于桥水库菹草过度生长对水质的影响及成因分析
2011-05-10陈孝军王立义高学平
张 晨,陈孝军,王立义,高学平
(1. 天津大学建筑工程学院,天津 300072;2. 天津市引滦工程于桥水库管理处,天津 301900;3. 天津市水利局引滦工程管理处,天津 301900)
近年来 的研究成果表明,沉水植物对富营养化水体水质具有一定净化效果[1-4].但是,一些草型湖泊、水库存在草型富营养化问题,在短时间内沉水植物过度生长使水体水质恶化,影响水利工程的正常运行,甚至危及人民饮水安全,如山东的南四湖、内蒙古的乌梁素海和哈素海、天津的于桥水库和尔王庄水库等都出现沉水植物疯长的现象.
国内关于草型富营养化的研究成果较少,尚士友等[5-6]利用沉水植物收割工程技术对草型湖泊进行治理,以机械化方式适度控制了乌梁素海草型富营养化;顾久君等[7]通过对乌梁素海菹草腐烂分解过程的研究表明,植物体内氮、磷的释放速率随水中营养盐浓度升高而增加.国外涉及草型富营养化的研究已取得了一定的成果.Miranda等[8]、Frodge等[9]揭示了草型富营养化浅水水库中沉水植物覆盖范围与水体溶解氧浓度之间的关系.Roman等[10]通过实验证实了草型富营养化湖泊水质恶化主要受沉水植物过度生长影响.Coops等[11]介绍了荷兰两座草型富营养化湖泊通过抑制沉水植物生长从而使湖泊水质得到改善,并证实改变水下光照条件能有效抑制沉水植物生长.
本文通过分析于桥水库草型富营养化特征,研究菹草过度生长对水库水质的影响,探究于桥水库菹草疯长发生成因,提出抑制菹草过度生长、防治草型富营养化的措施,为安全供水提供参考依据.
1 于桥水库草型富营养化特征
于桥水库位于天津市蓟县城东4,km、蓟运河左支流州河上游出山口处,北纬 40°02′,东经 117°25′,库区由东向西倾斜(见图1).作为引滦入津工程的调蓄水库,总库容15.59×108,m3,兴利库容为3.85×108m3,正常蓄水位21.16,m(大沽),正常蓄水位水面面积86.8,km2,平均水深4.5,m.引滦通水26年来,于桥水库水体营养状态已呈中度富营养水平[12],其中氨氮、总氮、总磷属水库无机污染物,1993—2006年的年均入库负荷分别为651.9,t、4,534.4,t和246.5,t;用高锰酸盐指数表征有机污染物,其年均入库负荷为2,965.6,t.
根据蓟县水产局 2006—2007年水生植物调查,于桥水库大型水生植物有 21种,分属于 14科 13属.按水生植物生态类型分类,有挺水植物 6种,浮叶植物2种,沉水植物13种.于桥水库大型水生植物的优势种为菹草(Potamogeton crispusLinn.)、狐尾藻(Myriophyllum spicatumLinn.)、眼子藻(Potamogeton malaianusMiq.)、荇菜(Nymphoides peltata(Gmel)O.Kuntze.)、野菱(Trapa inciseSieb. et Zucc.)和芦苇(Phragmites communisTrin.)等.

图1 于桥水库监测点分布状况Fig.1 Distribution of monitoring sections in Yuqiao ig. Reservoir
调查结果显示:2006年 5月菹草在库区为绝对优势种,约占水库总面积的 35%~61.5%,即约 30~53,km2,荇菜和野菱面积共约 8.3%,库区菹草植株数量约占总数的 80%,生物湿重分别为菹草 2.8~4.6,kg/m2、荇菜 0.5,kg/m2、野菱 0.3,kg/m2;6 月菹草开始衰败并腐烂,在河道内和岸边大量堆积,动用大量人力物力进行清除.2006年6月,荇菜、狐尾藻、穿叶眼子藻约占 3.25,km2,生物湿重分别为荇菜 0.6~3.4,kg/m2、狐尾藻 0.45~3.5,kg/m2、眼子藻 0.20~0.45,kg/m2.于桥水库草型富营养化特征显著,5—6月库区菹草超过水库面积的 30%,生物量大于2,kg/m2,属过度生长.库区水域均有菹草生长,水深在 2.0,m以下的浅水水域菹草密度较大,主河道中心水域密度较小,沿主河道向两侧呈条带状分布,距主河道越远分布密度越大.于桥水库菹草的发育历期基本一致,一年内,菹草生物量累计曲线呈单峰型,4月中旬至5月中旬为指数生长期,5月中旬至6月中旬为芽殖体发生期,该期生物量为高峰期.
2 材料与方法
参照《水库渔业资源调查规范》(SL 167—1996)中大型水生植物调查规范,选择密集区、一般区和稀疏区布设9个采样点,用GPS定位,如图1所示.2006年5月12日和6月28日进行了2次调查采样.
采用现场监测的方法记录于桥水库水位和各水质指标浓度.监测断面为库心、库北、库西、库东和放水洞.监测指标包括溶解氧 DO、pH 值、高锰酸盐指数CODMn、总氮TN、总磷TP、透明度和水温.太阳辐射强度数据来源于中国气象科学数据共享服务网.
用碘量法(GB 7489—87)测定溶解氧;用玻璃电极法(GB6 920—86)测定 pH 值;用酸性高锰酸钾氧化法(GB 11892—89)测定高锰酸盐指数;用碱性过硫酸钾消解紫外分光光度法(GB 11894—89)测定总氮;用钼酸铵分光光度法(GB 11893—89)测定总磷;用塞氏盘法(SL 87—1994)测定透明度.
3 菹草过度生长对水库水质的影响
3.1 对溶解氧和pH值的影响
过度生长的菹草占据了大量水体空间,生长旺盛期因光合作用强烈,出现水体溶解氧过饱和现象,同时菹草吸收水中大量CO2造成库区pH值骤然升高.
2006年5—6月于桥水库各监测断面DO和pH值见图2.由图2可知,5月中旬至6月上旬,菹草生长旺盛,于桥水库溶解氧急剧升高,由 6,mg/L升至8~9,mg/L,pH值亦急剧升高,由8.5左右升至 10左右;进入6月中旬,菹草开始衰败死亡,溶解氧逐渐降低,pH值也随之降至8.5.

图2 于桥水库各监测断面DO和pH值变化曲线Fig.2 Variations of DO and pH in each monitoring section in Yuqiao Reservoir
3.2 对高锰酸盐指数和营养盐的影响
尽管菹草在生长期摄取水体和底质中大量营养盐,对净化水质有一定作用,但其死亡后腐败分解,消耗溶解氧,释放出大量的氮和磷,造成菹草聚集区高锰酸盐指数和营养盐浓度升高,水体变黑发臭,鱼类大量死亡,水体二次污染.
2006年 5—6月于桥水库各监测断面 CODMn、TN和TP变化曲线如图3所示.由图3可知,CODMn浓度由 5月中旬的 3.5,mg/L上升至 6月中旬的5,mg/L左右,其原因为 6月中旬菹草进入植株衰亡期,在腐败分解过程中释放的有机物消耗溶解氧使CODMn浓度升高.6月初至 6月中旬,菹草衰亡后释放营养盐,水体营养盐负荷进一步加大,TN由2.22,mg/L升高至2.57,mg/L,TP由0.02,mg/L升高至0.06,mg/L,分别提高了 15.8%和 200%.6月 6日和 7日,由于大量死亡的菹草随水流运动至出水口,致使放水洞附近CODMn、TN和TP浓度剧然升高,分别至7.3,mg/L、2.89,mg/L和0.08,mg/L,短期内恶化供水水质,待完成打捞作业后,浓度分别恢复至 4,mg/L、2.10,mg/L和 0.03,mg/L,恶化的水质状况与打捞后相比,分别提高了 82.5%、37.6%和 167%.分析结果表明,水草衰亡释放的高营养盐和有机质对水质影响较大,时间自6月8日—13日,持续1周.
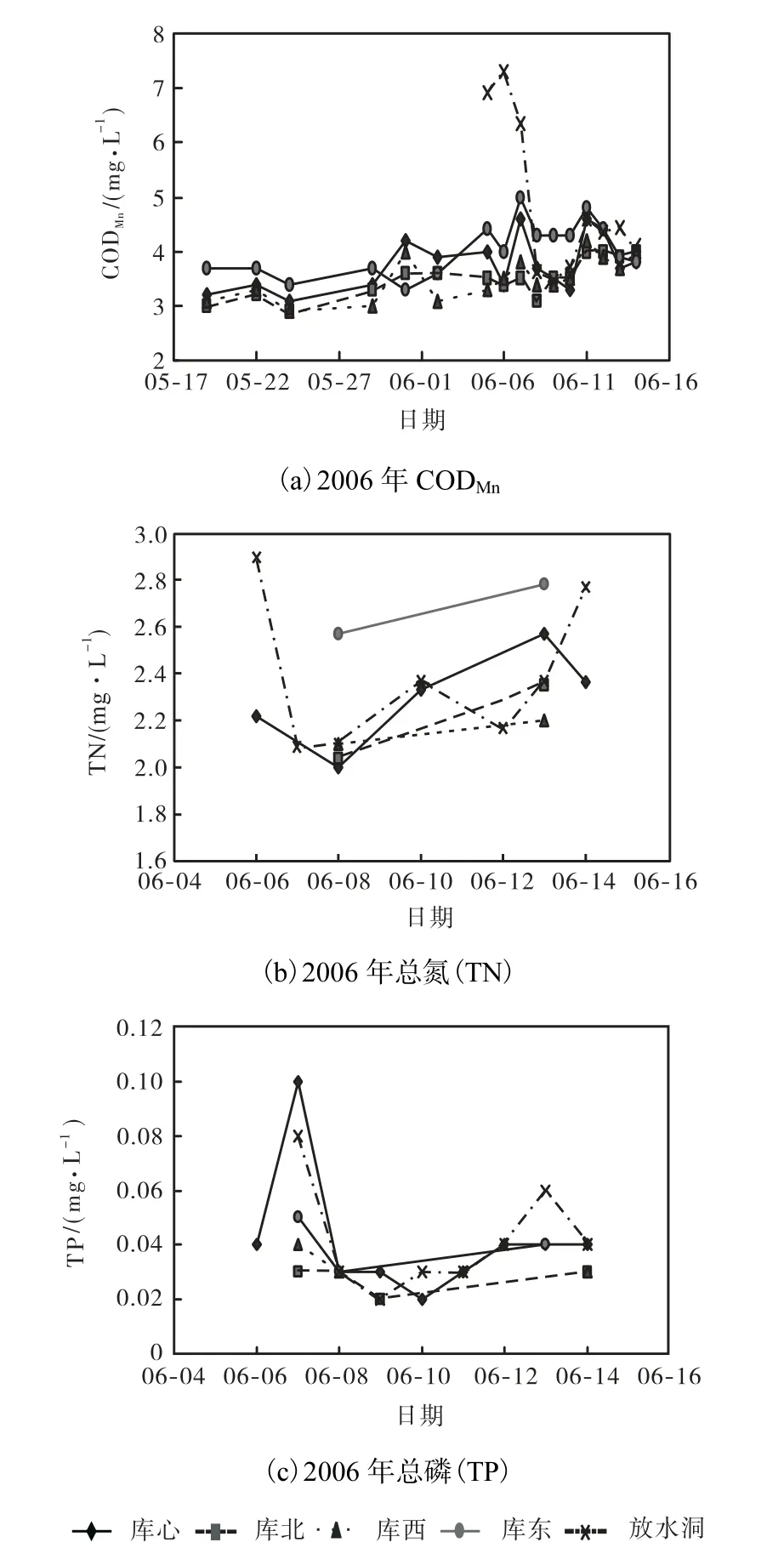
图3 于桥水库各监测断面CODMn、TN和TP变化曲线Fig.3 Variations of CODMn,TN and TP in each monitoring section in Yuqiao Reservoir
综上所述,于桥水库草型富营养化特征显著,5—6月菹草过度生长影响水库供水水质安全.5月中旬至6月上旬是菹草生长期,于桥水库水体 DO 浓度、pH值急剧升高,CODMn、TN和TP浓度略有下降,适量生长对水体有一定净化作用,但过度生长影响水库安全运行;进入6月中旬菹草死亡腐败期,水体DO浓度、pH 值降低,CODMn、TN和TP浓度升高,营养盐含量升高,水质状况恶化程度近1倍,持续时间约为1周.
4 菹草过度生长成因分析
光照、营养物质和水温为菹草生长提供能量,是影响菹草生长率的关键因素.于桥水库呈中度富营养水平,就菹草而言,营养物取之不尽,故导致于桥水库出现菹草疯长现象的主要原因是光照强度(太阳辐射强度)、透明度、水深和水温等条件.
4.1 光照、透明度和水深因素
根据 Lambert-Beer定律,当入射光的波长、介质温度一定时,介质的吸光度 A(%)与介质厚度 L(m)和浊度C(JTU)呈正比的关系,数学表达式为

式中: Ei为入射光照度,lx;Et为透过光照度,lx;μ为吸光系数.
为了便于分析某一水深下的太阳辐照度,对式(1)进行变换,可得

式中: E(H)为水深 H 处的太阳辐照度,W/m2;oE为水面处的太阳辐照度,W/m2;H 为水深,m;μe为由透明度确定的吸光系数,取值范围为0.2~4.0,m-1[13].μe反映的物理意义是在水深一定时,透明度越高,该水深处的太阳辐照度越强,故μe与透明度呈反比,文中定义为透明度的倒数.由式(2)可知,水下太阳辐照度与水深呈负指数关系,且消减的快慢与水体透明度有关,即水下太阳辐照度随着水深的增加而减少,且透明度越低沿水深方向消减越快.
于桥水库 4—6月水位、库心透明度和水下太阳辐照度变化曲线如图 4和图 5所示,数值列于表 1中.由图表可知,4月水位略有升高,升高了 0.66,m,水下太阳辐照度 E ( H )沿水深方向递减,但由于透明度随水深升高而增高,消减减慢,且水面 Eo从171.7,W/m2提高到 208.5,W/m2,故水下 E ( H )有所提高,平均每米提高25.5%,河底处 E ( H )从8.9,W/m2提高到 12.5,W/m2,提高了 40.4%;5月 1日—16日,水位升高了 0.59,m,透明度随水深升高而增高,水面 Eo基本不变,水下 E ( H ) 也变化不大,河底处 E ( H ) 从12.5,W/m2提高到 12.8,W/m2,提高了 2.4%;5月 16日—6月 1日,水位升高了 0.68,m,由于菹草生长繁密,透明度降低,水下 E ( H )沿水深方向消减增快,虽然水面 Eo从 198.7,W/m2提高到 259.4,W/m2,但水下E( H )有所降低,平均每米降低 10.0%,河底处 E ( H)从 12.8,W/m2降低到 3.8,W/m2,降低了 70.3%;6月 1日—15日,水位升高了0.42,m,由于6月中旬菹草死亡后进行了打捞工作,透明度有所回升,水面 Eo基本不变,水下 E ( H )有所提高,平均每米提高 32.6%,河底处 E ( H )从 3.8,W/m2提高到 13.1,W/m2,提高了245%;6月15日—30日,水位降低了0.31,m,透明度随水位降低而降低,水下 E ( H )沿水深方向消减增快,虽然水面 Eo从 251.5,W/m2提高到 297.7,W/m2,但水下 E ( H ) 有所降低,平均每米降低 7.5%,河底处E( H )从13.1,W/m2降低到5.5,W/m2,降低了58.0%.
以上分析结果表明,于桥水库菹草指数生长期内(4月中旬—5月中旬)水下太阳辐照度充足,在11.7,~208.5,W/m2之间,菹草摄取充分的光照和营养快速生长;芽殖体发生期内(5月中旬—6月中旬)虽然水下太阳辐照度出现降低趋势,在 3.8~259.4,W/m2之间,但大部分时间仍然满足菹草生长所需的光照条件,菹草疯长.同时,不难发现,当透明度一定时,水位越高,水下太阳辐照度越低,菹草所能获得的光照能量越少.
4.2 水温因素
春季 3、4月份的于桥水库库心水温监测值如表2所示.由监测数据可知,4月水温处于 10~15,℃,菹草的最适温度范围正是 10~20,℃[14],菹草迅速生长,而其他沉水植物的最适温度为 20~30,℃;5月水库绝大空间内开始繁育芽殖体,占绝对优势种,而此时其他物种才先后萌发,处于弱势种群;6月中旬,随着水温的逐渐升高,菹草植株先后死亡、解体,芽殖体沉入水底越夏休眠.
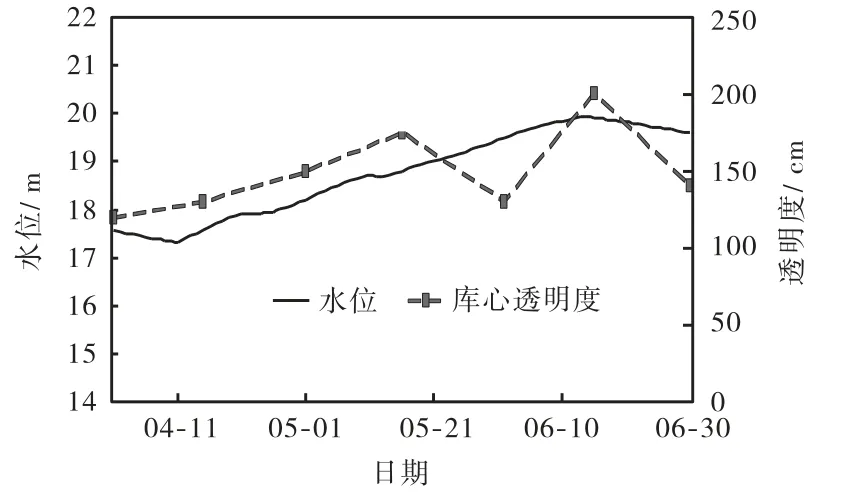
图4 2006年4—6月于桥水库水位和库心透明度变化曲线Fig.4 Variations of water level and water transparency in Fig.4 center monitoring section during April—June 2006

图5 库心水下太阳辐照度随水深和透明度变化曲线Fig.5 Variations of light intensity with depth and transparency in center monitoring section

表1 2006年4—6月于桥水库水位、库心水深、透明度、μe和Eo值Tab.1 Hydraulic variables in center monitoring section during April—June 2006

表2 2006年3月、4月于桥水库库心水温监测值Tab.2 Temperature monitoring values in center monitoring section during March—April 2006
综上所述,由于于桥水库光照强度、透明度、水深和水温条件均有利于菹草生长,使得菹草在快速生长阶段时间和空间上无竞争对手,导致菹草可以充分利用水库空间范围内的各种资源,种群得以充分发育,占尽优势,过度生长使于桥水库草型富营养化越发严重.同时,4月是控制菹草过度生长的关键时期.
5 讨 论
通过上述分析,针对于桥水库草型富营养化特征及菹草疯长原因,可考虑采取提高水位、改变高水位运行时间等措施来抑制菹草过度生长、防治草型富营养化,具体方法如下所述.
通过优化于桥水库调度运行方案,提高水位、改变高水位运行时间,抑制菹草过度生长.近年来,经隧洞、黎河段引滦河水入于桥水库的周期比较稳定,每年分2次调水,一次4月底至6月初,一次9月底至12月初,调水量7×108m3左右;于桥水库向下游市区县输水基本上常年运行,平均每天输水约 163×104m3(18.93,m3/s).统计表明,多年汛前引水量2.0×108m3,引水 45天,日均入库水量 500×104m3、出库水量150×104m3.考虑到3—4月份是菹草生长关键时期(返青期),可将调水日期控制在每年3月中下旬至5月上旬,即将第1次调水期提前20天或1个月,以增加水库蓄量,使水位提高 0.6~1.0,m,如表1中2006年4月1日和4月15日水位提高1,m,假设透明度保持不变,水下太阳辐照度将降低 56.5%,大大降低菹草所需的光照能量,从而抑制菹草过度生长.
菹草生长需要摄取营养盐和光能量,一方面,入库的2×108m3水量,通过水体交换,降低了水体和底质中营养盐含量;另一方面,库水位提高了 0.6~1.0,m,水下光照将减少 1/2左右,消减了菹草吸收的能量.同时,引水期间入库水流流速较大,对菹草生长有一定破坏作用.因此,在菹草生长到成熟期以前即返青期,提高库水位,改变高水位运行时间是抑制菹草过度生长、防治草型富营养化的有效措施之一,但仍需对调水量、水库水位及抑制效果做进一步系统地研究.
6 结 论
(1) 6月中旬,过量菹草衰亡释放的高营养盐和有机质对水质影响较大,恶化程度近 1倍,持续时间约为 1周.研究表明,导致于桥水库出现菹草过度生长现象的主要原因是光照度(太阳辐照度)、透明度、水深和水温等条件.
(2) 4月是控制菹草过度生长的关键时期,对Lambert-Beer定律公式进行变换,得到水下太阳辐照度与水深呈负指数关系,提出采用提高水位、改变高水位运行时间抑制菹草过度生长的新方法.
[1]王国祥,濮培民,张圣照,等. 冬季水生高等植物对富营养化湖水的净化作用[J]. 中国环境科学,1999,19(2):106-109.
Wang Guoxiang,Pu Peimin,Zhang Shengzhao,et al. The purification ability of aquatic macrophytes for eutrophic lake water in winter[J].China Environmental Science,1999,19(2):106-109(in Chinese).
[2]王文林,王国祥,李 强,等. 菹草-伊乐藻群落对富营养化水体水质的净化效果[J]. 南京师范大学学报:自然科学版,2006,29(4):111-116.
Wang Wenlin,Wang Guoxiang,Li Qiang,et al. Purification effect ofPotamogeton crispus-Elodea nuttalliicommunity on eutrophic water body quality[J].Journal of Nanjing Normal University:Natural Science,2006,29(4):111-116(in Chinese).
[3]Hu M H,Ao Y S,Yang X E,et al. Treating eutrophic water for nutrient reduction using an aquatic macrophyte(Ipomoea aquatica Forsskal)in a deep flow technique system[J].Agricultural Water Management,2008,95:607-615.
[4]Dierberg F E,DeBusk T A,Jackson S D,et al. Submerged aquatic vegetation-based treatment wetlands for removing phosphorus from agricultural runoff:Response to hydraulic and nutrient loading[J].Water Research,2002,36(6):1409-1422.
[5]尚士友,申庆泰,杜健民,等. 内蒙古乌梁素海沉水植物的收割工程技术[J]. 湖泊科学,2004,16(2):169-177.
Shang Shiyou,Shen Qingtai,Du Jianmin,et al. The harvesting project technology of submerged plant in Lake Wuliangsuhai of Inner Mongolia[J].Journal of LakeSciences,2004,16(2):169-177(in Chinese).
[6]尚士友,杜健民,李旭英,等. 草型富营养化湖泊生态恢复工程技术的研究:内蒙古乌梁素海生态恢复工程试验研究[J]. 生态学杂志,2003,22(6):57-62.
Shang Shiyou,Du Jianmin,Li Xuying,et al. Ecological restoration engineering technology of eutrophic lake:A case study of Lake Wuliangsuhai in Inner Mongolia[J].Chinese Journal of Ecology,2003,22(6):57-62(in Chinese).
[7]顾久君,金朝晖,刘振英. 乌梁素海沉水植物腐烂分解试验研究[J]. 干旱区资源与环境,2008,22(4):181-184.
Gu Jiujun,Jin Zhaohui,Liu Zhenying. Experimental studies on decomposition process of submerged macrophytes from Wuliangsuhai Lake[J].Journal of Arid Land Resources and Environment,2008,22(4):181-184(in Chinese).
[8]Miranda L E,Hodges K B. Role of aquatic vegetation coverage on hypoxia and sunfish abundance in bays of a eutrophic reservoir[J].Hydrobiologia,2000,427:51-57.
[9]Frodge D J,Thomas G L,Pauley G B. Effects of canopy formation by floating and submergent aquatic macrophytes on the water quality of two shallow Pacific Northwest lakes[J].Aquatic Botany,1990,38:231-248.
[10]Roman C T,Barrett N E,Portnoy J W. Aquatic vegetation and trophic condition of Cape Cod(Massachusetts,USA)kettle ponds[J].Hydrobiologia,2001,443:31-42.
[11]Coops H,Doef R W. Submerged vegetation development in two shallow,eutrophic lakes[J].Hydrobiologia,1996,340:115-120.
[12]张 晨,刘万宏,王立义,等. 模糊数学在水库水质综合评价中的应用[J]. 安全与环境学报,2009,9(1):90-92.
Zhang Chen,Liu Wanhong,Wang Liyi,et al. Application of fuzzy mathematics in the evaluation of the water quality of reservoirs[J].Journal of Safety and Environment,2009,9(1):90-92(in Chinese).
[13]Ji ZhenGang.Hydrodynamics and Water Quality:Modeling Rivers,Lakes,and Estuaries[M]. New Jersey:John Wiley & Sons,Inc,2007:74.
[14]金送笛,李永函,王永利. 几种生态因子对菹草光合作用的影响[J]. 水生生物学报,1991,15(4):295-302.
Jin Songdi,Li Yonghan,Wang Yongli. Effects of ecological factors on photosynthesis ofPotamogeton crispus[J].Acta Hydrobiologica Sinica,1991,15(4):295-302(in Chinese).
