新蚜虫疠霉感染致病蚜虫的空间分布特征及其理论抽样技术
2012-02-28和淑琪杜广祖李正跃
陈 斌, 和淑琪, 杜广祖, 李正跃
(云南农业大学植物保护学院,生物多样性控制病虫害教育部重点实验室,生物多样性控制作物病虫害应用技术国家工程研究中心,昆明 650201)
新蚜虫疠霉[Pandor a neoaphidis (Remaudière& Hennebert)Hu mber](虫霉目:虫霉科)是分布范围广、流行性强的蚜科专化性虫生真菌[1-2],常引发桃蚜的流行病,在桃蚜种群自然控制中具有重要作用[2-3],也是云南省昆明地区十字花科蔬菜蚜虫病原真菌的主要种类[4-5]。国内外已对新蚜虫疠霉的分布与生物学[5-7]及其应用[8-9]等方面进行了研究报道。邱丽绚等[10]对小白菜田萝卜蚜(Lipaphis er ysi mi)(原误定为菜缢管蚜Rhopal osiphu m pseudobr assicae)新蚜虫疫霉的空间分布进行了研究,而对于发生流行普遍的新蚜虫疠霉在甘蓝田桃蚜种群的空间分布型及田间调查理论抽样技术尚未见报道。生物种群的空间分布型是生物在特定空间中存在特征,研究虫生真菌在甘蓝田桃蚜种群中的空间分布特征,对深入研究其流行规律、准确预测预报及合理利用具有重要意义。因此,本研究立足于云南特殊的地理气候环境条件下桃蚜及新蚜虫疠霉在十字花科蔬菜中发生流行普遍的实际,系统调查,采用空间分布特征指标分析其空间分布格局,并对其抽样技术进行了拟合,为深入研究和利用该虫生真菌提供理论依据。
1 材料和方法
1.1 调查地点及调查方法
调查地点:设在云南中部昆明市嵩明县蔬菜基地,N25°05′~25°28′,E102°40′~103°20′。属典型的温带、暖温带和北亚热带混合型气候,年均气温14℃,全年分为雨季和旱季,每年6-8月份为雨季,降水充沛,湿度较大,9月到第2年5月为旱季,湿度较低。主要种植蔬菜、玉米、马铃薯和水稻等作物。
调查方法:2009年1-12月,选择种植管理水平基本一致的甘蓝田,每块田随机抽样10株,检查计数每株被感染桃蚜数量。每月上、中、下旬各调查一次,每次调查10块田。统计每块田桃蚜被新蚜虫疠霉感染的数量,用Taylor幂法则测定其聚集度,用Iwao回归方程测定其在不同时间的空间分布型,分析其空间分布特征。
1.2 空间分布型测定
1.2.1 聚集度指标检验
按照 丛 生 指 标[11]、m*/m 指 标[12]、扩 散 系 数C[13]、Ca指标[14]、负二项式分布 K 指标进行检验,各指标的计算公式如下:
丛生指标I:I=S2/m-1,当I>0时为聚集分布;当I=0时为随机分布;当I<0时为均匀分布。
m*/m指标:即平均拥挤度与其平均密度之比,当m*/m>1时为聚集分布;当m*/m=1时为随机分布;当m*/m <1时为均匀分布。
扩散系数C:C=S2/m,当C>1时为聚集分布;当C=1时为随机分布;当C<1时为均匀分布。
Ca指标:Ca=(S2/m-1)/m,当Ca>0时为聚集分布;Ca=0时为随机分布;Ca<0时为均匀分布。
Water负二项式分布K 指标:K=m2/(S2-1)。当K<0时为均匀分布;当K→+∞时为随机分布;当0<K<8时为聚集分布。
1.2.2 线性回归方程检验
Iwao m*-m 检验[15]:m*=α+βm,其中,α为分布的基本成分按大小分布的平均拥挤度,β为基本成分的空间分布图式。当α>0时,个体间相互吸引;当α=0时,分布的基本成分是单个个体;当α<0时,个体间相互排斥。当β>1时为聚集分布;β=1时为随机分布;β<1时为均匀分布。
Tayl or幂法则检验[16]:lg S2=l g(α)+b l g(m)。当b>1时呈聚集分布;b=1时呈随机分布;b<1时呈均匀分布。
1.2.3 聚集原因分析
用Blackit h聚集均数λ[13]:λ=×γ,其中γ为自由度为2 Kc、概率为0.5时的χ2分布值。当λ>2时,其聚集原因是由病原体本身的习性和环境条件或由其中一个因素所引起;当λ<2时,其聚集原因则是由于环境条件所引起。
1.3 抽样技术
2+β-1),式中:n为理论抽样数;x为平均密度(头/样方);t为一定置信度下t分布值。取概率保证值t=1.96,允许误差 D=0.1、0.2、0.3,0.4,0.5;α、β为Iwao回归式中的参数。建立理论抽样模型,计算新蚜虫疠霉感染桃蚜不同虫口密度下所需的理论抽样数。
1.4 数据统计方法
丛生指标、m*/m指标、扩散系数C、Ca指标、负二项式分布K指标均在DPS统计软件[17]进行,被感染蚜量间的方差值采用LSD极小显著差异法进行统计。
2 结果与分析
2.1 新蚜虫疠霉空间分布型
2.1.1 新蚜虫疠霉空间分布的聚集度指标
根据1-12月各调查田块新蚜虫疠霉感染桃蚜的平均感染蚜量(X),方差(V)及平均拥挤度()分析结果,其平均拥挤度为(3.889±0.685)~(58.654±4.560),I指标为0.643~17.978,/m 指标为1.429~2.746 7,Ca指标为0.399~1.738,扩散系数C为1.633~18.968,K 指标为1.242~8.297。根据种群聚集分布的指标值I>0,/m>1,Ca>0,C>1,0<K<8的要求,该研究中新蚜虫疠霉空间分布的/m指标值、Ca指标、扩散系数C和K 指标值均符合聚集分布特征参数值,表明新蚜虫疠霉的空间分布型呈聚集分布格局。
2.1.2 新蚜虫疠霉空间分布特征
将新蚜虫疠霉感染桃蚜的平均密度与方差间的关系用上述幂函数公式进行拟合,结果为α=0.12,b=1.36,经卡方(χ2)检验,χ2=76.85<(359)=124.34,故拟合有效。说明新蚜虫疠霉在甘蓝上桃蚜种群中的分布为聚集分布。
2.1.3 新蚜虫疠霉负二项分布的公共K值
由于α=0.12>0,b=1.36>1,因此新蚜虫疠霉在甘蓝田间的分布型为具公共K值的负二项分布,其个体间互相吸引,负二项分布的公共K值为2.87,卡方值=124.34(df=359),在空间以个体群结构的形式存在,且个体群间也呈聚集分布,个体群平均大小指数(即m+1)也与平均密度呈极显著正相关。
2.2 不同季节新蚜虫疠霉空间分布特征
2.2.1 聚集度指标
不同季节新蚜虫疠霉空间分布的结果表明,1-12月其平均拥挤度为(3.889±0.685)~(58.654±4.56),均满足聚集分布特征值。根据种群聚集分布的指标值I>0/m>1,Ca>0,C>1,0<K<8的要求,本研究中/m指标值、Ca指标、扩散系数C和K指标值均符合聚集分布特征参数值(表1)。表明新蚜虫疠霉在甘蓝田呈聚集分布格局。
2.2.2 空间分布型特征

表1 不同季节新蚜虫疠霉的聚集度指标
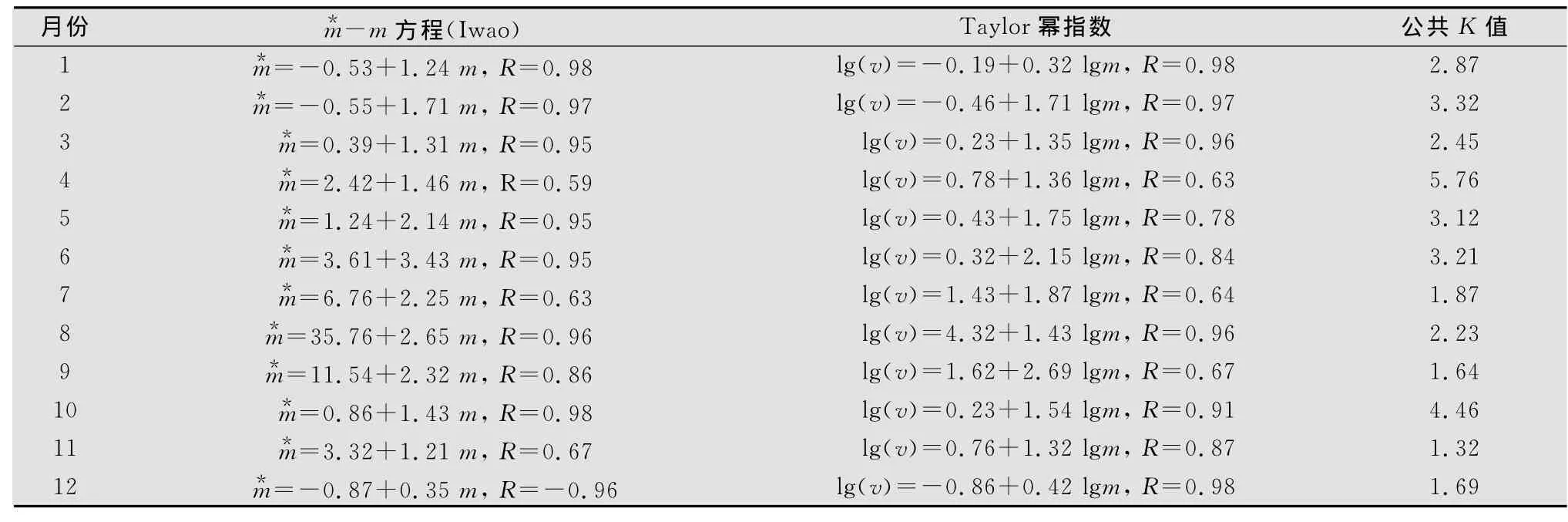
表2 不同时间新蚜虫疠霉空间分布型特征1)
2.3 聚集原因分析
Blackith(1961)[13]提出,昆虫种群空间聚集原因可通过公式λ=×γ(式中:γ为自由度为2 Kc,概率为0.05时的χ2分布值)进行计算和评价。即当λ<2时,种群聚集的原因可能是由于某些环境因素作用的结果;当λ>2时,其聚集原因可能由于病原菌本身的特性所引起,也可能由于昆虫本身的特性与环境的异质性两个因素所引起。

依据上式,当λ=2时,m=0.94。因此,当新蚜虫疠霉感染后罹病桃蚜的平均密度<0.94头/株时,其在甘蓝上的聚集与当时病原菌自身的特性有关;而当平均密度≥0.94头/株时,新蚜虫疠霉在甘蓝田内的聚集分布除与当时病原菌的特性有关外,还与其寄主蚜虫本身的聚集行为有关。
2.4 新蚜虫疠霉空间抽样技术

通过以上理论模型,计算出甘蓝田新蚜虫疠霉在不同罹病桃蚜种群密度下所需的理论抽样数(表4)。
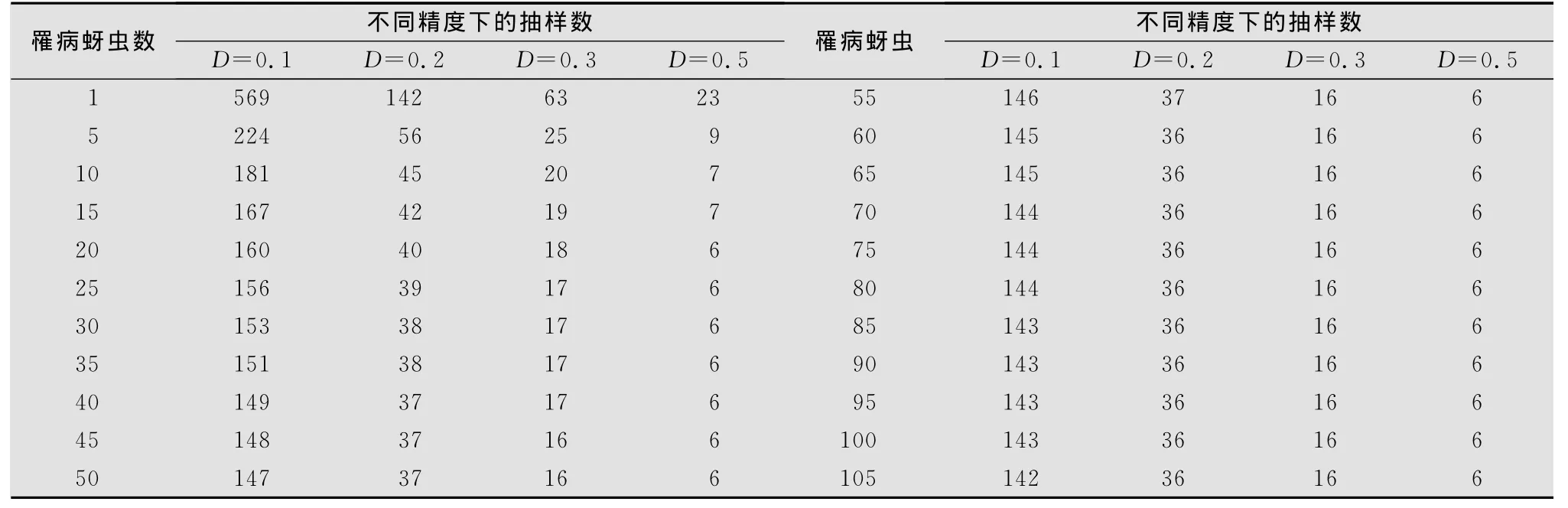
表4 不同罹病桃蚜密度下的理论抽样数
从表4结果看出,当新蚜虫疠霉感染的桃蚜平均密度≤1头/株时,取概率保证值t=1.96,允许误差D=0.1、0.2、0.3和0.5时,其理论抽样数依次为569、142、63株和23株。当平均密度在5头时,所需抽样数分别为224、56、25、9株。而当平均密度在10头以上时,其理论抽样数随虫口密度的升高而平稳下降,虫口密度越高,理论抽样数越少。综合比较4个模型,在新蚜虫疠霉密度较低,即低于每样方5头时,以模型(4)较为适宜,在虫口密度较高,即每样方10头以上时,以模型(3)较宜。
综合近年来在云南菜区新蚜虫疠霉病害流行情况,在6-8月份,对该病害的总体理论抽样以模型(3)理论抽样为宜,即抽查20株左右样方;而在其他季节,每样方虫口密度5头以下时,抽查30株左右的样方。
3 讨论
根据昆虫空间分布评价的5项聚集度指标及Taylor幂法则与Iwao回归方程,新蚜虫疠霉在甘蓝田内桃蚜种群中呈聚集分布中的负二项分布,该结果与邱丽绚等[10]报道的萝卜蚜种群中的新蚜疫霉的空间分布研究结果一致。此外,根据桃蚜在甘蓝田的空间分布特点[18],新蚜虫疠霉的分布型与寄主昆虫的分布型一致,该结果也与Royal et al.[8]提出的新蚜虫疠霉为密度依赖型病原菌结果及关雄等[19]报道的柑橘叶片上粉虱座壳孢菌在柑橘树冠上的空间分布格局与柑橘粉虱在柑橘上的空间分布格局一致的结论相似,这也许正是新蚜虫疠霉病害发生流行性强的重要原因之一。另外,根据一年中新蚜虫疠霉感染桃蚜的聚集强度的季节性变化规律,6-10月份平均拥挤度为全年内最高时段,这与刘树生等[18]报道的桃蚜在一年内聚集高峰期一致,同时也与昆明地区近年来桃蚜种群中新蚜虫疠霉病害的发生流行实际一致[4-5]。
寄主昆虫的习性也是影响昆虫病原微生物聚集特性的重要因素之一[10],新蚜虫疠霉能寄生桃蚜等多种蚜虫[2],在昆明地区多种蚜虫中均有发现[3,5],而甘蓝上除桃蚜外,还有大量的甘蓝蚜,但在平时的调查中发现甘蓝蚜感染率很低[5],因此本研究中仅采用甘蓝上被新蚜虫疠霉感染的桃蚜的密度来分析新蚜虫疠霉的空间分布及理论抽样技术,该结果与关雄等[19]根据柑橘叶片上粉虱座壳孢菌子座数进行取样、分析粉虱座壳孢菌在柑橘树冠上的空间分布格局的结果相似,由此本研究取样方法及其结果可以反映该菌在甘蓝田的空间分布特征。
新蚜虫疠霉的空间聚集分布的基本单位为个体群,当平均感染蚜量≥0.94头/株时,聚集分布的原因除与当时病原菌的特性有关外,还与其寄主蚜虫本身的聚集行为有关,而当新蚜虫疠霉对桃蚜的平均感染蚜量<0.94头/株时,聚集与当时病原菌自身的特性有关,这种特性可能是由于新蚜虫疠霉可通过有翅蚜的迁飞扩散传播[5-7]、捕食性瓢虫的活动取食来扩散和传播[7]特性及该菌分生孢子较强的弹射能力,使该菌在呈聚集分布的桃蚜种群中呈现典型的聚集分布。
[1] Hu mber R A.Fungi:Identification[M]∥Lacey L A.Mannual of Techniques in Insect Pat hology.New Yor k:Academic Press,1997:153-185.
[2] 李增智主编.中国真菌志(第十三卷:虫霉目)[M].北京:科学出版社,2000.
[3] 蒲蛰龙,李增智.昆虫真菌学[M].合肥:安徽科学技术出版社,1996.
[4] 陈斌,李正跃,桂富荣,等.云南省昆明地区菜蚜新蚜虫疠霉发生流行的规律[J].华中农业大学学报,2008,27(2):204-207.
[5] Chen B,Li Z Y,Feng M G.Occurrence of ento mopat hogenic f ungi on migratory alate aphids in Yunnan[J].Biocontrol,2008,53(2):317-326.
[6] Feng M G,Chen C,Chen B.Wide dispersal of aphid-pathogenic entomophthorales among aphids relies upon migratory alates[J].Envir on mental Ento mology,2004,6(5):510-516.
[7] Roy H E,Pell J K,Alderson P G.Targeted dispersal of the aphid pat hogenic f ungus Er ynia neoaphidis by the aphid predator Coccinell a septempunctata [J].Biocontrol Science and Technology,2001,11(1):99-110.
[8] Shah P A,Aebi M,Tuor U.Infection of Macrosiphum euphorbiae with mycelial preparations of Er ynia neoaphidis in a greenhouse trial[J].Mycological Research,2000,104(6):645-652.
[9] Wilding N,Mardell S K,Brobyn P J.Introducing Er ynia neoaphidis into a field population of Aphis f abae:for m of the inoculu m and effectof irrigation[J].Annals of Applied Biology,1990,108(1):373-385.
[10]邱丽绚,黄耀坚,郑本暖.新蚜疫霉流行病的空间分布型[J].昆虫天敌,1989,11(4):1-4.
[11]David F N,Moore P G.Notes on contagious distribution in plant population[J].Annals of Botany,1954,18(1):47-53.
[12]Lloyd M.Mean cro wding[J].Jour nal of Ani mal Ecology,1967,36(1):1-30.
[13]徐汝梅.昆虫种群生态学[M].北京:北京师范大学出版社,1987.
[14]Kuno E.Use of the regression of mean crowing on mean density for esti mating sa mple size and the transfor mation of data for analysis of variance[J].Researches on Population Ecology,1968,10(2):210-214.
[15]Iwao S.A new regression met hod for analyzing the aggregation pattern of ani mal populations[J].Researches on Population Ecology,1968,10(1):1-20.
[16]Taylor L R.Aggregation variance and the mean[J].Nature,1961,189(4766):732-735.
[17]唐启义,冯明光.实用计算机统计分析及其计算机处理平台[M].北京:中国农业出版社,2002.
[18]刘树生,汪信庚,吴晓晶,等.甘蓝和白菜上桃蚜种群的空间格局及其时序动态[J].昆虫学报,1996,39(2):158-164.
[19]关雄,高日霞.粉虱座壳孢菌的空间分布型及其抽样技术[J].福建农学院学报,1991,20(1):29-33.
