不同底质改良处理对三种挺水植物光合特性的影响*
2011-12-11包先明范成新史刚荣
包先明,范成新,史刚荣
(1:淮北师范大学生命科学学院,淮北235000)
(2:中国科学院南京地理与湖泊研究所,南京210008)
不同底质改良处理对三种挺水植物光合特性的影响*
包先明1,范成新2,史刚荣1
(1:淮北师范大学生命科学学院,淮北235000)
(2:中国科学院南京地理与湖泊研究所,南京210008)
以巢湖水域三种优势挺水植物为材料,研究了不同底质处理下植物光合生理特性的差异.结果表明,三种挺水植物地上部生物量以香蒲最大,芦苇次之,菖蒲最小.香蒲的叶绿素a(Chl.a)、叶绿素b(Chl.b)和类胡萝卜素(Car)含量、气孔导度、胞间CO2浓度、蒸腾速率均显著大于芦苇和菖蒲,光合速率、Fv/Fm和ΦPSⅡ显著高于菖蒲.相关性分析表明,地上部分生物量的变化与光合速率呈显著正相关.净光合速率与气孔导度、胞间CO2浓度均呈显著正相关关系.因此,较大的气孔导度、较高的光合色素含量以及Fv/Fm和ΦPSⅡ,是香蒲之所以具有较高净光合速率和生物量的重要机制.四种底质改良方式(划耕、有机质、客土和加沙)均能不同程度地提高植物叶片的光合色素含量(Chl.a、Chl.b、Car和Chl.a/Chl.b)和光化学反应中心活性(Fv/Fm和ΦPSⅡ),增加气孔导度和胞间CO2浓度,从而提高光合速率并最终促进植物的生长.其中,以划耕和有机质处理的效果最为显著.
挺水植物;底质;气体交换;光合色素;叶绿素荧光
近年来,随着我国经济的迅速发展,排污量日益增加,大量含有氮、磷元素的污染物排入湖(库),使水体的营养物质负荷量不断增加,造成水体富营养化[1].我国目前66%以上的湖泊、水库处于富营养化水平,其中重富营养和超富营养的占22%.国内外对湖泊水体富营养化控制已进行了较多的研究和报道[2-4],但过去的富营养化防治主要集中在理化方法和工程措施,很少注意到生态系统结构和功能的调整[5].
水生植物污水净化技术相对其他水体修复技术具有以下优点[6-8]:1)能有效地将不可溶性水体底部的有机物降解为可溶性有机物;2)能抑制蓝藻及其他耗氧强的菌类与藻类的生长与繁殖,增加水中的溶氧量;3)分解水中含氮有机物,最终能使铵态氮和硝态氮大大减少,提高水生生物的成活率;4)不向水体投放药剂,不会形成二次污染;5)工程造价较低,不需耗能或低耗能;6)用生物生态方法设计水景,有利于降低建设和养护管理成本,并易创造人与自然相融合的优美环境.因此,利用水生植物修复受污水体的研究受到日益广泛的关注[9].挺水植物不仅本身能吸收污水中氮、磷,拦截、过滤污染物,同时还能提高整个湿地生态系统微生物数量,调整其群落结构,促进湿地生态系统的硝化和反硝化作用,强化其净化能力.另外,部分植物还有克藻效应[10].利用挺水植物构建人工湿地和恢复重建自然湿地,已成为湖泊水体富营养化控制的关键措施.
由于不同植物的生理特性不同,对水体环境的适应性也不同.因此,选择适合水生环境特点的植物以及采用适宜的栽培措施是水生植物净化修复技术的关键所在.光合作用是植物生长发育的基础,受遗传特性和环境因素的相互影响.湿地植物通过光合产氧改善湿地氧环境,有利于污染物的净化[11-12].雒维国等[13]发现,随着芦苇净光合速率的增加,湿地溶解氧浓度上升,脱氮能力得到提高.黄娟等[14-15]的研究表明,植物净光合速率与湿地总氮去除率呈显著正相关.卢晓明等[16]发现磷、铵态氮的去除率与蒸腾作用和光合作用的相关性显著.光合效率不仅是估计植物潜在生产力和对环境适应能力的重要指标,同时也是衡量植物净化能力的重要参数.目前,对湿地植物的光合特性等的研究已有报道[17-18],但关于不同底质对挺水植物光合特性的影响及其与植物种的交互效应方面的研究未见报道.
本研究以巢湖流域三种优势挺水植物为材料,对不同底质处理下植物光合生理特性差异进行比较,揭示其光合生理生态特征对底质处理的响应机制,旨在为湖泊生态系统的底质修复与水生植被重建提供科学依据.
1 材料与方法
1.1 试验设计
实验选用的3种挺水植物为:芦苇(Phragmites communis)、香蒲(Typha minina)、菖蒲(Acorus calamus),所有实验水生植物全部是来自巢湖水域的土著物种.
根据沉积物分布的特点,于2009年8月20日在巢湖流域水域选取了一个未经疏浚的采样点,用沉积物采样器采集表层沉积物样品,同时在现场用若干25L加仑桶采集样点处湖水,一并带回实验室.沉积物共设计为5种处理:①原状(即对照,不作任何处理);②加土(在沉积物表层覆盖5cm厚的疏松土壤);③ 加沙(在沉积物表层覆盖5cm厚的细沙);④加有机质(在表层5cm沉积物中均匀加入200g水稻秸杆);⑤划耕(用金属铁丝对表层5cm沉积物划松).每个处理设3个重复.采集的沉积物样品经各种处理后分别置入聚乙烯塑料桶(高110cm,内径70cm)中,保持沉积物厚度30cm,将水生植物的幼苗分别移种到实验塑料桶中.覆盖实验所用的土样直接取于巢湖岸边未经污染的土壤样品,自然风干研磨后过20目筛备用;细沙则取自湖岸的建筑工地,然后过20目筛备用;水稻秸杆取自湖岸农田,晒干,粉碎过20目筛备用.对采集的大桶水样通过25号浮游生物网过滤后制得上覆水样,将制得的上覆水分别缓慢注入各种处理的塑料桶中,保持水面距桶口高度15cm,放在露天实验场进行水生植物生长培养实验.
1.2.测定指标与方法
将植株齐土面剪断,测量植株高度后,在105℃下杀青30min,然后在65℃下烘干至恒重,测定其生物量.植物叶绿素含量的测定采用80%丙酮与无水乙醇1∶1体积混合浸提法,用UV-4802型紫外可见光分光光度计在663、645、470nm波长下测光密度OD值,叶绿素含量基于叶片鲜重,以mg/g(FW)表示.叶绿素荧光参数用Mini PAM(Walz,Effeltrich,德国)测定.植株叶片暗适应30min后,测定暗适应后叶绿素的最小荧光(Fo)和红光饱和脉冲后的最大荧光(Fm),据此计算可变荧光(Fv,最大荧光和最小荧光之差)和可变荧光和最大荧光之比(Fv/Fm,是光系统Ⅱ光化学反应量子产额的量度).在光化光1000μmol/(m2·s)下测定瞬时荧光(Fs),在1s的5000μmol/(m2·s)饱和脉冲下测定最大荧光(Fm'),计算光系统Ⅱ有效量子产额(ΦPSⅡ):ΦPSⅡ=ΔF/Fm'=(Fm'-Fs)/Fm'.净光合速率(Pn)、气孔导度(Gs)、细胞间 CO2浓度(Ci)和蒸腾速率(E)测定在植株顶部向下第一完全展开功能叶进行.测定仪器为Li-6400便携式光合作用测定系统(Li-Cor Inc.,美国),测定时使用开放式气路.用LED光源叶室(6400-02)控制测定条件:CO2浓度为380±15μmol/mol,光强为1000μmol/(m2·s),叶室温度 25 ±2℃.
1.3 统计分析
原始数据用SPSS for windows 11.5进行统计分析,利用one-way ANOVA分析不同底质处理对植物各项指标的影响及这些指标的种间差异,若主效应显著,用Duncan检验进行多重比较,确定相应指标在不同处理间的差异是否显著(P<0.05).利用two-way ANOVA分析底质处理和植物种对光合生理参数的交互效应.用相关性分析揭示各光合参数间的相互关系.
2 结果与分析
2.1 不同底质对植株生长的影响
在不同底质处理下,香蒲和芦苇的株高均显著大于菖蒲,而在香蒲和芦苇之间差异不显著.香蒲的地上部生物量在任何底质处理下均显著高于芦苇和菖蒲,但芦苇和菖蒲生物量之间的差异仅在划耕和有机质处理下达到显著水平.与原状(对照)处理相比较,不同底质处理均能显著提高香蒲和芦苇的株高和生物量,其中以划耕和有机质处理下,株高和生物量的增幅最为显著.方差分析表明,底质和植物种对株高具有显著交互作用(P<0.01),但对生物量的交互作用不显著(P<0.05)(图1).不同底质处理对植物最终生物量的影响与生长前期结果一致,以香蒲的生物量最大,且均以有机质和划耕处理生物量增加最为显著(表1).
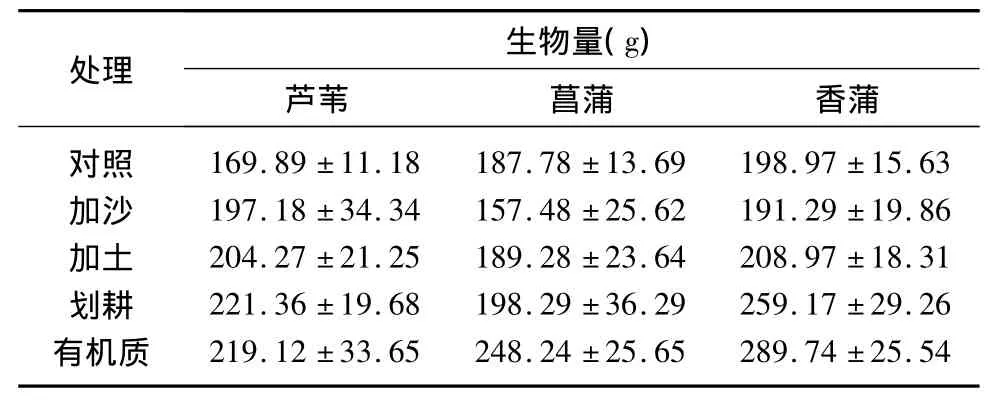
表1 挺水植物最终生物量Tab.1 Biomass of three emergent plant species
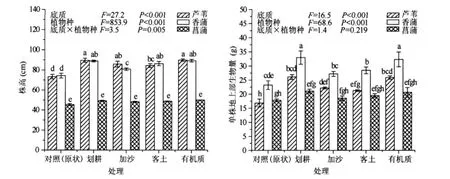
图1 不同底质对三种挺水植物株高和生物量的影响(各图的左上方数据为二因素方差分析的结果;误差线上不同字母表示经Duncan检验数据之间的差异显著(P<0.05),下同)Fig.1 Effects of sediment types on plant height and shoot biomass in three emergent plants
2.2 不同底质对光合色素的影响
叶绿素 a(Chl.a)、叶绿素 b(Chl.b)和类胡萝卜素(Car)含量以及叶绿素 a与叶绿素 b比值(Chl.a/Chl.b)既在三种挺水植物之间存在显著的种间差异,又显著受不同底质处理方式的影响,同时还显著受到植物种和底质处理方式的交互作用影响(图2).在三种植物之间,Chl.a和Car含量以香蒲最高,芦苇次之,菖蒲最低;香蒲叶片的Chl.b含量显著高于芦苇和菖蒲,而在后两者之间的差异不显著;Chl.a/Chl.b以菖蒲最低,且与香蒲和芦苇存在显著差异,而在香蒲和芦苇之间差异不显著.在不同底质处理之间,与对照(原状)相比,四种底质处理方式不仅能显著增加三种挺水植物叶片的Chl.a、Chl.b以及Car含量,还能提高Chl.a/Chl.b,以划耕和有机质处理的效果最为显著(图2).
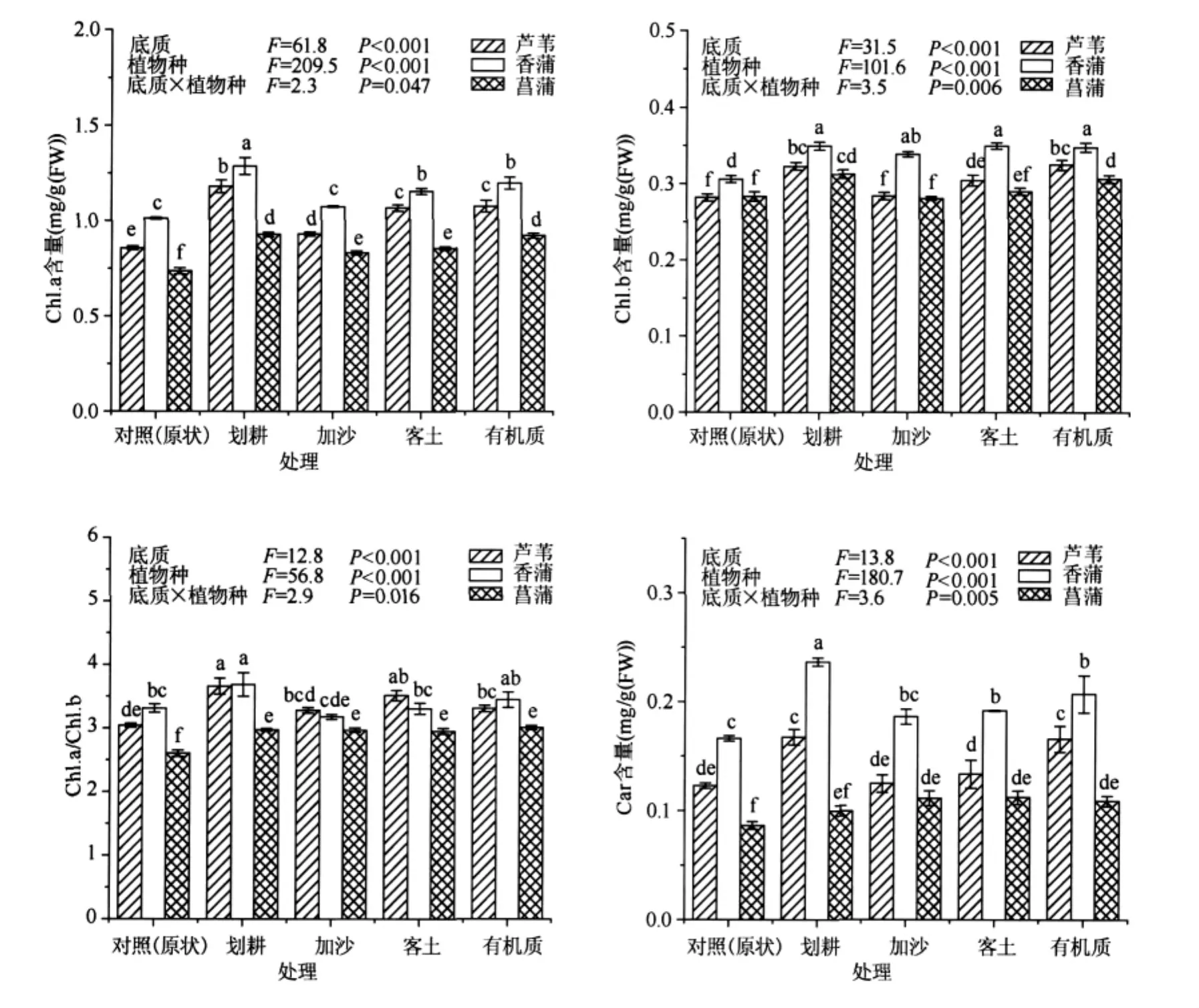
图2 不同底质对三种挺水植物光合色素的影响Fig.2 Effects of sediment types on pigment content in three emergent plants
2.3 不同底质对叶绿素荧光参数的影响
三种挺水植物叶绿素荧光参数存在显著差异(图3).在不同底质修复条件下,香蒲均表现出较高的Fv/Fm和ΦPSⅡ,而菖蒲的Fv/Fm和ΦPSⅡ均较小.芦苇的Fv/Fm在原状、划耕和有机质处理下显著大于菖蒲,与香蒲没有显著差异;在加沙和客土处理下,显著小于香蒲,与菖蒲没有显著差异.芦苇的ΦPSⅡ在原状、划耕和加沙处理下显著小于香蒲,与菖蒲没有显著差异;在有机质和客土处理下,显著小于香蒲.
不同底质处理对叶绿素荧光参数也有显著影响(图3).香蒲和菖蒲的Fv/Fm在划耕、客土、加沙和有机质处理下均显著高于对照(原状),芦苇的Fv/Fm在划耕和有机质处理下也显著高于对照(原状),但在客土和加沙处理下与对照(原状)的差异不显著.香蒲和芦苇的ΦPSⅡ在划耕、客土和有机质处理下均显著高于对照(原状),但在加沙处理下与对照(原状)的差异不显著.对于菖蒲,除划耕处理能显著提高叶片ΦPSⅡ外,其它底质处理对ΦPSⅡ的影响均不显著.方差分析结果表明,底质和植物种对Fv/Fm和ΦPSⅡ具有显著交互作用(P<0.01)(图3).
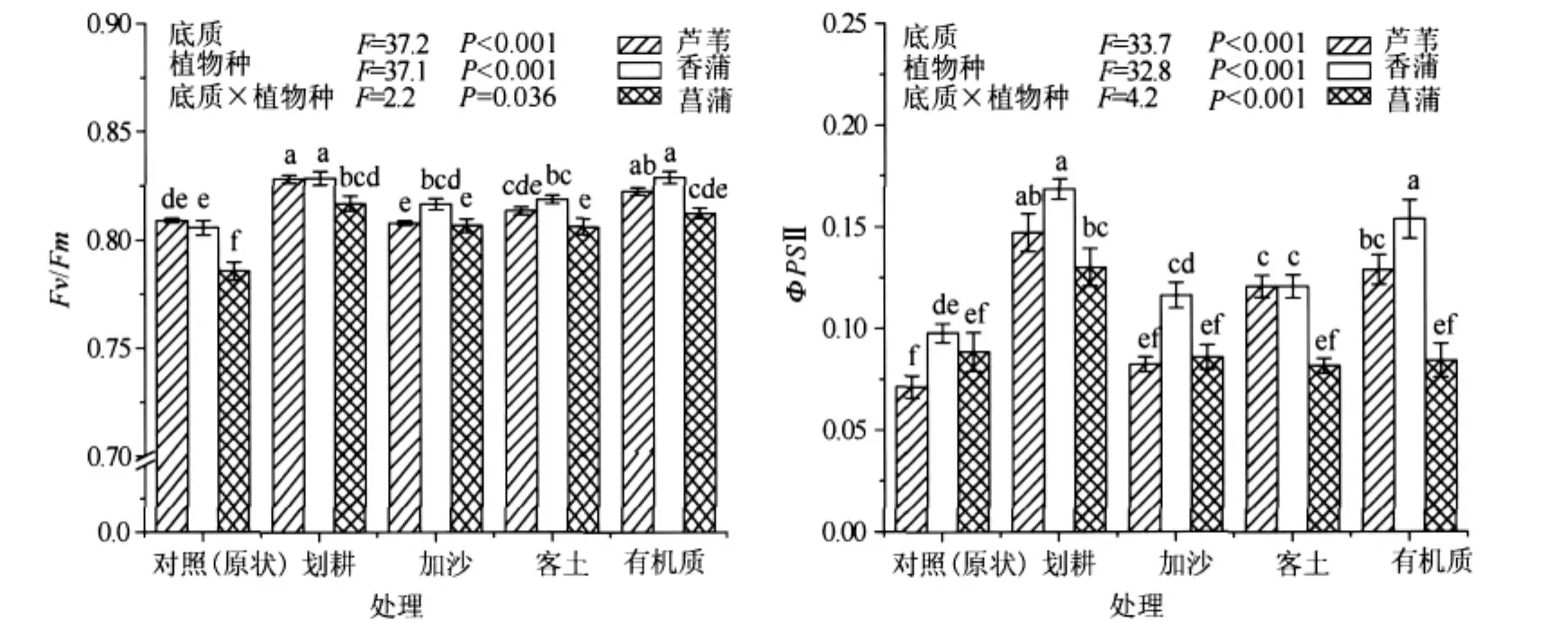
图3 不同底质对三种挺水植物Fv/Fm和ΦPSⅡ的影响Fig.3 Effects of sediment types on Fv/Fm and ΦPS Ⅱ in three emergent plants
2.4 不同底质对气体交换的影响
在三种植物中,香蒲的净光合速率和气孔导度最大,芦苇次之,而菖蒲则明显低于前两者(图4).同样,香蒲的胞间CO2浓度和蒸腾速率均显著大于芦苇和菖蒲,菖蒲的蒸腾速率显著大于芦苇,而胞间CO2浓度的差异则因底质处理方式不同而不同,菖蒲的胞间CO2浓度在原状、加沙和客土处理下显著小于芦苇,在划耕处理下显著大于芦苇,而在有机质处理下二者没有显著差异(图4).
除加沙处理下芦苇和香蒲净光合速率与对照(原状)的差异不显著以外,其它底质处理下三种植物的净光合速率均显著高于对照(图4).与对照(原状)相比,不同底质处理均显著提高了芦苇和菖蒲的气孔导度,但香蒲的气孔导度仅在划耕处理下显著高于对照(原状).
香蒲的胞间CO2浓度仅在有机质处理下显著小于对照(原状),其它处理与对照没有显著差异;芦苇的胞间CO2浓度在加沙处理下高于对照,在划耕和有机质处理下低于对照,在客土处理下与对照的差异不显著;香蒲胞间CO2浓度在划耕和有机质处理下高于对照,在客土和加沙处理下与对照的差异不显著(图4).
除加沙处理以外,其它三种底质处理均显著降低香蒲叶片的蒸腾速率,其中以有机质处理效果最佳.相反,四种处理却显著提高了菖蒲的蒸腾速率.芦苇的蒸腾速率在加沙和客土处理下也显著增大,但在划耕和有机质处理下与对照(原状)差异不显著(图4).
方差分析表明,底质和植物种对净光合速率、气孔导度、胞间CO2浓度和蒸腾速率均具有显著交互作用(P <0.01)(图4).
3 讨论
3.1 三种挺水植物光合生理特性的差异
本文研究结果表明,三种挺水植物地上部生物量以香蒲最大,芦苇次之,菖蒲最小.相关性分析表明,地上部生物量的变化与光合速率呈显著正相关(r=0.944,P<0.01).因此,可以认为光合速率的差异是三种挺水植物生长特性差异的重要原因.由于香蒲叶宽,含纤维量高,韧性强,香蒲全草为良好的造纸原料,同时可以用来编制草袋、草包、草席等,具有较高的经济价值和药用价值[19],加之具有适应性强,生长迅速,生物量大等特点,可作为构建巢湖流域人工湿地的首选挺水植物.

图4 不同底质对三种挺水植物气体交换的影响Fig.4 Effects of sediment types on gas exchange in three emergent plants
通常认为,光合速率主要受气孔限制和非气孔限制两个因素的影响[20].Farquhar等[21]认为,可用胞间CO2浓度判断气孔导度是否是光合速率相应变化的主要原因.只有当Ci与Pn变化方向相同,两者都减小时,才可认为光合速率的下降主要是由于气孔导度的降低引起的,否则,如果Pn下降,Ci升高,便可以肯定光合速率下降的决定因素是叶肉细胞光合活性的降低,而不是气孔导度的因素[22].本文研究结果表明,光合速率与气孔导度(r=0.774,P <0.01)、胞间 CO2浓度(r=0.571,P <0.05)均呈显著正相关关系.表明净光合速率受气孔导度的影响较大,气孔行为对其光合作用碳的固定显示了明显的主导控制作用.香蒲的气孔导度、胞间CO2浓度、蒸腾速率均显著大于芦苇和菖蒲,由此可见,较高的气孔导度和胞间CO2浓度,是香蒲具有较高光合速率和生物量的重要原因.
光合色素是植物叶片光合作用的主要物质基础,其含量的变化,既可反映植物叶片光合作用功能的强弱,也可作为衡量植物对生境条件适应性的一个重要指标.本文研究结果表明,香蒲叶片的Chl.a、Chl.b和Car含量均显著高于芦苇和菖蒲,这一结果与净光合速率的种间差异相一致.菖蒲叶片Chl.a/Chl.b显著小于香蒲和芦苇,而在香蒲和芦苇之间的差异不显著.叶片主要通过叶绿素等来捕捉光能进行光合作用.叶绿素含量多少反映了叶片的光合能力的强弱.Chl.a/Chl.b可以反映捕光色素复合体Ⅱ(LHCⅡ)在所有含叶绿素的结构中所占的比重,其值升高表明LHCⅡ含量的减少[23].类胡萝卜素既是光合色素,又是内源抗氧化剂,它可以耗散过剩光能,清除活性氧,从而防止膜脂过氧化,保护光合机构[24].因此,香蒲较高的叶绿素含量、Car含量以及Chl.a/Chl.b既有利于叶片对光能的捕获和利用效率,同时又能对光合机构起到保护作用.
光化学效率的高低直接决定叶片光合作用的高低,因而是光合作用的重要限制因子[25].正常情况下,植物叶片叶绿素吸收的光能主要通过光合电子传递、叶绿素荧光发射和热耗散3种途径来消耗,这3种途径之间存在着此消彼长的关系,光合作用和热耗散的变化会引起荧光的相应变化.因此,可以通过对荧光的观测来探究植物光合作用和热耗散的情况[26].Fv/Fm是暗适应条件下PSⅡ的最大光化学效率,反映了PSⅡ反应中心最大光能转化效率[27].ΦPSⅡ表示PSⅡ反应中心部分关闭情况下的实际光化学效率,反映植物叶片在光下用于电子传递的能量占吸收光能的比例[28].在三种挺水植物中,香蒲的Fv/Fm和ΦPSⅡ均最大,而菖蒲的Fv/Fm和ΦPSⅡ最小.说明香蒲的光合机构更有利于把所捕获的光能以更高的速度和效率转化为化学能,从而为碳同化提供更充足的能量,这也是其光合速率较高的重要原因.
3.2 不同底质对植物光合生理特性的影响
底质是水生植物生根、繁殖及稳定生长的基本条件,同时也是植物养分的直接来源之一[29].许多研究表明,随着底质营养的增加,沉水植物的生长加快[30-32],但Madsen等[33]发现底质营养对植物相对生长率的影响不大.除底质营养以外,底质结构也可能影响沉水植物的生长[34].然而,关于底质对挺水植物的影响,迄今尚不清楚.
挺水植物不同于沉水植物,其叶片大部分暴露在空气中.一方面由于植物的大部分叶片处于强光环境,因此叶片的光合特性表现出典型的阳生植物特征[17-18];另一方面在很大程度上降低了叶片从水体吸收矿质营养的可能性,因而受底质结构和营养状况的影响更大.本研究对底质进行改良的主要目的在于改变其紧实度,提高了土壤的通气性,从而为植物的生长提供较好的生境条件.但不同处理在改变底质紧实度的同时,又不可避免地会影响到其它因子,并进一步影响到植物的生长.总体上,四种底质改良方式(划耕、有机质、客土和加沙)均能不同程度地提高植物叶片的光合色素含量(Chl.a、Chl.b、Car和 Chl.a/Chl.b)和光化学反应中心活性(Fv/Fm和ΦPSⅡ),增加气孔导度和胞间CO2浓度,从而提高光合速率并最终促进植物的生长.
然而,不同底质处理下植物的生长和光合特性存在显著差异,其中以划耕和有机质处理对植物生长和光合特性的促进作用最为明显.其原因可能是划耕只改变底质的紧实度,提高了土壤的通气性,但不改变土壤的营养状况及其它理化特性;添加有机质既提高了土壤的通气性,又增加了有机质含量,改善了营养状况.由于划耕和有机质处理都保留了本土底质,而土著种由于长期生长在这种底质上,对本土底质最为适应,因而有利于植物的生长.客土处理在增加通气性的同时,在很大程度上改变了底质的理化特性,由于三种挺水植物均为土著种,对客土的适应性相对较差,从而制约了植物的生长;加沙处理在改变以底质的紧实度的同时,降低了底质营养,同样不利于植物的生长.因此,在构建湿地生态系统时,可以采用划耕和添加有机质等底质改良措施促进植物的生长,加速湿地植被构建的进程.
[1]郑焕春,周 青.微生物在富营养化水体生物修复中的作用.中国生态农业学报,2009,17:197-202.
[2]胡春华,濮培民,王国祥等.冬季净化湖水的效果与机理.中国环境科学,1999,19:561-565.
[3]Schnoor JL,Light LA,McCutcheon SC et al.Phytoremediation of organic and nutrient contaminants.Environmental Science and Technology,1995,29:318-323.
[4]Reuter JE,Djohan T,Goldman CR.The use of wetlands for nutrient removal from surface runoff in a cold climate region of california-results from a newly constructed wetland at lake tahoe.Journal of Environmental Management,1992,36:35-53.
[5]童昌华,杨肖娥,濮培民.富营养化水体的水生植物净化试验研究.应用生态学报,2004,15:1447-1450.
[6]Greenway M.Suitability of macrophytes for nutrient removal from surface flow constructed wetlands receiving secondary treated sewage effluent in Queensland,Australia.Water Science and Technology,2003,48:121-128.
[7]吴建强,阮晓红,王 雪.人工湿地中水生植物的作用和选择.水资源保护,2005,21:1-6.
[8]白峰青,郑丙辉,田自强.水生植物在水污染控制中的生态效应.环境科学与技术,2004,27:99-100.
[9]胡绵好,袁菊红,张 玲等.不同品种黑麦草对富营养化水体净化能力的比较.环境科学学报,2009:1740-1749.
[10]袁东海,高士祥,任全进等.几种挺水植物净化生活污水总氮和总磷效果的研究.水土保持学报,2004,18:77-80.
[11]Brix H.Treatment of wastewater in the rhizosphere of wetland plants-the root-zone method.Water Science and Technology,1987,19:107-118.
[12]Fennessy MS,Cronk JK,Mitsch WJ.Macrophyte productivity and community development in created freshwater wetlands under experimental hydrological conditions.Ecological Engineering,1994,3:469-484.
[13]雒维国,王世和,黄 娟等.植物光合及蒸腾特性对湿地脱氮效果的影响.中国环境科学,2006,26:30-33.
[14]黄 娟,王世和,雒维国等.人工湿地污水处理系统植物光合作用特性的研究.安全与环境工程,2006,13:55-57.
[15]黄 娟,王世和,雒维国等.植物光合特性及其对湿地 DO分布,净化效果的影响.环境科学学报,2006,26:1828-1832.
[16]卢晓明.植物净化槽处理城市黑臭河水的效果,机理及工程示范[学位论文].上海:华东师范大学,2009.
[17]付为国,李萍萍,卞新民等.镇江北固山湿地芦苇光合日变化的研究.西北植物学报,2006,26:496-501.
[18]李林锋.湿地植物香蒲光合特性及其影响因素分析.广东海洋大学学报,2009,29:77-82.
[19]赵丽萍,许 卉.野生植物资源香蒲的综合开发与利用.中国林副特产,2007:80-81.
[20]Mediavilla S,Santiago H,Escudero A.Stomatal and mesophyll limitations to photosynthesis in one evergreen and one deciduous Mediterranean oak species.Photosynthetica,2002,40:553-559.
[21]Farquhar GD,Sharkey TD.Stomatal conductance and photosynthesis.Annual Review of Plant Physiology,1982,33:317-345.
[22]Chartzoulakis K,Patakas A,Kofidis G et al.Water stress affects leaf anatomy,gas exchange,water relations and growth of two avocado cultivars.Scientia Horticulturae,2002,95:39-50.
[23]Anderson JM,Aro EM.Grana stacking and protection of photosystem Ⅱ in thylakoid membranes of higher plant leaves under sustained high irradiance:an hypothesis.Photosynthesis Research,1994,41:315-326.
[24]米海莉,许 兴,李树华等.水分胁迫对牛心朴子,甘草叶片色素,可溶性糖,淀粉含量及碳氮比的影响.西北植物学报,2004,24:1816-1821.
[25]韩瑞宏,卢欣石,高桂娟等.紫花苜蓿(Medicago sativa)对干旱胁迫的光合生理响应.生态学报,2007,27:5229-5237.
[26]Rohácˇek K.Chlorophyll fluorescence parameters:the definitions,photosynthetic meaning,and mutual relationships.Photosynthetica,2002,40:13-29.
[27]Krause GH,Weis E.Chlorophyll fluorescence and photosynthesis:the basics.Annual Review of Plant Biology,1991,42:313-349.
[28]孙宪芝,郑成淑,王秀峰.高温胁迫对切花菊‘神马’光合作用与叶绿素荧光的影响.应用生态学报,2008,19:2149-2154.
[29]翟水晶,胡维平,邓建才等.不同水深和底质对太湖马来眼子菜(Potamogeton malaianus)生长的影响.生态学报,2008,28:3035-3041.
[30]Xie Y,An S,Wu B et al.Density-dependent root morphology and root distribution in the submerged plant Vallisneria natans.Environmental and Experimental Botany,2006,57:195-200.
[31]Jiang J,Zhou C,An S et al.Sediment type,population density and their combined effect greatly charge the short-time growth of two common submerged macrophytes.Ecological Engineering,2008,34:79-90.
[32]Crossley M,Dennison W,Williams R et al.The interaction of water flow and nutrients on aquatic plant growth.Hydrobiologia,2002,489:63-70.
[33]Madsen T,Cedergreen N.Sources of nutrients to rooted submerged macrophytes growing in a nutrient-rich stream.Freshwater Biology,2002,47:283-291.
[34]Lauridsen T,Jeppesen E,Andersen F.Colonization of submerged macrophytes in shallow fish manipulated Lake Væng:impact of sediment composition and waterfowl grazing.Aquatic Botany,1993,46:1-15.
Effects of different sediment materials on photosynthetic characters of three emergent plant species
BAO Xianming1,FAN Chengxin2&SHI Gangrong1
(1:College of Life Sciences,Huaibei Normal University,Huaibei 235000,P.R.China)
(2:Nanjing Institute of Geography and Limnology,Chinese Academy of Sciences,Nanjing 210008,P.R.China)
To investigate the effects of sediment types on growth and photosynthetic characteristics in emergent plants,the leaf pigment contents,chlorophyll fluorescence and gas exchange of three dominant species(Phragmites communis,Typha minina and Acorus calamus)from aquatic vegetation of Lake Chaohu,which were grown in five different sediment types,were examined.The results showed that in the three emergent plant species,the shoot biomass was highest in T.minina,followed by P.communis,and the lowest in A.calamus.The chlorophyll-a content(Chl.a),chlorophyll-b content(Chl.b),carotenoid content(Car),stomatal conductance,intercellular CO2concentration,transpiration rate in T.minina were significantly higher than that in A.calamus and in P.communis,net photosynthetic rate,Fv/Fm and ΦPS Ⅱ in T.minina were significantly higher than that in A.calamus.According to correlation analysis,the shoot biomass was positively correlated to net photosynthetic rate,while net photosynthetic rate was significantly correlated to stomatal conductance and intercellular CO2concentration.Therefore,the results indicated that the high stomatal conductance,pigment content,as well as Fv/Fm and ΦPS Ⅱ in T.minina plants might lead to the high shoot biomass and net photosynthetic rate.Compared to the control(the original sediment),using the four sediment treatments(loosen sediments,organic sediments coverage,soil sediments coverage and sand sediments coverage)in varying degrees have increased pigment contents(Chl.a,Chl.b,Car and Chl.a/Chl.b),improved the activity of reaction center(Fv/Fm and ΦPS Ⅱ),and enhanced stomatal conductance and intercellular CO2concentration.As a result,the sediments enhanced photosynthetic rate and plant growth,and the loosen sediments and organic sediments coverage had high effects among the four treatments.
Emergent plants;sediment;gas exchange;pigment contents;chlorophyll fluorescence
* 国家水体污染控制与治理科技重大专项(2008ZX07103-003)和国家自然科学基金项目(50979102)联合资助.2010-08-03收稿;2010-11-22收修改稿.包先明,男,1978年生,博士,副教授;E-mail:xmbao96@126.com.
