reagent购自上海生工,琼脂糖为Bio-Rad公司产品,总RNA抽提试剂盒为大连宝生物工程有限公司(Ta Ka Ra)产品。所用无水乙醇、异丙醇、氯仿等有机溶剂为中国医药(集团)上海化学试剂公司产品,均为分析纯。
1.3 室内毒力测定
1.3.1 白僵菌供试菌株的继代培养
将供试菌株在无菌条件下从沙土管接种在SDAY培养基(质量浓度4%)葡萄糖,1%蛋白胨,2%酵母膏,p H7.0),为F1代,培养20 d左右后刮取孢子,将孢子制备成1×107个/mL浓度的孢子悬液,涂布于SDAY培养基,共进行F2~F5代培养,每个菌株各世代培养10皿,收集每个世代孢子,均配制成1×107孢子/mL浓度的孢子悬浮液供试。
1.3.2 萌发率的测定
取相同量菌悬液接种于SDAY培养基中(终浓度为1×105孢子/mL),26℃培养16 h,测其萌发率,每个菌株重复3次。
1.3.3 室内生物测定方法
将室内饲养、龄期一致的2龄玉米螟幼虫浸入1×107孢子/mL的孢子悬液中5~10 s后,用滤纸吸去多余药液,将试虫转移到正常条件下饲养。每处理4次重复,每重复浸虫20头,并设无菌水处理作为对照。在黑暗环境下培养10 d,每天记录被白僵菌感染的试虫数。
1.3.4 数据统计与分析方法
根据调查数据,计算各处理的校正死亡率,公式如下:
死亡率(P1)=死亡虫数(K)/处理总虫数(N)×100%;
校正死亡率(P2)=[处理死亡率(Pt)-空白对照死亡率(P0)]/[1-空白对照死亡率(P0)];
利用DPS分析软件进行统计分析数据。
1.4 半定量RT-PCR检测继代培养对白僵菌蛋白酶、几丁质酶基因表达的影响
1.4.2 样品收集及总RNA提取
从培养供试菌株的PDA培养基上刮取100 mg分生孢子,采用Trizol法提取总RNA。利用紫外分光光度计测定各处理中总RNA的浓度,并保证A260/A280比值在 1.8~2.0之间,以确保总 RNA纯度。
1.4.2 半定量 RT-PCR[7-11]
利用紫外分光光度计测定样品RNA浓度,每个处理上样量为50 ng总RNA,利用蛋白酶(cdep1)(AY040532.1)、几丁质酶(chit1)(AY145-440.1)及管家基因18S r DNA(GQ302680.1)特异性引物进行c DNA第1链合成。以各基因c DNA第1链为模板进行目的片段PCR,反应中调整反应循环数,以10、15、20个循环每隔5个循环进行一次反应,PCR反应条件为以目的基因条带指数增长期循环数为准确定基因表达量的差异。各基因PCR反应体系为:2μL c DNA产物,0.5μL Taq DNA聚合酶,目的基因(或18Sr DNA基因)上下游引物各1μL,d NTPs 1μL,10×buffer 5μL,Mg Cl2(25 mmol/L)3μL,终体积50μL。反应程序如下:94℃2 min,94℃50 s,58℃(cdep1)/52 ℃ (chit1)/58 ℃(18S r DNA)50 s,72 ℃ 1 min (10、15、20、25个循环),72℃10 min。4℃保存。
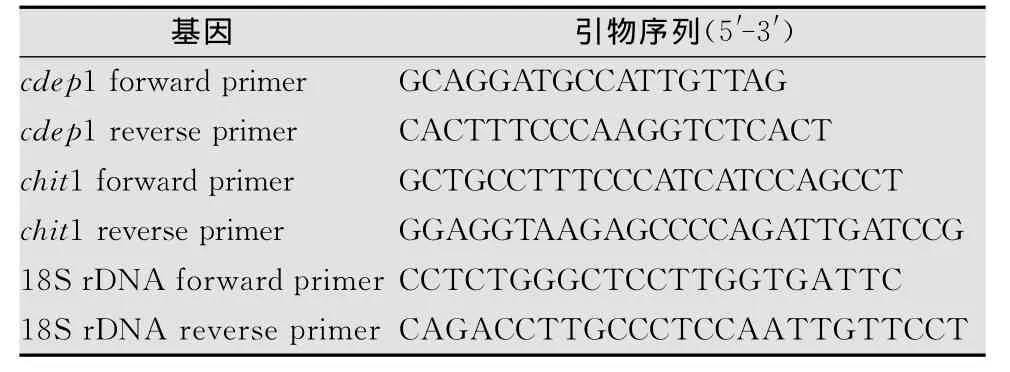
表1 待测基因所用引物序列
根据上述PCR反应结果确定各基因扩增指数期循环数,以供试菌株不同世代各基因c DNA为模板,进行PCR扩增。琼脂糖凝胶电泳胶浓度1.5%。
2 结果与分析
2.1 继代培养对白僵菌孢子萌发率的影响
两菌株在SDAY培养基中经16 h培养后,其萌发率如图1所示。两个菌株的萌发率变化规律基本相似,F2代萌发率明显高于F1代,分别达到77%和70%,在随后的继代培养中,F3、F4、F5代萌发率依次降低,L1-1的F5代萌发率仅为2%。

图1 供试白僵菌菌株不同世代萌发率测定(p=0.05)
2.2 继代培养对白僵菌杀玉米螟毒力的影响
白僵菌两菌株不同世代对亚洲玉米螟2龄幼虫的毒力测定结果见表2和表3。菌株L1-1 F1代孢子对亚洲玉米螟的致死中时(LT50)和致死90%时间(LT90)分别为186.63 h和284.16 h,而F2代孢子分别为180.61 h和209.61 h,明显低于F 1代,而F3~F5代孢子的LT50和LT90逐渐增加,明显高于F2代和F1代,说明其毒力在不断下降。D6-2菌株F1代孢子对亚洲玉米螟的LT50和LT90分别为170.09 h和190.21 h,而 F2代分别为137.70 h和165.84 h,明显低于F1代,而F3~F5代孢子的LT50和LT90逐渐增加,明显高于F2代和F1代,说明随着继代次数的增加其毒力也在不断下降。
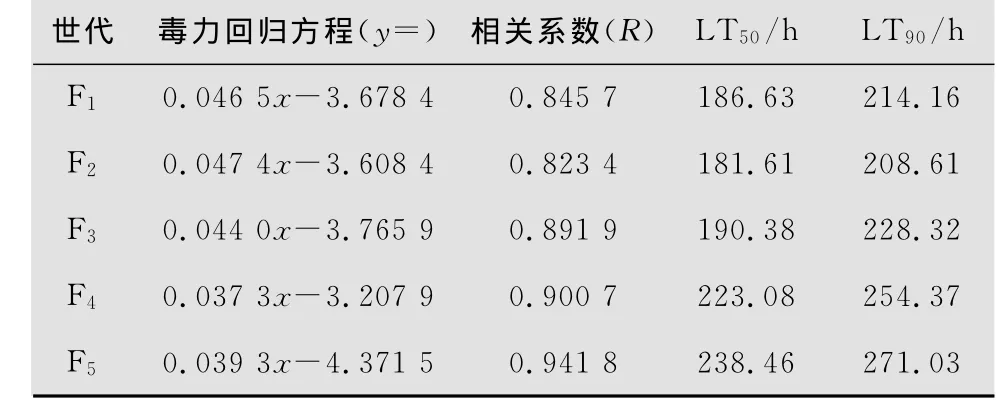
表2 白僵菌L1-1菌株不同世代对亚洲玉米螟2龄幼虫的毒力(10 7孢子/mL)

表3 白僵菌D6-2菌株不同世代对亚洲玉米螟2龄幼虫的毒力(10 7孢子/mL)
2.3 继代培养对白僵菌毒力相关基因表达的影响
2.3.1 RT-PCR 反应循环数的确定
利用供试菌株目的基因和管家基因c DNA第1链为模板,进行目的基因半定量RT-PCR反应循环数的确定。从图2中可以看出,18S r DNA基因和chit 1基因在15~20个循环时条带差异明显,而cdep 1基因在10~15个循环时条带差异明显,因此,以18个循环为18S r DNA基因和chit 1基因的检测循环数,以13个循环作为cdep1基因的检测循环数。

图2 管家基因及毒力相关基因半定量RT-PCR循环数的确定
2.3.2 目的基因的半定量RT-PCR检测
为了明确继代培养对球孢白僵菌毒力相关酶基因表达情况的影响,以18Sr DNA基因为内参,利用半定量RT-PCR对cdep 1基因和chit 1基因的表达情况进行了检测。从图3可以看出,在内参基因稳定表达,也就是总RNA量相同的情况下,两个供试菌株L1-1和D6-2的cdep 1基因和chit1基因在F 1和F2代表达稳定,随着继代次数的增多,供试菌株两个基因的PCR条带明显变弱,说明其表达量在不断下降,这与对亚洲玉米螟毒力测试的结果一致。

图3 继代培养对球孢白僵菌毒力相关酶基因cdep1和chit1基因mRNA表达的影响
3 讨论
毒力测定结果表明,两株白僵菌供试菌株的F2代比F1代对亚洲玉米螟2龄幼虫的毒力有所增强,而F3~F5代对玉米螟的毒力呈下降趋势。球孢白僵菌通过分泌蛋白酶、几丁质酶和脂酶来降解昆虫体壁,其中几丁质酶和蛋白酶具有重要作用[12-14]。本研究利用半定量RT-PCR方法对两株白僵菌中两种毒力相关酶基因在不同世代的表达情况进行了测定,发现随着继代次数的增多,供试菌株两个毒力相关酶基因的表达量均呈下降趋势,与对亚洲玉米螟毒力测定结果一致。出现这种现象可能是由于F1代直接接种于沙土管,菌株并没有表现出很高的萌发率和毒力,在F1活化之后,F2代达到其毒力最大值,F3~F5依次减弱。
在白僵菌继代培养过程中,随着继代次数的增多,会有多个生物性状发生改变。P.Rajanikant h等[15]报道继代培养对产孢量和毒力有很大的影响。唐晓庆等[6]报道了继代培养中菌落都会发生变化,并提出了菌落局变发生的几种可能的遗传机制。唐晓庆等人[16]报道了继代培养对抗旱力的影响,结果显示继代培养能增强菌株的抗旱力,但菌株生活力减弱。樊美珍等[17]通过研究证明,以SDAY为基础培养基的不同C/N比、不同p H培养基中,均未发现菌落局变及菌种退化,SDAY最适于白僵菌生长。本试验中采用SDAY为培养基研究供试白僵菌菌株萌发率、毒力及其相关酶基因的表达,结果表明这些指标均发生了变化,并且表现出相似的变化规律。F1与F2代各项指标基本相似或者略低于F 2代,从F2代开始到F5代,萌发率、毒力及其相关酶基因的表达呈现逐渐降低的趋势,这和唐晓庆等人所报道的继代培养中产生不同类型的分离株,这些分离株多表现为产孢量下降、毒力降低等与生产性状退化类似的变异现象基本相符[18]。其可能的原因是由于F1刚从沙土管中接出,菌株新陈代谢能力下降,经过相当于活化阶段的F1代后,在F2代各种指标都达到最大值,从F2代到F5代的继代培养,菌株毒力相关酶基因表达量逐渐降低,从而导致了毒力的下降。Ansari等[19]对另一广泛应用的生防真菌绿僵菌(Metar hiziu m anisopliae)的3株商业化菌株进行了12代的继代培养,发现其毒力并未发生变化,并将其作为绿僵菌高效菌株的筛选标准。生防真菌的毒力在继代培养过程中可能依赖其自身的遗传背景,本研究中只对两株白僵菌菌株进行了继代培养对毒力及毒力相关因素的影响,是否在白僵菌中存在经继代培养而毒力不发生变化的菌株,还需要进一步研究。
由于白僵菌对靶标昆虫的致病力由多种因素造成[6],本研究只是从蛋白酶和几丁质酶基因的表达方面进行了研究,关于其他影响因素如萌发率、毒素代谢水平等还需进一步研究。同时,在菌种使用过程中,定期进行虫体复壮,可以恢复菌株毒力这一问题上存在争论[20],本试验只对培养基继代培养白僵菌毒力退化情况进行了测定,对于通过虫体进行菌株复壮并未研究,后者可作为以后研究的新方向。
[1] 林海萍,韩正敏,张昕,等.球孢白僵菌研究现状及提高其杀虫效果展望[J].浙江林学院学报,2006,23(5):575-580.
[2] 冯明光.胞外蛋白酶和脂酶活性作为球孢白僵菌毒力指标的可靠性分析[J].微生物学报,1998,38(6):461-467.
[3] 王清海,万平平,黄玉杰,等.虫生真菌在害虫生物防治中的应用研究[J].山东科学,2005,18(4):37-41.
[4] Goettel M S,Hajek A E,Siegel J P,et al.Safety of f ungal biocontrol agents[M]∥Butt T M,Jackon C,Magan N.Fungi as biocontr ol agents,CAB Inter national,2001:347-375.
[5] Char nley A K,St.Leager R J.The role of cuticle-degrading enzy mes in f ungal pat hogenesis in insects[M]∥Cole E T,Hoch H C.Fungal spore disease initiation in plants and animals New York,London:Plenu m Press,1991:267-287.
[6] 唐晓庆,樊美珍,李增智.球孢白僵菌继代培养中菌落局变现象及环境影响因素的研究[J].真菌学报,1996,15(3):45-53.
[7] 周林福,陈离伟,姜云水,等.荧光定量PCR与半定量PCR检测HBV DNA的对比分析[J].中国病理生理杂志,2005,21(5):837,848.
[8] 边杉,王颖,于涛,等.聚丙烯酰胺凝胶银染技术在半定量RT-PCR中的应用[J].中国药科大学学报,2004,35(2):178-182.
[9] 石艳丽,郭学平,王凤山,等.半定量RT-PCR法测定链球菌透明质酸合酶 mRNA 的水平[J].食品与药品,2005,7(2 A):22-24.
[10]徐春兰,汪以真.半定量RT-PCR法分析猪肌肉组织细胞谷胱甘肽过氧化物酶mRNA表达水平[J].中国兽药杂志,2005,39(8):3-6.
[11]陈昕,王保莉,曲东,等.小麦硫转运蛋白基因半定量RT-PCR检测方法的建立[J].西北植物学报,2006,26(2):309-313.
[12]范艳华.球孢白僵菌降解寄主体壁的几丁质酶和蛋白酶的分子改良[D].重庆:西南大学,2006.
[13]Fang Weiguo,Leng Bo,Xiao Yuehua,et al.Cloning of Beauveria bassiana chitinase gene bbchit1 and its application to i mpr ove f ungal strain vir ulence[J].Applied and Envir on mental Microbiology,2005,71(1):363-370.
[14]Yanhua Fan,Weiguo Fang,Yuehua Xiao,et al.Directed evol ution for increased chitinase activity[J].Appl Micr obiol Biotechnol,2007,76:135-139.
[15]Rajanikant h P,Subbarat na m G V,Raha man S J,et al.Effectof frequency of subculturing of different isolates of Beauveria bassiana Vuillemin on their biological pr operties[J].Bio-resource Manage ment,2011,2(1):60-65.
[16]唐晓庆,黄勃,李增智.继代培养对球孢白僵菌抗旱力的影响[J].安徽农业大学学报,1996,23(3):289-292.
[17]樊美珍,李增智,唐晓庆.白僵菌菌种退化及其控制[J].安徽农业大学学报,1996,23(3):239-245.
[18]唐晓庆,唐燕平,李增智.球孢白僵菌菌种退化及其对马尾松毛虫防治的影响[J].安徽农业大学学报,1996,23(3):246-253.
[19]Ansari M A,Butt T M.Effects of successive subculturing on stability,vir ulence,conidial yield,ger mination and shelf-life of ento mopat hogenic f ungi[J].Jour nal of Applied Micr obiology,2011,110:1460-1469.
[20]Hall R A.Effectof repeated subcult uring on agar and passaging t hr ough an insect hoston pat hogenicit y,mor phology,and growt h-rate of Verticilliu m lecanii[J].Jour nal of Invertebrate Pat hology,1980,36(2):216-222.
