Candida etchellsii产2(5)-乙基-4-羟基-5(2)-甲基-3(2H)-呋喃酮的影响因素*
2012-01-12冯杰詹晓北季新跃郑志永王栋张丽敏
冯杰,詹晓北,季新跃,郑志永,王栋,张丽敏
1(江南大学生物工程学院糖化学与生物技术教育部重点实验室,江苏无锡,214122)2(无锡市恒禾工程咨询设计有限公司,江苏 无锡,214031)
2(5)-乙基-4-羟基-5(2)-甲基-3(2H)-呋喃酮(简称HEMF)主要存在于酱油中,是酱油中的主体香味物质[1],故称为酱油酮[2],具有高强度的甜焦糖酱香特色[3],是杂环香体类呋喃同系衍生物中极其重要而且用途极为广泛的香料之一[4]。HEMF可代替天然香料添加在酿造食品如调味料、酒、无酒精饮料及果制品如果酱、果浆、罐头等食品中,具有加强其天然香味的作用[5]。同时,HEMF是美国食用制造者协会(FEMA)认可的安全食用香料。此外,最近日本学者研究发现HEMF还具有抗肿瘤的活性[6]。
国内目前还没有关于对发酵法生产HEMF的研究和报道,国际上也鲜有报道。根据文献报道,张海林等[8]在研究酵母菌产呋喃酮(HDMF和HEMF具有相似的结构和性质)类物质时发现CaCl的添加对酵母菌株产 HDMF有较大的促进作用,Sugawara等[9]研究了酵母菌发酵产味噌的过程中产HEMF,产量在100 ppm以下,并且进一步研究了HEMF的形成机制。Ohata等[3]利用13C同位素示踪方法发现酵母菌发酵产HEMF的形成机制,主要是由戊糖和游离氨基酸通过美拉德反应产物经过酵母菌产生的酶作用后形成HEMF。
本研究以1株从酱油中筛选得到的产2(5)-乙基-4-羟基-5(2)-甲基-3(2H)-呋喃酮的耐高盐度酵母菌株C.etchellsii CICIM Y0600作为研究对象,主要研究了在不同盐度和添加不同种类氨基酸条件下,菌体生长和产物HEMF合成的变化情况。
1 材料与方法
1.1 菌种
Candida.etchellsii CICIM Y0600,江南大学生化工程与反应器研究室保藏。
1.2 培养基
斜面培养基(g/L):蛋白胨10,葡萄糖 20,酵母粉 5,NaCl 180~240,琼脂粉 1.5,pH 5.8 ~6.0。
种子培养基(g/L):蛋白胨10,葡萄糖 20,酵母膏 5,NaCl 180~240,pH 5.8~6.0。
发酵基础培养基(g/L):葡萄糖 75,木糖 25,不同种类氨基酸 5,酵母粉 1,KH2PO410,MgSO4·7H2O 5,NaCl 180 ~240,pH 5.2。
1.3 实验设备
发酵罐:美国New Brunswick Scientific公司BioflowⅢSystem 7 L发酵罐。
1.4 分析方法
生物量的测定:取菌悬液5 mL,10 000 r/min离心10 min弃上清液后加入5 mL水稀释后在分光光度计上于600 nm下测定吸光度(结果与干重法比对,换算为g/L)。
智能变电站是基于数字化建设,因此基于智能变电站的硬件是数字的基础上,通过先进的传感器和通信网络,从而实现全景采集变电站信息,和一个全面的数据库的建立,使变电站自动化操作,维护和智能分析与决策能力实现,进一步提高电力企业的管理水平和适应能力。
还原糖的测定:DNS比色法[10]。
HEMF的测定:高效液相色谱(HPLC)法。条件:高效液相色谱仪为安捷伦1200,色谱柱为安捷伦ZORBAX XDB-C18 柱(150 ×4.6 mm,5 μm),柱温25℃;进样量为20 μL,检测波长265 nm。流动相流速0.8 mL/min,流动相组成:60%甲醇和40%双蒸水。
2 结果与分析
2.1 C.etchellsii分批发酵过程曲线
以耐高盐度的C.etchellsii为研究对象,选择培养基中的甘氨酸作为氨基酸源,在180 g/L NaCl浓度条件下对其产HEMF进行分批发酵研究,结果如图1所示。

图1 180 g/L NaCl浓度下C.etchellsii生长和产HEMF过程曲线
由图1中的C.etchellsii生长和产HEMF过程曲线可知,菌体生长分为3个阶段,0~4 h为菌体生长的迟滞期,此后4~40 h菌体生长处于对数期,产物HEMF开始产生,在40 h后菌体生长进入稳定期,最大菌体干重为11.11 g/L,而产物HEMF在120 h达到最大值57.74 mg/L。
2.2 不同NaCl浓度对C.etchellsii产HEMF影响
张海林等[8]在研究酵母菌产呋喃酮(HDMF和HEMF具有相似的结构和性质)类物质时发现,NaCl的添加对酵母菌株产HDMF有较大的促进作用。由于本研究中C.etchellsii筛选自高盐稀态发酵酱油中,该类型酱油中的NaCl浓度为180~240 g/L,故在发酵基础培养基中补充5 g/L甘氨酸,然后分别向发酵体系中添加180 g/L,200 g/L,220 g/L和240 g/L NaCl,考察NaCl浓度对C.etchellsii合成HEMF的影响。选择发酵140 h时结束并分别对其生物量和HEMF产量进行测定,结果如表1所示。
由表1可以看出,在200 g/L NaCl浓度条件下,菌体浓度最高,而在220 g/L NaCl浓度条件下,HEMF产量和对菌体浓度的产物得率最高。这说明菌体生长和产物合成的最适条件不一致,这种情况下通常采用分阶段调控的策略,最大程度的同时满足菌体生长和产物合成需要。
选择初始的NaCl浓度为200 g/L,在发酵20 h,40 h和60 h分别补加NaCl至220 g/L,发酵140 h时结束并测定结果,如表2所示。
由表2中的数据分析可知,选择初始200 g/L NaCl浓度,在发酵20 h,40 h和60 h分别补加NaCl至220 g/L后,发酵至140 h测得HEMF最终发酵产量分别为76.79 mg/L,98.11 mg/L和89.28 mg/L。可见,利用两阶段添加NaCl策略在发酵40 h时补加NaCl在保证最大菌体浓度的同时,HEMF产量最高。
2.3 添加不同氨基酸对C.etchellsii产HEMF影响
Ohata等[3]利用13C同位素示踪方法发现酵母菌发酵产HEMF的形成机制,主要是由戊糖和游离氨基酸通过美拉德反应后的产物经过酵母菌产生的酶作用后形成HEMF。因此,氨基酸的添加对HEMF的合成有影响。为了考察氨基酸对C.etchellsii合成HEMF的影响,选择200 g/L NaCl为条件,分别选择20种游离氨基酸为氨基酸源对其产HEMF进行研究,在发酵140 h时对其菌体浓度和HEMF含量进行测定,结果如图2所示。

图2 添加不同氨基酸对C.etchellsii产HEMF影响结果
由图2中20种游离氨基酸对 C.etchellsii产HEMF影响结果可知,除酪氨酸(Tyr)对C.etchellsii产HEMF有抑制作用外,其他19种氨基酸对其产HEMF均有不同程度的促进作用,其中,丙氨酸(Ala)、精氨酸(Arg)、甘氨酸(Gly)和赖氨酸(Lys)的促进作用最明显,在这4种条件下,HEMF的产量分别为76.77 mg/L、97.85 mg/L、88.45 mg/L 和 67.11 mg/L。
以添加不同氨基酸对C.etchellsii产HEMF影响为基础,选择200 g/L NaCl浓度,进一步考虑不同种类的氨基酸组合对C.etchellsii产HEMF的影响。选取单个添加中产量较高的丙氨酸(Ala)、精氨酸(Arg)、甘氨酸(Gly)和赖氨酸(Lys)为氨基酸源,对其进行组合,在发酵140 h时对其菌体浓度和HEMF含量进行测定,结果如表3所示。
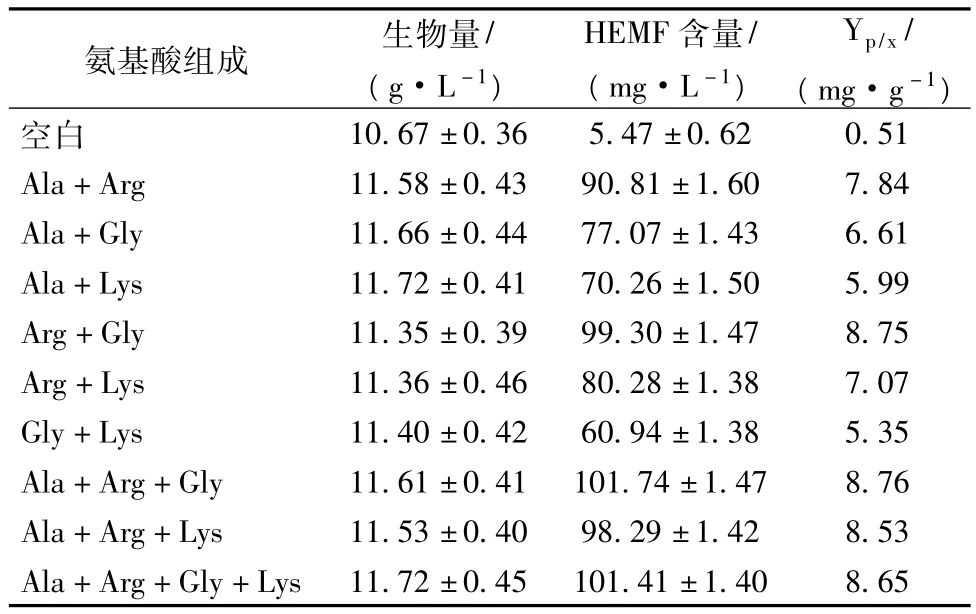
表3 氨基酸组合添加对C.etchellsii产HEMF影响结果
由表3中的数据分析可知,氨基酸组合对C.etchellsii产HEMF有不同的影响,其中 Ala+Arg+Gly组合中HEMF产量和对菌体的产物得率最高,分别为101.74 mg/L和8.76 mg/g。而Ala+Arg+Gly+Lys组合中的HEMF产量和对菌体的产物得率也较高,分别为101.41 mg/L和8.65 mg/g,与Ala+Arg+Gly组合中差别不大,由于考虑到实际操作的可行性和简便性,因此可以确定Ala+Arg+Gly组合可以更好地促进C.etchellsii产HEMF。
2.4 NaCl分阶段调控结合氨基酸添加对C.etchellsii生产HEMF影响
为了进一步强化 HEMF的发酵生产强度,将NaCl分阶段调控与氨基酸添加结合应用。选择200 g/L NaCl为条件,根据2.2中的研究结果分别在40 h和60 h补加NaCl至220 g/L。氨基酸源选用Ala+Arg+Gly组合,在发酵140 h时对其菌体浓度和HEMF含量进行测定,结果如表4所示。

表4 两阶段NaCl添加策略和氨基酸组合添加策略结合对C.etchellsii产HEMF影响结果
由表4中的结果分析可知,两阶段NaCl添加策略和氨基酸组合添加策略结合对 C.etchellsii产HEMF均有不同程度的促进作用,其中在发酵40 h时添加NaCl至220 g/L,并结合Ala+Arg+Gly组合时,C.etchellsii产HEMF和对菌体的产物得率最高,分别为110.74 mg/L和9.51 mg/g。
2.5 7 L发酵罐验证2种添加策略对C.etchellsii产HEMF影响
为了验证两阶段NaCl添加策略和氨基酸组合添加策略相结合对C.etchellsii产HEMF影响,在7 L发酵罐上对C.etchellsii产HEMF进行验证研究,结果如图3所示。

图3 两阶段NaCl添加策略和氨基酸组合添加对C.etchellsii产HEMF上罐验证结果
由图3中的过程曲线可知,发酵40 h时C.etchellsii生长进入稳定期,最大菌体浓度为10.91 g/L,在100 h HEMF产率最大,为121.51 mg/L,比空白(200 g/L NaCl且没有添加氨基酸,图3中未标明)提高了21.2倍。
3 讨论
实验以1株增香酵母菌C.etchellsii为研究对象,研究了在不同盐度和添加不同种类氨基酸条件下,C.etchellsii菌体生长和产物HEMF合成的情况。
由结果分析可知,发酵体系中添加200 g/L和220 g/L NaCl利于 C.etchellsii合成 HEMF,当 NaCl浓度增加到240 g/L时不利于菌体的生长和产物HEMF的生成。分析原因是菌体细胞的培养环境与菌体生长密切相关,由于C.etchellsii筛选自高盐稀态发酵酱油中,该类型酱油中的NaCl浓度为180~240 g/L,因此,经过高盐度环境的驯化,C.etchellsii已经适应在高浓度NaCl环境中生长。但是,当NaCl浓度过高(≥240 g/L)时,超过了C.etchellsii的适应极限,此时菌体生长缓慢,菌体浓度较小。因此,采取分阶段调控NaCl浓度的策略,可在保证C.etchellsii正常生长的同时获得高产量的HEMF。而根据文献报道[8],高盐度环境利于HDMF和HEMF等呋喃酮类物质的形成,根据C.etchellsii生长的高盐度环境猜测原因可能是在适合菌体生长的环境下,高盐度使得菌体细胞膜的通透性结构有所改变,对底物的利用具有更高的选择性,使得其代谢向利于HEMF生成的方向进行。氨基酸作为C.etchellsii代谢产HEMF的前体物质,在HEMF的产生过程中起重要作用,因此选择不同种类和不同类别的氨基酸组合对于HEMF的生成是很重要的,通过研究结果可知,选择Ala+Arg+Gly组合更利于HEMF的生成,这可能是C.etchellsii菌体内源性形成这3种氨基酸的效率较低,需外援补充以满足HEMF合成的需要。当三者以合适的比例组合添加到发酵体系中时,对C.etchellsii菌体内HEMF的合成产生了协同促进效应,从而提高了HEMF的产量。
[1] Blank I,Fay L B.Formation of 4-hydroxy-2,5-dimethyl-3(2H)-furanone and 4-hydroxy-2(or 5)-ethyl-5(or 2)-methyl-3(2H)-furanone through Maillard reaction based on pentose sugars[J].Journal of Agricultural and Food Chemistry,1996,44(2):531-536.
[2] Li X,Hiramoto K,Yoshida M,et al.Identification of 2,5-dimethyl-4-hydroxy-3(2H)-furanone(DMHF)and 4-hydroxy-2(or 5)-ethyl-5(or 2)-methyl-3(2H)-furanone(HEMF)with DNA breaking activity in soy sauce[J].Food and Chemical Toxicology,1998,36(4):305-314.
[3] Ohata M,Kohama K,Morimitsu Y,et al.The formation mechanism by yeast of 4-hydroxy-2(or 5)-ethyl-5(or 2)-methyl-3(2H)-furanone in miso[J].Bioscience Biotechnology and Biochemistry,2007,71(2):407-413.
[4] Sasaki M,Nunomura N,Matsudo T.Biosynthesis of 4-hydroxy-2(or 5)-ethyl-5(or 2)-methyl-3(2H)-furanone by yeasts[J].Journal of Agricultural and Food Chemistry,1991,39(5):934-938.
[5] Cao X H,Hou L H,Lu M F,et al.Improvement of soysauce flavour by genome shuffling in Candida versatilis to improve salt stress resistance[J].International Journal of Food Science and Technology,2010,45(1):17-22.
[6] Hauck T,Landmann C,Bruhlmann F,et al.Formation of 5-methyl-4-hydroxy-3[2H]-furanone in cytosolic extracts obtained from Zygosaccharomyces rouxii[J].Journal of Agricultural and Food Chemistry,2003,51(5):1 410 -1 414.
[7] 张精安.2,5-二甲基-4-羟基-3(2H)呋喃酮的合成研究[J].精细化工,2001,18(1):31-33.
[8] 张海林,范文来,徐岩.高产2,5-二甲基-4-羟基-3(2H)-呋喃酮(DMHF)酵母菌株的选育[J].中国酿造,2009(11):20-23.
[9] Sugawara E,Sakurai Y.Effect of media constituents on the formation by halophilic yeast of the 2(or 5)-ethyl-5(or 2)-methyl-4-hydroxy-3(2H)-furanone aroma component specific to miso[J].Bioscience Biotechnology and Biochemistry,1999,63:749-752.
[10] He L,Xu Y Q,Zhang X H.Medium factor optimization and fermentation kinetics for phenazine-1-carboxylic acid production by Pseudomonas sp.M18G[J].Biotechnology and Bioengineering,2008,100(2):250-259.
