Akt对哮喘小鼠肺组织TNF-α的调节作用
2011-05-25张宝辉马腾刘晓湘方秀斌
张宝辉,马腾,刘晓湘,方秀斌
(中国医科大学基础医学院神经生物学教研室,沈阳 110001)
支气管哮喘(简称哮喘)为一由非特异性炎症引起的以气道高反应性和气道炎症为主要特征的慢性炎症疾病。其发病机制比较复杂,许多炎性细胞、因子、介质等均参与其发病过程。研究证明,神经生长因子(nerve growth factor,NGF)通过作用于炎性细胞、作用于神经元,参与哮喘时的气道炎性反应、气道收缩和气道高反应性的,最近有研究报道,Akt/PKB参与了哮喘发病机制中NGF介导的信号传导通路,本人以往的实验也证明Akt/PBK能上调肺及C7-T5段脊神经节及相应脊髓后角内IL-1β的表达[1],也能上调肺组织内血管内皮细胞生长因子(vascular endothelial growth factor,VEGF)的表达[2],说明Akt/PBK很可能通过调节细胞因子IL-1β和VEGF的表达来参与NGF介导的哮喘发病机制。有研究证明,TNF-α通过激发细胞因子网络造成细胞因子的级联反应,进而诱生其他的炎性介质从不同的环节参与炎性反应,引起器官组织损伤,是过敏性支气管哮喘中的一个重要启动因子[3]。但是Akt/PBK能否通过调节TNF-α分子的表达参与哮喘仍未见文献报道,本研究利用免疫荧光、Western blot方法检测TNF-α的表达来了解Akt/PBK能否通过调节TNF-α分子的表达参与哮喘。
1 材料与方法
1.1 实验动物分组及模型制备
雌性BALB/c小鼠30只,由首都医科大学实验动物中心提供。按随机数字表法分为:(1)哮喘组10只,第1天每只小鼠腹腔注射20μg OVA(Sigma公司)和2 mg氢氧化铝混合于0.5 ml PBS中,第8天、15天每只小鼠腹腔注射10μg OVA和1 mg氢氧化铝混合于0.5 ml PBS中,第16天开始将小鼠置于封闭容器中,给予4%OVA(PBS配成)雾化吸入激发哮喘发作,每天 1 次,每次 25 min,连续 7 d;(2)Akt/PKB阻断组,第19天开始雾化吸入前3 h鼻腔给 Akt/PKB 阻断剂-LY294002(0.3 mg/kg),其余均与哮喘组相同;(3)正常对照组,以PBS代替OVA注射和雾化吸入,其余均与哮喘组相同。各组最后一次激发后24 h处死。
1.2 方法
1.2.1 AniRes2005肺功能仪测气道反应性:0.4%的戊巴比妥钠腹腔麻醉小鼠(80 mg/kg),疼痛反射消失后固定在小鼠体描箱内操作台上,气管插管,游离颈外静脉,行静脉穿刺,固定针柄,封闭体描箱。乙酰甲胆碱(mAch)用磷酸缓冲液(0.1 mol/L Ph 7.4)配成 0.025、0.05、0.075、0.1 mg/kg 经颈外静脉各注射0.1 ml记录波形和数据。
1.2.2 免疫荧光:将肺组织以恒冷箱切片机连续切片(片厚8μm),冷风干燥后,室温下血清封闭20 min,甩干,滴加一抗TNF-α(购自武汉博士德生物公司),4℃孵育过夜,PBS充分洗涤,滴加二抗(购自美国Sant Crutz公司),孵育40 min,PBS充分洗涤,甘油封片,选取3组动物的左肺相同部位的8张切片,每张切片选取六个区域,用OlympusBX51型荧光显微镜进行观察。绿色荧光为阳性表达。
1.2.3 Western blot免疫蛋白印记:计算加样量,加入样品缓冲液,沸水浴中加热5 min使蛋白质变性,制备12%分离胶,4%浓缩胶,将样品加在凝胶表面样品槽中。电泳后湿转至硝酸纤维素膜上。5%脱脂奶粉TTBS缓冲液4℃过夜。一抗TNF-α、β-actin(1∶200,)室温孵育2 h,洗膜,辣根酶标记羊抗兔IgG-HRP(1∶5000,Santa Cruz)室温孵育 1 h,洗膜后加入ECL(enhanced chemiluminescence)试剂(SantaCruz)反应1 min,暗室曝光显影后冲洗胶片。凝胶成像分析系统上摄像分析,测得目标带的IDV(integrated density value),进而计算出各组样品TNF-α目标带的IDV与内参照β-actin IDV的比值。
1.3 统计学分析
采用SPSS 13.0统计软件对数据进行统计分析,结果用x±s表示,组间比较进行t检验。检验水准 α=0.05。
2 结果
2.1 哮喘模型制作结果
正常对照组雾化吸入生理盐水后无明显哮喘反应,哮喘组动物在雾化吸入卵蛋白3次后,大部分哮喘发作,表现为呼吸频率加深加快,肋间隙凹陷,刺激性呛咳;Akt/PKB阻断组动物,前3次雾化吸入卵蛋白后,大部分哮喘发作,应用阻断剂后,诱喘反应不再明显。
2.2 AniRes2005肺功能仪测气道阻力
哮喘组小鼠对mAch的浓度反应曲线明显上移,其呼气阻力和吸气阻力显著高于正常(P<0.01),表明哮喘小鼠对mAch的气道反应性显著高于正常小鼠,表明哮喘模型建立成功。Akt/PKB阻断组对mAch的浓度反应曲线明显低于哮喘组(P<0.01),提示Akt/PKB阻断剂-LY294002对哮喘的气道高反应有阻断作用(图1,图2)。

图1 各组小鼠的吸气阻力Fig.1 Inspiratory resistance measured with A ni R es 2005 pulmonary function meter in each group
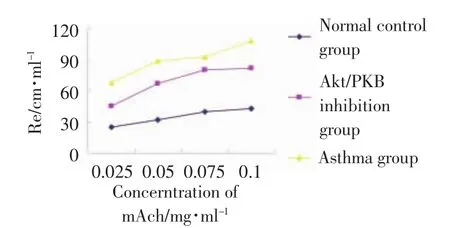
图2 各组小鼠的呼气阻力Fig.2 Expiratory resistance measured with A ni R es 2005 pulmonary function meter in each group
2.3 免疫荧光
从免疫荧光图片上可以看出,哮喘组小鼠的肺内炎性细胞、气道上皮(图3B)的TNF-α的阳性表达量显著高于正常对照组小鼠(图3A),而Akt/PKB阻断组(图3C)则明显低于哮喘组。而显微图像分析结果也表明,哮喘组TNF-α阳性反应产物MOD值明显高于对照组,Akt阻断组TNF-α阳性反应产物MOD值明显低于哮喘组(P<0.01,表1)。

图3 免疫荧光方法检测各组小鼠TN F-α蛋白的表达 ×200Fig.3 Expression of TN F-αprotein detected by immunofluorescence in each group ×200

表1 各组小鼠的TN F-α蛋白的光密度值比较(x±s)T ab.1 Mean optical density of TN F-α in each group(x ±s)
2.4 Western blot结果
哮喘组小鼠肺组织TNF-α蛋白的光密度值为3.82±0.24,与正常对照组 2.51±0.12相比,表达量明显升高(P<0.01);Akt/PKB阻断组TNF-α蛋白的光密度值为3.27±0.18,比哮喘组明显减少(P<0.01),说明经鼻腔滴入Akt/PKB阻断剂(LY-294002)效果比较确切。

图4 western blot方法检测各组小鼠TN F-α蛋白的表达Fig.4 Expression of TN F-αprotein detected by W estern blot in each group
3 讨论
本室以往的实验发现,NGF能够上调哮喘小鼠体内SH2-Bβ和Akt的表达,提示在NGF介导的哮喘发病机制中,NGF上调其下游底物SH2-Bβ和Akt的表达是它参与哮喘发病机制调节的途径之一[4,5],以往的研究证明 Akt/PBK 能上调肺及 C7-T5段脊神经节及相应脊髓后角内IL-1β的表达,也能上调肺组织内TNF-Α的表达,说明Akt/PBK很可能通过对细胞因子IL-1β和TNF-Α的表达来参与哮喘的发病机制。
Tan等[6]报道,重度过敏性支气管哮喘的TNF-α水平高于轻、中度患者,TNF-α水平越高,病情越重,也有研究证明TNF-α可通过转录因子(AP-1)而引起一些与炎性反应有关的细胞因子、受体、酶的基因表达,引起气道上皮细胞脱落,诱发或加重哮喘[7]。研究发现TNF-α单独作用不能诱导嗜酸性粒细胞趋化因子过多的释放与表达,但如果与IL-13、IL-4协同作用可以使嗜酸性粒细胞趋化因子的释放与表达是正常的20倍,表明TNF-α可显著增加IL-13对嗜酸性粒细胞趋化因子的刺激作用,引起嗜酸性粒细胞的聚集[8,9]。还有研究发现,对现有治疗不敏感的哮喘患者,其气道肿瘤坏死因子α(TNF-α)的表达增高,在过敏性哮喘的研究中也发现,慢性抗原暴露可导致气道TNF-α的产生。进一步的研究则证明,患者吸入TNF-α可增加气道的反应性,同时伴有嗜中性粒细胞性炎性反应[10]。Barnes等[11]研究发现,哮喘急性期与缓解期痰液、支气管肺泡灌流液或血清INF-α水平均明显高于健康对照者,说明炎症过程中TNF-α表达增加,在炎性细胞浸润、激活中起重要作用,但是Akt/PKB蛋白能否通过调节TNF-α参与哮喘发病机制的调节,尚未见文献报道。
在本实验中,从免疫荧光结果可以看出,与正常对照组相比,哮喘组、Akt/PKB阻断组小鼠肺内炎性细胞、气道上皮TNF-α的荧光表达量显著高于正常对照组小鼠,而Akt/PKB阻断组小鼠肺内炎性细胞、气道上皮的TNF-α的荧光表达量则明显低于哮喘组,而Western blot结果也证明,哮喘组、Akt/PKB阻断组小鼠肺组织内TNF-α蛋白表达量明显高于正常对照组,而Akt/PKB阻断组TNF-α蛋白表达量比哮喘组则明显减少,这结果提示Akt/PKB很可能是通过上调TNF-α的表达来参与NGF介导的哮喘发病机制。
本实验的结果提示利用Akt阻断剂直接或间接调节TNF-α的表达水平可能是治疗哮喘的新途径。
[1]张宝辉,刘玉丽,方秀斌,等.Akt对哮喘小鼠下呼吸道及内脏感觉传入部位 IL-1β 表达的影响[J].解剖科学进展,2007,13(4):289-292.
[2]张宝辉,刘玉丽,方秀斌,等.Akt对哮喘小鼠肺组织VEGF表达的上调作用[J].解剖科学进展,2008,14(1):39-42.
[3]Spielmann S,Kemer T,Ahlers O,et al.Early Detection of Increased Tumour Necrosis Factor Alpha (TNFalpha)and Soluble TNF Receptor Protein Plasma Levels after Trauma Reveal Associations with the Clinical Course[J].Anaesthesiol Scand,2001,45(3):364.
[4]刘玉丽,曹国龙,方秀斌,等.NGF对哮喘小鼠气道阻力和肺组织Akt/PKB 表达的影响[J].解剖科学进展,2006,12(4):304-306.
[5]LIUY,ZHANGB,FANGX,et al.NGFmediated SH2-bβ/Akt signal pathway activated in allergic airway challenge in mice[J].Respirology,2010,15(1):80-84.
[6]Tan EC,Lee BW,Tay AW,et al.Asthma and TNF Varianin in Chineseand Malys[J].Allergy Med,2007,54(4):402-403.
[7]Ciprandi G,Cirillo L.Mizolastine and Fexofenadine Modulat Cytokine Pattern after Nasal Allergen Challenge [J].Aller Immunol,2004,36(4):146-150.
[8]Moore PE,Church TL,Chism DD,et al.IL-13 and IL-4 Cause Eotaxin Release in Human Airway Smooth Muscle Cells:a Role for ERK[J].AmJPhysiol Lung Cell Mol Physiol,2002,282(4):847-853.
[9]丁宁,肖慧,高巨,等.细胞外调节蛋白激酶信号通路参与机械牵张诱导肺泡上皮细胞高迁移率族蛋白B1的表达调控[J].解剖学报,2009,40(6):919-922.
[10]Berry M,Brightling C,Pavord I,et al.TNF-alpha in asthma[J].Curr Opin Pharmacol,2007,7(3):279-282.
[11]Barnes PJ,Adcock M.How do eortieosteroids work in asthma[J]?Ann InternMed,2003,139(5 Pt1):3592-3701.
