小窝蛋白-1反义寡核苷酸在AVP诱导大鼠心肌成纤维细胞增殖erk1/2信号转导中的作用及辛伐他汀的干预效应
2011-08-02何燕萍杨军岭赵连友刘少伟解放军三二三医院陕西西安70054
何燕萍 杨军岭 赵连友 刘少伟 (解放军三二三医院,陕西 西安 70054)
近年来,小窝和小窝蛋白的细胞生理作用日益受到人们的重视,尤其是在信号转导、胆固醇运输及肿瘤抑制等方面有着重要的意义〔1〕。目前一些研究也提示,小窝结构的标志蛋白是小窝蛋白,可能与高血压的病理病生改变有关。文献报道,小窝蛋白可调节细胞外基质重构和纤连蛋白基质的更新,从而参与高血压的细胞外基质重构和组织修复病理病生过程〔2〕。并已证实,血管加压素(AVP)在高血压左室肥厚发生和发展中起重要作用〔3〕,心肌成纤维细胞(CFs)上有小窝蛋白-1大量表达〔4〕。AVP介导CFs增殖作用的信号转导如何在小窝上实现,目前尚不清楚。研究还发现,适当的胆固醇水平对维持小窝结构十分重要,胆固醇与小窝蛋白的结合能稳定小窝蛋白寡聚体,从而协同小窝蛋白介导的信号转导〔5〕。他汀类药是目前具有广泛心血管保护作用的以降低胆固醇为主的降脂药物,其抑制CFs增殖的信号转导机制是否与其降胆固醇作用并进而与影响小窝结构有关,目前还不十分明确。本研究设计合成小窝蛋白-1 AS ODN片段,旨在通过这种特异性抑制靶基因表达的细胞生物学研究手段,探讨小窝蛋白-1在CFs增殖erk1/2信号转导中的作用,以及辛伐他汀干预效应与小窝蛋白-1介导erk1/2信号转导的关系,为他汀类防治高血压心脏间质重构的发病机制提供新认识,也为他汀类药物防治心脏重构提供新的理论依据。
1 材料与方法
1.1 材料 雄性SD大鼠,体重200~250 g,6~8周龄,由第四军医大学实验动物中心提供。AVP、甲基-β-环糊精(MβCD)、黄体酮(Pro)、胆固醇、erk1/2抗体、p-erk 1/2抗体、p21抗体、cyclinA抗体、cav 1抗体为Sigma公司产品,Western印迹荧光检测试剂盒购自Santa Cruz公司,辛伐他汀纯品由湖北丝宝药业有限公司惠赠。
1.2 方法
1.2.1 心肌成纤维细胞(CFs)的培养和鉴定 腹腔麻醉(戊巴比妥50 mg/kg)6~8周龄SD大鼠后,无菌条件下迅速取出大鼠心脏,修剪去除心房和血管,仅留取心室部分。剪碎,PBS冲洗3次,加0.06%胶原酶Ⅱ和0.2%胰酶于37℃消化10~15 min,分 3次消化完毕。消化液经 100目滤网过滤,1 000 r/min离心共3次,每次用PBS清洗悬浮。所得细胞置于25 mm3培养瓶,100 ml/L胎牛血清的DMEM 100 ml培养瓶中,在37℃、50 ml/L CO2及饱和湿度下培养100 min,采用差速贴壁法分离CFs,待细胞生长近融合时,采用1∶3传代。在倒置显微镜、透射电镜下观察培养的细胞呈梭形、多角形,细胞质透明,细胞核明显变大,呈椭圆形,常含有2~3个核。经免疫组化纤维粘连蛋白染色阳性和平滑肌肌动蛋白染色阴性鉴定为所需的CFs,纯度达0.98,锥虫蓝染色细胞活力大于0.97。实验采用3~4代细胞。
1.2.2 cav1反义寡核苷酸设计及处理 根据大鼠cav1的脚手架域cDNA序列设计反义、正义及错配寡核苷酸,序列运用Blast程序在GenBank数据库进行同源性检索,无同源序列。由上海生工有限公司合成,聚丙烯酰胺凝胶电泳纯化,寡核苷酸分子两端碱基均行硫代修饰,反义寡核苷酸序列如下〔6〕:5'-AAA CTG TGT GTC CCT TCT GG-3',所有碱基均以硫代磷酸基修饰,并以该序列的错义寡核苷酸为对照(错义序列5'-AAA CAG TCT GAC CGT TGT GG-3')。将cav1-AS ODN用无血清DMEM稀释后加样,终浓度为5×10-5mol/L。用脂质体导入法将cav1-AS ODN导入CFs,孵育24 h后,检测CFs的DNA合成,及p-erk1/2和cav1表达。
1.2.3 CFs的DNA合成功能测定 以3H-TdR掺入率反映CFs的DNA合成功能。取对数生长期CFs,0.25%胰酶消化,用含10%胎牛血清的DMEM调整细胞悬液浓度为1×104/ml,加入96孔培养板中,每孔加入200 μl,静置培养至细胞接近融合。弃上清,加入无血清DMEM,继续培养24 h,使细胞维持生长静止状态。分别加入不同试剂干预。在最后的干预药物加入时,同时加入3H-TdR 7.4×104Bq/ml,12 h后吸弃上清液,二蒸水冲洗,加10%三氯乙酸洗5 min,加0.3 mol/L NaOH处理30 min,多头细胞收集器收集细胞裂解液至玻璃纤维滤纸上,烤干。将玻璃纤维滤纸置于闪烁计数管中,每管加入闪烁液0.5 ml,液态闪烁计数仪测定放射性,结果以cpm值表示。
1.2.4 蛋白免疫印迹分析 收集不同条件处理的CFs,全细胞裂解液裂解细胞,4 000 r/min离心去除沉淀,BCA试剂测定蛋白含量。1×SDS凝胶加样缓冲液调蛋白浓度使各组一致,用10%SDS-聚丙烯酰胺凝胶电泳分离。收集CFs与等体积2×电泳加样缓冲液混合后,煮沸5 min,每泳道上样10 μl。采用蛋白提取液提取总蛋白,紫外分光光度计检测蛋白含量。灌制15%的分离胶和4%积层胶,取每组细胞100 μg(20 μl)蛋白加入上样孔,90 V恒压垂直电泳2 h后,取出凝胶,在BioRad转移槽中28 mA恒流湿转18 h后,取出PVDF膜进行抗原抗体反应(cav1一抗稀释浓度为1∶1 000,二抗为1∶3 000)。为观察蛋白质转移情况,对硝酸纤维素膜进行丽春红S染色,观察完毕后用去离子水将染液漂洗干净,DAB显色,待显色适度时,用水漂洗终止显色反应,凝胶成像系统扫描,软件分析计算A值。erk 1/2抗体、p-erk1/2抗体、p21抗体、cyclin A抗体免疫印迹步骤同上,抗体浓度参照试剂说明书。
1.2.5 实验分组 (1)空白对照组:无血清DMEM培养液;(2)AVP组;(3)Sim组;(4)cav1反义链单独作用组;(5)cav1错义链单独作用组;(6)cav1反义链+AVP组;(7)cav1错配链+AVP组;(8)MβCD 组;(9)Pro组;(10)15 μg/ml胆固醇组;(11)Pro+15 μg/ml胆固醇组;(12)MβCD+15 μg/ml胆固醇组;(13)Sim+胆固醇组:又分为Sim+10 μg/ml胆固醇组亚组、Sim+15 μg/ml胆固醇组亚组、Sim+20 μg/ml胆固醇组亚组;用DMEM调整AVP终浓度为1×10-7mol/L,Sim终浓度为10-7mol/L,cav1-AS ODN 终浓度为 5 μmol/L。每组设6~8个复孔。
2 结果
2.1 cav1反义寡核苷酸对大鼠CFs的DNA合成的影响cav1反义和错义寡核苷酸分别导入大鼠CFs并孵育24 h后,大鼠CFs内3H-TdR掺入率分别相当于空白对照组(100±5)%的(157±7)%和(95±5)%,其中反义寡核苷酸组与空白对照组比较,差异非常显著(P<0.01)。错义寡核苷酸组与空白对照组比较无统计学差异(P>0.05)。在10-7mol/L的AVP单独干预CFs 24 h后,大鼠CFs内3H-TdR掺入率相当于空白对照组的(172±4)%,与空白对照组相比,差异非常显著(P<0.01)。将 cav1反义或错义寡核苷酸分别与10-7mol/L的AVP共同干预CFs 24 h后,大鼠CFs内3H-TdR酸掺入率分别相当于空白对照组的(212±6)%和(170±3)%,其中反义寡核苷酸+10-7mol/L的AVP共同干预组与10-7mol/L的AVP单独干预组比较,差异非常显著(P<0.01)。错义寡核苷酸+10-7mol/L的AVP共同干预组与10-7mol/L的AVP单独干预组比较无统计学差异(P>0.05)。
2.2 cav1反义寡核苷酸对大鼠CFs内erk1/2 MAPK活性的影响 cav1反义和错义寡核苷酸分别干预大鼠CFs 5 min后,大鼠CFs内p-erk1/2蛋白表达量分别相当于空白对照组的(2.3±0.3)倍和(1.1±0.2)倍,其中反义寡核苷酸组与空白对照组比较,差异非常显著(P<0.01)。错义寡核苷酸组与空白对照组比较无差异(P>0.05)。10-7mol/L的AVP单药干预CFs 5 min后,CFs内p-erk1/2蛋白表达量相当于空白对照组的(5.7±0.2)倍,与空白对照组相比,差异非常显著(P<0.01)。将cav1反义或错义寡核苷酸分别与10-7mol/L的AVP共同干预CFs 5 min后,大鼠CFs内p-erk1/2蛋白表达量分别相当于空白对照组的(7.9±0.3)倍和(5.5±0.2)倍,其中反义寡核苷酸+10-7mol/L的AVP共同干预组与10-7mol/L的AVP单独干预组比较,差异非常显著(P<0.01)。错义寡核苷酸+10-7mol/L的AVP共同干预组与10-7mol/LAVP单独干预组比较无差异(P>0.05)。见图1。
2.3 cav1反义寡核苷酸对大鼠CFs内p21和细胞周期蛋白A蛋白表达的影响 在用cav1反义寡核苷酸单独干预或者与10-7mol/L的 AVP共同干预大鼠CFs 24 h后,两组CFs内p21蛋白表达强度分别较对照组下降,细胞周期蛋白A表达强度升高。见图2。
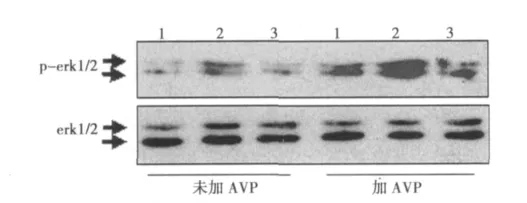
图1 cav1反义寡核苷酸对大鼠CFs内erk1/2 MAPK活性的影响
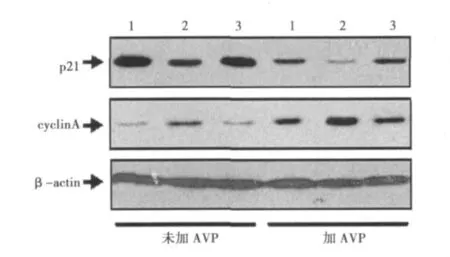
图2 cav1反义寡核苷酸对大鼠CFs内p21和细胞周期蛋白A蛋白表达的影响
2.4 胆固醇对大鼠CFs胞膜上cav1蛋白表达的影响 用2%的MβCD或10 μg/ml Pro或10-7mol/L Sim干预大鼠CFs 24 h后,CFs胞膜上cav1蛋白表达量分别相当于对照组(100±5)%的(61±3)%、(61±2)%和(66±4)%,与对照组比较,差异均非常显著(P<0.01)。
用15 μg/ml的胆固醇分别与2%的MβCD或10 μg/ml Pro共同干预CFs 24 h后,CFs胞膜上cav1蛋白表达量分别相当于对照组的 (86±2)%和 (86±3)%,与2%的MβCD或10 μg/ml Pro单独作用组比较,统计学差异均非常显著 (P<0.01)。用10、15或 20 μg/ml胆固醇分别与 10-7mol/L Sim共同干预CFs 24 h后,CFs胞膜上cav1蛋白表达量分别相当于对照组的 (86±3)%、 (91±4)%和 (94±5)%,其中10 μg/ml胆固醇和 10-7mol/L Sim 共同干预与 10-7mol/L Sim单独作用组比较,差异显著 (P<0.05),15 μg/ml或20 μg/ml胆固醇+10-7mol/L Sim共同干预组与10-7mol/L Sim单独作用组比较,差异均非常显著 (P<0.01)。15 μg/ml胆固醇单独干预CFs 24 h后,CFs胞膜上cav1蛋白表达量相当于对照组的 (119±5)%,与对照组比较,统计学差异显著 (P<0.05)。见图3。
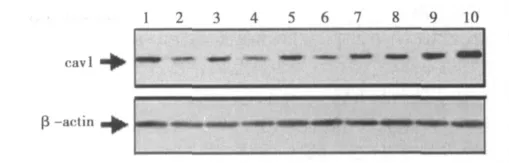
图3 胆固醇对cav1蛋白表达的影响
3 讨论
信号转导是近年来细胞生物学研究中的主要领域,心脏重构机制也与之关系十分密切。这种信号转导过程首先是在生物膜上进行的,而究竟是在生物膜的什么部位,目前还不十分清楚。研究证实,细胞膜上小窝结构是许多信号分子完成跨膜信号转导的“驿站”,小窝蛋白是小窝结构的标志性蛋白,可与信号分子直接接触,调节细胞的功能状态〔7〕。文献还报道,丝裂原激活的蛋白激酶(MAPK)中,erk1/2信号通路的上游蛋白Ras、Raf-1和 erk1/2本身定位于小窝区域〔8〕。研究已证实,AVP可通过其V1受体诱导新生大鼠CFs增殖〔9〕,从而具有一定的促纤维化作用。本研究结果表明,AVP可活化erk1/2,同时增强细胞周期蛋白A表达,并抑制细胞周期蛋白抑制因子p21的表达,从而丰富了对AVP促增殖作用机制的认识。本研究采用cav1-AS ODN导入细胞后,可与编码cav1的双链或单链DNA结合,抑制剪接或降解目的mRNA,阻断翻译起始过程,也就是作用于cav1遗传信息传递的各个步骤发挥其反义作用。研究表明,cav1-AS ODN对基础状态及AVP诱导的CFs增殖起促进作用,从而提示,升高cav1蛋白的表达水平可能会抑制AVP诱导的CFs增殖,进而发挥抗心肌纤维化作用。同时,小窝是细胞膜上富含胆固醇和鞘脂的膜微区,胆固醇对于维持小窝的完整性是必需的,是细胞内多种负性信号分子的锚定部位和信号分子对话(cross-talk)的重要场所,胆固醇对于维持小窝的完整及信号转导起着重要作用〔5〕。黄体酮也有类似作用,并且主要影响小窝内的胆固醇水平。本研究用β-甲基环糊精和黄体酮干扰了细胞膜上的胆固醇水平后,cav1的表达减少,影响了小窝形成,进而干扰了小窝内的信号转导。
目前关于他汀类药物抗纤维化机制研究正处于初步认识阶段,已经明确他汀类药物对血清胆固醇水平正常的高血压病患者有一定的降压作用,可间接改善心肌的肥厚〔10〕,一方面与降低内源性胆固醇水平有关,另一方面可能抑制甲羟戊酸及类异戊二烯的合成,影响小G蛋白的成熟、定位和激活,从而阻断MAPK途径的激活等作用有关。然而,已研究表明类异戊二烯化在细胞膜上的网格蛋白而不是小窝内完成〔11〕。本研究表明Sim通过降低细胞膜上胆固醇水平,减少了cav1的表达水平。在增加细胞膜上的胆固醇水平后,cav1表达增加,可与Sim共同对erk 1/2增殖信号起负性调控作用。因此,对血清胆固醇水平正常的高血压病患者在服用他汀类药物的同时,适当地增加胆固醇类物质的摄入可能有利于他汀类药物的抗心肌纤维化作用。
1 Liu P,Rudick M,Anderson RG.Multiple functions of caveolin-1〔J〕.J Biol Chem,2002;277(44):41295-8.
2 Sottile J,Chandler J.Fibronectin matrix turnover occurs through a caveolin-1 dependent process〔J〕.Mol Biol Cell,2005;16(2):757-68.
3 赵连友,陈永清.蛋白激酶C抑制剂对精氨酸升压素调控心脏成纤维细胞增殖及p27蛋白表达的影响〔J〕.中华医学杂志,2003;83(5):421-4.
4 Zhao YY,Liu Y,Stan RV,et al.Defects in caveolin-1 cause dilated cardiomyopathy and pulmonary hypertension in knockout mice〔J〕.Proc Natl Acad Sci,2002;99(17):11375-80.
5 Rothblat GH,de la Llera-Moya M,Atger V,et al.Cell cholesterol efflux:integration of old and new observations provides new insights〔J〕.J Lipid Res,1999;40(5):781-96.
6 Rothberg KG,Heuser JE,Donzell WC,et al.Caveolin,a protein component of caveolae membrane coats〔J〕.Cell,1992;68(4):673-82.
7 Gratton JP,Bernatchez P,Sessa WC.Caveolae and caveolins in the cardiovascular system〔J〕.Cir Res,2004;94(11):1408-17.
8 Liu P,Ames GF.Platelet-derived growth factor activates mitogen-activated protein kinase in isolated caveolae〔J〕.Proc Natl Acad Sci,1997;94(25):13666-70.
9 Yang XD,Zhao LY,Zheng QS,et al.Effects of arginine vasopressin on growth of rat cardiac fibroblasts:role of V1 receptor〔J〕.J Cardiovasc Pharmacol,2003;42(1):132-5.
10 Masao T,Koichi N,Hironori N.Statins as antioxidant therapy for preventing cardiac myocyte hypertrophy〔J〕.J Clin Invest,2001;108:1429-37.
11 Laufs U,Marra D,Node K,et al.3-Hydroxy-3-methylglutaryl-CoA reductase inhibitors attenuate vascular smooth muscle proliferation by preventing Rho GTPase-induced down-regulation of p27(Kip1)〔J〕.J Biol Chem.1999;30;274(31):21926-31.
